Гидробиоценозы соленых рек бассейна гипергалинного озера Эльтон (ООПТ природный парк «Эльтонский»)
")
Автор: Зинченко Т.Д., Шитиков В.К.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Оригинальные статьи
Статья в выпуске: 4 т.34, 2025 года.
Бесплатный доступ
Представлены результаты анализа гидробиоценозов соленых рек аридного региона Приэльтонья (район Прикаспийской низменности, бассейн гипергалинного озера Эльтон) на базе изучения планктонных и донных сообществ в августе 2013 и 2018 гг. Использовались результаты обработки гидробиологических проб, взятых на 13 станциях пяти высокоминерализованных рек. Была сформирована матрица численностей 136 видов и таксонов, представителей зоопланктона, макрозообентоса и мейобентоса. Для обработки данных с помощью многомерного статистического анализа значения матриц численностей трансформировали в единую шкалу баллов от 0 до 6 с применением алгоритма нахождения оптимальных границ. Станции отбора проб сгруппированы в зависимости от уровня минерализации воды: 1 – >25 г/л, 2 – 10–25 г/л и 3 – <10 г/л. Использование статистических моделей размещения видов в пространстве абиотических факторов позволило установить, что неоднородность разнообразия планктонных и донных сообществ обусловлена биотопическими отличиями и уровнем минерализации водных масс. Показано, что для высокоминерализованных гидросистем Приэльтонья четкое разделение жизненных форм планктонных и донных сообществ не очевидно.
Соленые реки, Приэльтонье, зоопланктон, макрозообентос, мейобентос, структура сообществ, статистический анализ
Короткий адрес: https://sciup.org/148333004
IDR: 148333004 | УДК: 574.52(282.247.431.2) | DOI: 10.24412/2073-1035-2025-34-4-4-11
Hydrobiocenoses Of The Saline Rivers Of The Basin Of Hypergaline Lake Elton (Protected Area Nature Park
The results of the analysis of hydrobiocenoses of saline rivers in the arid region of the Elton region (the area of the Caspian lowland, the basin of the hypergaline Lake Elton) based on the study of planktonic and bottom communities in August 2013 and 2018 are presented. The results of processing hydrobiological samples taken at 13 stations of five highly mineralized rivers were used. A matrix of numbers of 136 species and taxa representing zooplankton, macrozoobenthos, and meiobenthos was formed. To process the data using multidimensional statistical analysis, the values of the numerical matrices were transformed into a single score scale from 0 to 6 using an algorithm for finding optimal boundaries. Sampling stations are grouped according to the level of mineralization of water: 1 – >25 g/l, 2 – 1025 g/l and 3 – <10 g/l. The use of statistical models of the distribution of species in the space of abiotic factors allowed us to establish that the heterogeneity of the diversity of planktonic and bottom communities is due to biotopic differences and the level of mineralization of water masses. It is shown that for the highly mineralized hydraulic systems of the Elton region, a clear separation of the life forms of planktonic and bottom societies is not obvious.
Текст научной статьи Гидробиоценозы соленых рек бассейна гипергалинного озера Эльтон (ООПТ природный парк «Эльтонский»)
обычно входят виды с различным диапазоном толерантности к факторам среды.
Ниже приводится анализ гидрохимических и гидробиологических данных, полученных при отборе проб в 2013, 2018 гг. на разных биотопах пяти соленых рек. Изучено взаимодействие между видами макрозообентоса, мейобентоса и зоопланктона и абиотическими факторами. Применение комплекса методов многомерного статистического анализа позволило показать корреляционные связи распределения видов в структуре изучаемого метасообщества: согласованная встречаемость видов ( coherence ), образование ассоциаций видов ( clumping ), смена видового состава вдоль экологических градиентов ( turnover ).
Напомним, что изученные нами мезо- поли-и гипергалинные гидросистемы Приэльтонья функционируют в условиях стохастических, а часто и экстремальных природно-климатических воздействий, характерных для аридных зон, что вызывает резкие флуктуации численности отдельных популяций гидробионтов (Zinchenko et al., 2017). Все это определяет актуальность и неподдельный интерес к изучению взаимозави-симостей планктонных и донных сообществ высокоэвтрофных рек.
Нами было показано, что для изучаемых экосистем четкое разделение жизненных форм планктонных и донных сообществ не очевидно, т.е. отмечен переход бентосных животных к планктонному образу жизни, что приводит к высокой доле взаимопроникающих экологических группировок. Для анализа выраженности таких ассоциативных внутрисистемных связей использовался метод оценки канонических корреляций, выявивший тесную статистическую связь между группами макрозообентоса, мейобентоса и зоопланктона (Зинченко и др., 2019а). Сравнение результатов многолетних исследований показало как наличие высокой структурной согласованности и устойчивых закономерностей пространственного распределения, так и смену комплекса ведущих видов сообществ (Зинченко и др., 2019б). Многомерный ординационный анализ позволил установить характер согласованных реакций видов на изменение водной среды, а также выявить комплексы гидрохимических факторов, оказывающих наиболее значимое влияние.
Первым этапом изучения потенциальных механизмов формирования мета-сообщества является проверка статистической гипотезы о соответствии модели, основанной на наблюдениях, и стандартных паттернов, типичных при определенных условиях (Connor, Simberloff, 1979). В качестве модели-эталона обычно выбирается та или иная нулевая модель, т.е. распределение видов по участкам случайно и хаотично, когда кластеры не образуются, а влияние экологических градиентов отсутствует и связей между видами не обнаруживается (Sanderson, 2004; Hausdorf, Hennig, 2007).
В статье нами представлен дополнительный анализ результатов обработки проб макрозообентоса, мейобентоса и зоопланктона на участках пяти соленых рек и предложены подходы к решению следующих актуальных проблем теоретического и практического характера:
-
1. Анализ представленности основных элементов видовой структуры изучаемого метасообщества, в том числе, проверка гипотез о согласованной встречаемости видов ( coherence ), образовании компактных ассоциаций видов ( clumping ) и закономерной смене видового состава вдоль экологических градиентов ( turnover ).
-
2. Селекция набора видов, обилие которых статистически значимо связано с экологическим градиентом.
-
3. Построение корреляционной сети, определяющей наиболее важные связи внутри экосистемы.
Материал и методы
Исходный материал получен по результатам обработки образцов макрозообентоса, мейобентоса и зоопланктона на 13 различных участках пяти соленых рек бассейна оз. Эльтон (рр.Хара, Солянка, Большая Саморода, Черновка, Ланцуг), имеющих значительный градиент минерализации (от 6 до 50,1 г/л). Подробное описание и схема района исследований, методы отбора и обработки собранного материала представлены в публикациях (Zinchenko et al., 2017; Зинченко и др., 2018; 2019а; 2019б). На станциях наблюдений и отбора проб параллельно проводили мониторинг факторов среды, включающий измерения гидрологических и гидрохимических параметров. По совокупности последних каждый участок реки был отнесен к одной из трех градаций солености.
Для проведения статистического анализа использовали результаты двух гидробиологических съемок в августе 2013 и 2018 гг. на одних и тех же фиксированных створах. Полученные данные позволили сформировать таблицу численностей 136 таксономических групп, в том числе: 36 видов и родов макрозообентоса, 50 видов мейобентоса (экз./м2) и 50 вид зоопланктона (экз./м3). Для корректной совместной обработки данных значения численностей трансформировали в единую шкалу баллов от 0 до 6 и сформировали обобщенную матрицу таксономического обилия размерностью 26 x 136.
Анализ трех основных элементов структуры метасообщества (согласованности, оборота и сгущений видов – (Leibold, Mikkelson, 2002) проводили с целью установки типа паттерна для отображения распределения видов вдоль главной оси вариации факторов среды. Для этого на основе эмпирических данных формировали упорядоченную матрицу инцидентности, которую сопоставляли с аналогичными случайными матрицами с использованием предложенных статистических критериев (Presley et al., 2010). Частичная рандомизация нуль-модели предполагала неизменное видовое богатство участков и осуществлялась методом r1, то есть, установка в единицу произвольного элемента матрицы проводилась с вероятностью, пропорциональной общей частоте вида. Оценку значимости элементов структуры осуществляли в результате многократного повторения имитационного процесса Монте-Карло.
Тест на кластеризацию, для проверки гипотезы о наличии положительных ассоциаций видов, проводили на основе параметрического бутстре-па матрицы дистанций между участками с использованием количественной меры Кульчицкого. Применяли статистический критерий (Hennig,
Hausdorf, 2004), учитывающий соотношение суммарных расстояний внутри и между кластерами. Простейшая неограниченная ( unconstrained ) нулевая модель предполагала доступность биотопов для всех видов. В моделях с ограничениями задавалась определенная корреляционная структура в виде матрицы расстояний между отдельными участками по шкале экологического, пространственного или временного градиента.
Ранее (Зинченко и др., 2018; 2019а; 2019б) для ранжирования видов по степени их индикаторной валентности по отношению к градиенту солености использовался индекс IndVal Дюфрена-Лежандра. В представленном разделе применялся алгоритм «Борута» (Boruta) (Kursa, Rudnicki, 2014), который оценивает меру информативности каждого вида. Метод имеет хорошее статистическое обоснование, использует внешнее дополнение, полученное в ходе рандомизации, и выполняет проверку Н 0 на основе высокоэффективного ансамбля моделей случайного леса ( Random Forest ).
Тесноту статистической связи между отобранными видами оценивали методом «графического лассо» ( graphical lasso – (Friedman et al., 2007), с помощью которого находили оценку максимального правдоподобия матрицы коэффициентов частной корреляции Σ на основе алгоритма регуляризации (т.е. с учетом штрафа за сумму абсолютных значений Σ -1) . Полученный оптимальный граф отображен и интерпретирован нами в формате корреляционной сети, построенной по алгоритму Фрухтермана-Рейнгольда.
Статистический анализ выполняли с использованием среды программирования R версии 3.06 и ее дополнительных пакетов metacom, prabclus, Boruta, huge и qgraph.
Результаты и обсуждение
Проверка значимости статистических гипотез относительно закономерностей таксономической структуры метасообществ представлена в табл. 1. Отрицательное значение Z -критерия и значимость статистики для теста на согласованность видов свидетельствует о преобладании паттерна типа «шахматная доска»: композиции видов имеют больше пропусков непрерывности ( embedded absences ), чем это имеет место для случайной структуры. Незначимый Z -критерий для оборота видов подтверждает, что эти пропуски имеют равновероятный характер («плато мета-сообщества, пронизанное отверстиями»). В то же время, тест χ 2 на наличие сгущений отклоняет нулевую гипотезу об отсутствии кластеризации. Все эти особенности имеют место в случае, когда доминирующий градиент среды отсутствует, а распределение видов формируется под влиянием нескольких, в некотором смысле разнонаправленных внешних факторов.
Для проверки предположения о существовании пространственной автокорреляции видового состава участков рек с точками их окрестности было построено четыре модели (табл. 1). Две модели основаны на гипотезе, что вероятность образования повторяющихся ассоциаций видов больше на близких между собой участках (в пределах одной реки или принадлежащих к одной и той же градации солености). Для них выявлен статистически значимый уровень кластеризации таксонов ( р = 0,05 ÷ 0,06). Для моделей, считающих, что пространственной автокорреляции нет или она основана лишь на близости географических координат точек, гипотеза о случайном характере выявляемых кластеров не отклоняется.
Таблица 1. Статистическая значимость различных предположений относительно таксономической структуры метасообщества гидробионтов в соленых реках
Table 1. Statistical significance of various assumptions regarding the taxonomic structure of the hydrobiont meta-community in saline rivers
|
Проверяемые гипотезы |
Статистика |
р -значение |
|
1. Элементы метаструктуры (Leibold, Mikkelson, 2002): – согласованность видов ( coherence ) – оборот видов ( turnover ) – наличие сгущений ( boundary clumping ) |
–9,89 0,88 1,71 |
≈ 0 0,37 0,000001 |
|
2. Выраженность ассоциаций видов при различных ограничениях (Hennig, Hausdorf, 2004): – неограниченная модель – связь с пространственным расположением – локализация по рекам – связь с градиентом солености |
0,143 0,286 0,0495 0,0594 |
Для уточнения видов, образующих взаимосвязанные ассоциации, строили граф сети, представленный на рис. 1. Использованные коэффициенты частной корреляции обычно в 3-4 раза по абсолютной величине меньше, чем традиционные коэффициенты корреляции Пирсона, поскольку оценивают только «чистые» эффекты парного взаимодействия между двумя конкретными узлами сети после исключения совокупно- го влияния всех остальных узлов. Тем самым резко снижается вероятность эффекта «ложной корреляции». Корреляционные «плеяды» на рис. 1 состоят из комбинаций видов, совместная встречаемость которых обусловлена как биотическими взаимодействиями, так и взаимно согласованной реакцией на изменение факторов водной среды.
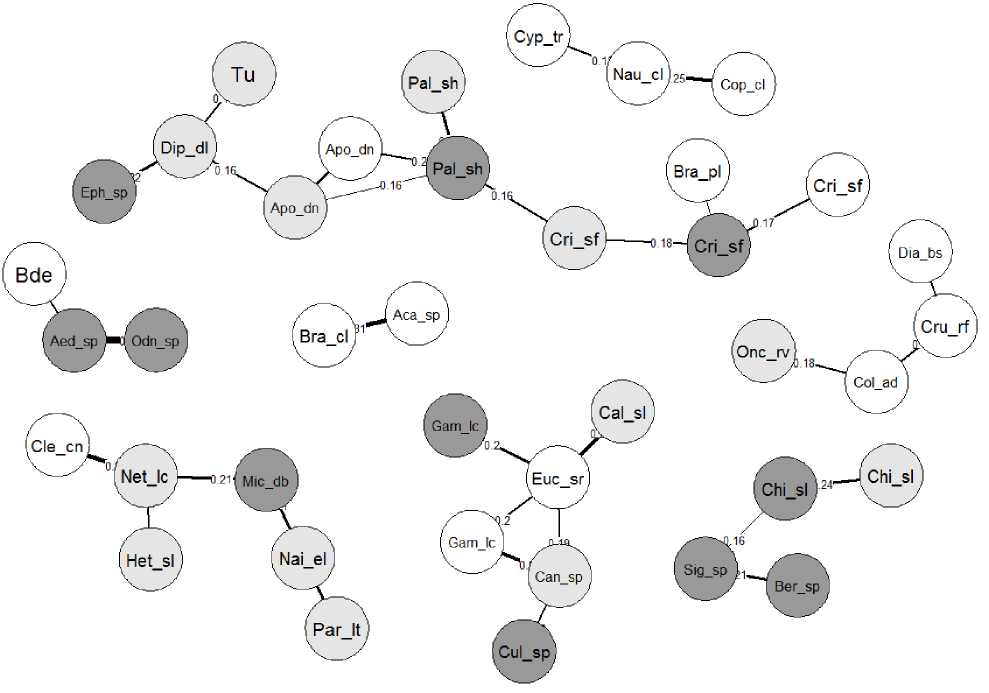
Рис. 1. Граф фрагмента сети частной корреляции видов гидробионтов.
(Здесь и далее на рис. 2 коды видов представлены в табл. 2; темно-серым цветом отмечены таксоны, выделенные в пробах макрозообентоса, серым – мейобентоса и белым – зоопланктона (Зинченко и др., 2020).
Fig. 1. Graph of a fragment of a partial correlation network of aquatic species. (Here and further in Fig. 2, the species codes are shown in the Table 2; taxa isolated from macrozoobenthos samples are marked in dark gray, meiobenthos in gray, and zooplankton in white (Zinchenko et al., 2020).
Для оценки тесноты связи видов гидробионтов с градиентом минерализации выполнили 99 итераций построения моделей «случайного леса», состоящих из 500 иерархических деревьев. С помощью алгоритма «Борута» оценивали важность (importance) каждого вида по Z-критерию, вычисленного с учетом снижения ошибки прогнозирования при замене эмпирического вектора обилия на случайный вектор. Было установлено (табл. 2), что из 136 исходных видов только 16 таксонов имеют статистически значимую связь с фактором минерализации воды, тогда как нахождение прочих видов в любой точке изучаемой системы соленых рек является случайным и равновероятным.
Чтобы оценить, какие показатели биотопов оказывают влияние на обитание отдельных видов, выполнен анализ главных компонент исходных матриц. На рис. 2 показан двудольный корреляционный граф, показывающий связи между 25 первыми таксонами из табл. 2 и тремя латентными факторами, определяющими главные оси многомерной вариации данных. Фактор 1 включает комплекс показателей, определяющих
Таблица 2. Таксоны зоопланктона (ZP), мейобентоса (MB) и макрозообентоса (ZB), обладающие наибольшей значимостью при построении моделей зависимости видового состава от градиента солености
Table 2. Taxa of zooplankton (ZP), meiobenthos (MB) and macrozoobenthos (ZB), which are of the greatest importance in constructing models of the dependence of species composition on the salinity gradient
|
Значимость |
Группа |
Код вида |
Наименование |
Встречаемость |
Z -критерий |
р -зна-чение |
|
Высокая |
ZB |
Pal_sh |
Palpomyia schmidti |
11 |
10,75 |
1,00 |
|
MB |
Apo_dn |
Apocyclops dengizicus |
8 |
6,92 |
1,00 |
|
|
ZP |
Apo_dn |
Apocyclops dengizicus |
9 |
8,60 |
1,00 |
|
|
ZP |
Cri_sf |
Cricotopus salinophilus |
19 |
7,15 |
0,99 |
|
|
ZP |
Aca_sp |
Acanthocyclops americanus |
4 |
3,73 |
0,75 |
|
|
ZB |
Cri_sf |
Cricotopus salinophilus |
16 |
3,65 |
0,73 |
|
|
ZP |
Bra_pl |
Brachionus plicatilis |
22 |
3,53 |
0,71 |
|
|
MB |
Het_sl |
Heterocypris salina |
5 |
3,31 |
0,69 |
|
|
ZP |
Cle_cn |
Cletocamptus confluens |
4 |
3,29 |
0,63 |
|
|
ZB |
Gam_lc |
Gammarus lacustris |
6 |
3,11 |
0,60 |
|
|
MB |
Mic_db |
Microchironomus deribae |
3 |
3,06 |
0,57 |
|
|
MB |
Chi_sl |
Chironomus salinarius |
11 |
2,77 |
0,51 |
|
|
Средняя |
MB |
Pal_sh |
Palpomiya schmidti |
7 |
2,29 |
0,38 |
|
ZP |
Meg_vr |
Megacyclops viridis |
7 |
2,18 |
0,37 |
|
|
MB |
Cyp_tr |
Cyprideis torosa |
17 |
2,32 |
0,36 |
|
|
MB |
Cri_sf |
Cricotopus salinophilus |
15 |
2,07 |
0,12 |
|
|
ZP |
Cru_rf |
Cricotopus rufiventris |
3 |
1,43 |
0,05 |
|
|
ZB |
Gly_sl |
Glyptotendipes salinus |
4 |
1,56 |
0,03 |
|
|
ZP |
Dia_bs |
Diacyclops bisetosus |
3 |
1,09 |
0,03 |
|
|
ZB |
Chi_ap |
Chironomus aprilinus |
4 |
0,35 |
0,02 |
|
|
MB |
Nai_el |
Nais elinguis |
4 |
1,54 |
0,02 |
|
|
Слабая |
ZB |
Tan_kh |
Tanytarsus kharaensis |
2 |
1,32 |
0,01 |
|
ZB |
Mic_db |
Microchironomus deribae |
8 |
0,96 |
0,01 |
|
|
MB |
Onc_rv |
Oncholaimus rivalis |
4 |
0,70 |
0,01 |
|
|
MB |
Mo_pv |
Monhystrella parvella |
23 |
1,62 |
0,01 |
|
|
ZB |
Chi_sl |
Chironomus salinarius |
10 |
0,09 |
∼ 0 |
|
|
ZB |
Aed_sp |
Aedes sp. |
3 |
0,25 |
∼ 0 |
|
|
MB |
Net_lc |
Nitokra lacustris |
3 |
1,07 |
∼ 0 |
|
|
MB |
Par_lt |
Paranais litoralis |
3 |
0,95 |
∼ 0 |
|
|
MB |
Can_sp |
Candona spp. |
3 |
0,88 |
∼ 0 |
|
|
MB |
Dip_dl |
Diplolaimelloides delyi |
5 |
0,72 |
∼ 0 |
|
|
ZB |
Ber_sp |
Berosus sp. |
3 |
0,72 |
∼ 0 |
|
|
MB |
Cal_sl |
Calodorylaimus salinus |
3 |
0,42 |
∼ 0 |
|
|
Для рис. 1 |
ZP |
Euc_sr |
Eucyclops serrulatus |
3 |
0,02 |
∼ 0 |
|
MB |
Tu |
Turbellaria |
4 |
0,00 |
∼ 0 |
|
|
ZB |
Cul_sp |
Culicoides sp. |
3 |
–0,08 |
∼ 0 |
|
|
ZP |
Cyp_tr |
Cyprideis torosa |
6 |
–0,40 |
∼ 0 |
|
|
ZP |
Gam_lc |
Gammarus lacustris |
3 |
–0,44 |
∼ 0 |
|
|
ZB |
Odn_sp |
Odontomyia sp. |
3 |
–0,45 |
∼ 0 |
|
|
ZB |
Sig_sp |
Sigara sp. |
4 |
–0,50 |
∼ 0 |
|
|
ZB |
Eph_sp |
Ephydra sp. |
4 |
–0,56 |
∼ 0 |
|
|
ZP |
Nau_cl |
Nauplii Cyclopoida |
12 |
–0,63 |
∼ 0 |
|
|
ZP |
Col_ad |
Colurella adriatica |
3 |
–0,72 |
∼ 0 |
|
|
ZP |
Bde |
Bdelloida |
4 |
–0,80 |
∼ 0 |
|
|
ZP |
Cop_cl |
Copepodit Cyclopoida |
8 |
–0,81 |
∼ 0 |
|
|
ZP |
Bra_cl |
Brachionus calyciflorus |
3 |
–1,72 |
∼ 0 |
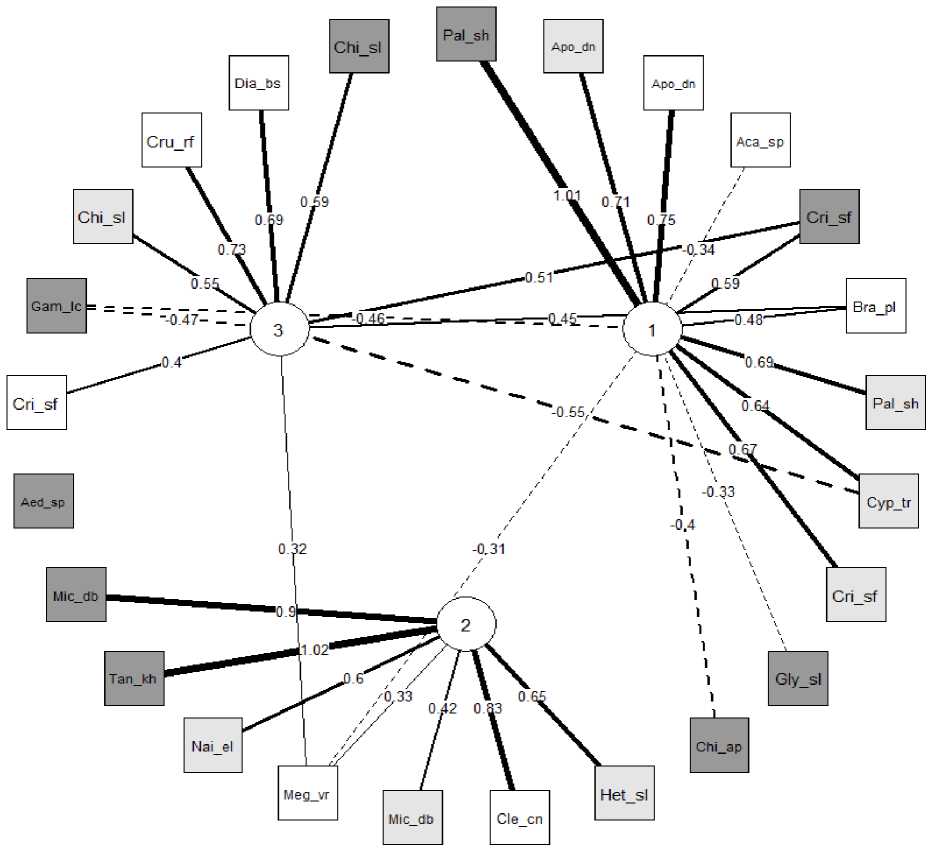
Рис. 2. Двудольный корреляционный граф между видами гидробионтов и тремя главными компонентами (Зинченко и др., 2020)
Fig. 2. Bipartite correlation graph between species of aquatic organisms and three main components (Zinchenko et al., 2020)
соленость воды: содержание хлорид-иона, кальция и всех трех форм азота (положительная корреляция +), а также сульфат-иона и глубину водотока (отрицательная корреляция –), фактор 2 тесно связан с содержанием кислорода и рН (+), концентрацией марганца и железа (–), а фактор 3 связан с содержанием хлорофилла «a», магния и с температурой воды (+), а также фосфатами и бикарбонатами (–).
Таким образом, популяции видов планктонных и донных сообществ соленых рек бассейна оз. Эльтон характеризуются достаточно устойчивыми закономерностями пространственного распределения численности в зависимости от факторов воздействия. Сравнение значимости моделей зависимости видового состава популяций 136 видов и таксонов от градиента солености по результатам исследований в 2013 и 2018 гг. позволило выявить довольно тесную сопряжен- ность видов в планктонных и донных сообществах от уровня минерализации. В то же время, выявленные флуктуации таксономического состава гидробионтов, динамичность гидрохимических характеристик и гидрологических факторов в условиях градиента солености позволяют рассматривать экосистему как некий пул популяций в системе средовой неопределенности. То есть, большинство видов распределены отдельными пятнами, что можно рассматривать, например, как «популяцию популяций» (Levins, 1970), указывая тем самым на неравновесность биотического разнообразия в условиях динамичности природных (зарастаемость биотопов макрофитами; сгонно-нагонные явления соленой воды в реки из гипергалинного озера в системе озеро–река) и климатических изменений. Обсуждая структуру распределения, взаимопроникновения, миграции популяций (Федоров, 2004)
планктонных и донных сообществ на примере высокоэвтрофных соленых рек, можно констатировать их тесную взаимосвязь. Эта зависимость обусловлена как биотическими взаимодействиями популяций видов фито- и бактериофагов при возможном «ключевом параметре» – ресурсной обеспеченности в эвтрофных водотоках (Зинченко и др., 2010, 2020; Zinchenko et al., 2014), так и реакцией видов на динамичность условий водной среды. Очевидно, что планктонные и донные сообщества следует рассматривать в их целостности с множеством взаимосвязанных элементов (по отношению друг к другу и окружающим условиям). Следует помнить, что особые формы взаимодействия со средой и между видами осуществляются, главным образом, посредством метаболитов, которые, как отмечает Г.Г. Винберг (1967), «служат важным интегрирующим водные сообщества фактором, подверженным действию естественного отбора».
Выполненный анализ элементов структуры метасообщества может быть использован для изучения планктонных и донных биоценозов, позволяющий сформулировать ряд гипотез для организации более детальных исследований и выявления процессов, обусловливающих установленное распределение видов в гиперэвтрофных условиях при высокой минерализации. Факторами, оказывающими значимое влияние на таксономическую структуру метасообществ, являются: содержание хлоридов, кальция, ионов аммония, сульфатов, фосфатов, бикарбонатов, концентрации марганца, железа, магния, хлорофилла «a» в условиях различных гидрофизических факторов – содержания растворенного кислорода, глубины, рН, температуры воды. По мнению В.Д. Федорова (Федоров, 2004: стр. 137), «Любой природный биотоп структурирован. Его структура определяется наличием всевозможного рода градиентов, отражающих изменение однородности его свойств в четырехмерном (четвертое измерение – время) пространстве».
Выделенные таксоценозы, адаптированные к обитанию в высокоминерализованных водах, обладают различной связанностью с ключевыми абиотическим факторами. Результаты анализа показали, что планктонные и донные сообщества соленых рек могут рассматриваться как консорциум метасообществ или структурных единиц речных экосистем, что в условиях Эльтонского парка подразумевает сохранения природной динамики естественных местообитаний.
Полученные данные структурной организации таксоценозов необходимы для понимания функционирования экосистем при проведении экологического мониторинга состояния уникальной гидроэкосистемы Приэльтонья.
Работа выполнена по теме НИР «Оценка современного биоразнообразия и прогноз его изменения для экосистем Волжского бассейна в условиях их природной и антропогенной трансформации» и при финансовой поддержке Российского фонда фундаментальных исследований, грант № 17-04-00135.
