Гидрофобные свойства и пленкообразующая способность штаммов рода Pseudomonas, изолированных из разных экологических ниш

Автор: Демаков В.А., Кузнецова М.В., Карпунина Т.И., Николаева Н.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2010 года.
Бесплатный доступ
Обнаружено, что большинство почвенных и клинических культур были пленкообразующими. Как гидрофильные по BATH-тесту штаммы (из водной фазы в фазу гексадекана экстрагировалось не более 5-8% клеток), так и более гидрофобные псевдомонады (экстрагировалось до 24,2%) проявляли высокую пленкообразующую способность, т.е. не было обнаружено четкого взаимного соответствия между гидрофильно-гидрофобными свойствами клеточной стенки бактерий и степенью пленкообразования. У клинических штаммов гетерогенность популяции по обоим признакам более выражена.
Почвенные бактерии, клинические штаммы, гидрофобность, специфическая и не- специфическая адгезия, пленкообразующая способность
Короткий адрес: https://sciup.org/147204880
IDR: 147204880 | УДК: 579.841.11:
Текст научной статьи Гидрофобные свойства и пленкообразующая способность штаммов рода Pseudomonas, изолированных из разных экологических ниш
В природных экотопах основная часть микроорганизмов существует в виде ассоциаций, определяемых общим термином «биопленки»: пространственно и метаболически структурированных сообществ, расположенных на границе раздела фаз и заключенных во внеклеточный полимерный матрикс (Николаев, Плакунов, 2007). Развитие прикрепленных сообществ – одна из основных стратегий выживания бактерий не только в окружающей среде, но и в организмах инфицируемых хозяев (Ильина и др., 2006). Установлено, что многие хронические инфекции вызываются бактериями, растущими в виде биопленок (Радионович и др., 2006; Costerton et al., 1999).
Формирование микробных сообществ проходит несколько стадий развития. На первом этапе перехода от планктонной формы к прикрепленному состоянию происходит сначала обратимая, а затем необратимая адгезия к поверхности (Сидоренко, 2004). На этапе обратимого прикрепления действуют неспецифические физико-химические силы взаимодействия между молекулами и структурами на поверхностях микроорганизма и твердого субстрата (Николаев, Плакунов, 2009). Показатель гидрофобности является одной из ключевых характеристик клеточной поверхности, определяющий адсорбционную иммобилизацию микроорганизмов. Вместе с тем, диссоциативные варианты бактерий, отражающие различия/изменения в клеточной оболочке, могут значительно отличаться по гидрофобности и адгезивным свойствам (Милько, Егоров,
-
1996) . Ответить на вопрос, является ли гидрофобность свойством, определяющим исход неспецифического взаимодействия, представляется возможным по конечному результату – эффективности формирования биопленок разными штаммами.
Цель данной работы – изучение гидрофобности и пленкообразующей способности различных представителей рода Pseudomonas .
Материалы и методы исследования
В работе использовали 16 штаммов почвенных бактерий рода Pseudomonas (I группа) и 11 изоля-тов Pseudomonas aeruginosa (II группа) , выделенных из различного клинического материала.
Гидрофобность клеток оценивали по относительному распределению между водной фазой и фазой органического растворителя гексадекана (Коваленко и др., 1998; Rosenberg et al., 1980). Для этого к суспензии клеток (3 мл), термостатированной в течение 15 мин при 35°С, добавляли гексадекан (0.2 мл, 0.5 мл), интенсивно встряхивали в течение 1 мин, затем снова термостатировали в аналогичном режиме и регистрировали изменение оптической плотности в водной фазе при λ = 590 нм. Среднее значение относительного изменения (в %) оптической плотности клеточной суспензии в водной фазе по сравнению с ее первоначальной величиной служило мерой гидрофобности бактериальных клеток. Расчет осуществляли по форму-
ле: % гидрофобности = (А с – A г ) / А с х 100, где А с – ОП 590 суспензии до обработки гексадеканом, A г – ОП 590 суспензии после обработки гексадеканом.
Образование биопленок изучали на поверхности 96-луночной полистироловой планшеты согласно И.А. Шагиняну (Шагинян и др., 2007). Суточные бульонные культуры в разведении 1/100 инокулировали в лунки по 150 мкл в 4-х повторах. После 24-часовой инкубации при 37°C содержимое лунок (планктонную культуру) аккуратно удаляли, пленки окрашивали 0.1% раствором генци-анвиолета (170 мкл в лунку) в течение 45 мин. Затем после 3-кратного промывания в лунки для экстрагирования краски добавляли по 200 мкл 96% этанола. Уровень экстракции (абсорбции) генци-анвиолета этанолом измеряли на спектрофотометре Ultrоspec 3300 при длине волны 580 нм в единицах оптической плотности (Ед, ОП 580 ). Степень пленкообразования соответствовала интенсивности окрашивания содержимого лунок красителем. Пленкообразующими считали культуры, если значение ОП превышало контроль в 3 и более раз (ОП 580 ≥1.0), склонными к адгезии – в 2–3 раза (0.6≥ОП 580 <1.0), остальные оценивали как непленкообразующие. Контролем служили лунки с LB-бульоном (Луриа-Бертани) (ОП 580 ≤0.3).
Результаты и их обсуждение
Показатель гидрофобности клеточной поверхности псевдомонад из различных экологических ниш не превышал 25%, M±σ составила 5.9±3.2% – для «почвенной» группы и 9.5±7.2% – для «клинической». Этот показатель варьировал у разных штаммов в обеих группах, причем более значительно во II группе.
При сравнении показателя пленкообразования в обеих группах выявлено следующее. Высокую пленкообразующую способность чаще проявляли клинические изоляты (81.8%). Основную часть почвенных бактерий составили штаммы, «склонные к адгезии» (таблица).
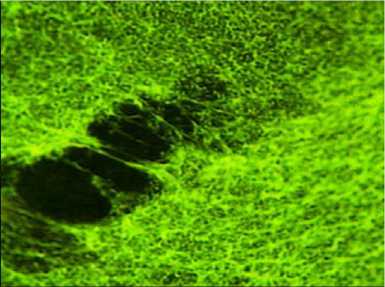
Биопленка, образованная штаммом P. aeruginosa 3–4 (в инвертированном варианте, увеличение 1×1500)
Отмечено, что как гидрофильные по BATH-тесту штаммы P. aeruginosa (в фазу гексадекана из водной фазы экстрагировалось не более 5–8% клеток), так и более гидрофобные псевдомонады (экстрагировалось до 24.2%) проявляли высокую пленкообразующую способность, т.е. не было обнаружено четкого взаимного соответствия между гидрофильно-гидрофобными свойствами клеточной стенки бактерий и степенью пленкообразова-ния. Учитывая, что псевдомонады разных видов подвергаются расщеплению на диссоцианты R-, S- и M-типов (Милько, Мартынкина, 1996), изучаемые штаммы были разделены по доминирую-щиму морфотипу колоний при росте на LB-агаре. Различные представители рода Pseudomonas формировали колонии преимущественно S-типа. Среди почвенных штаммов выявлены все три варианта, в то время как R-штаммы в группе клинических изолятов отсутствовали.
Прослежена взаимосвязь анализируемых показателей среди штаммов с разными морфотипами колоний. Оказалось, что гидрофобность клеток S-штаммов варьировала в широких пределах: выявлены как максимальные для данной выборки, так и нулевые показатели коэффициента гидрофобности. Клетки М-вариантов практически не переходили из суспензии в гексадекан (M±σ – 3.6±2.5%). Бактерии R-типа, изолированные из почвы, устойчиво проявляли гидрофобные свойства с минимальным разбросом показателей коэффициента гидрофобности. Важно подчеркнуть, что только для R-штаммов отмечена прямая зависимость между показателем гидрофобности и пленкообразо-ванием. В то же время, клинические штаммы с М-морфотипом колоний и низким коэффициентом гидрофобности все были отнесены к пленкообразующим. Как почвенные, так и клинические S-изоляты были вариабельны по обоим признакам.
Заключение
Псевдомонады относятся к убиквитарным микроорганизмам, которые отличает популяционная гетерогенность, обусловливающая возможность вида выжить практически в любых местах обитания. Такая вариабельность выявлена и по гидрофобности клеточной поверхности, и по пленкообразующей способности в обеих изученных группах, хотя у почвенных изолятов средние отклонения этих показателей были минимальными. По-видимому, это связано с относительно более постоянными условиями обитания бактерий в природных экосистемах, где отсутствуют агрессивные факторы, имеющие место в условиях ЛПУ (антибиотики, дезинфектанты, смена хозяина и др.).
Как показали наши исследования, закрепление этих микроорганизмов на поверхности и ее дальнейшая колонизация далеко не всегда детерминируется гидрофобностью клеток. Очевидно, что относительно низкую гидрофобность клеточной по- верхности у псевдомонад могут компенсировать P. aeruginosa и т.п. Это объясняет высокую плен-другие факторы адгезии, в том числе пили, жгути- кообразующую способность М- и S-вариантов. ки, альгинат, гемагглютинин, экзоэнзим S – для
Гидрофобные свойства и пленкообразующая способность штаммов рода Pseudomonas
|
Группа |
Штамм |
Морфотип колонии |
Показатель гидрофобности, % М±m |
Пленкообразующая способность, ОП 580 М±m |
Место и год выделения |
|
I |
А51 |
S |
9.2±1.2 |
0.728 ± 0.116 |
Солонец, Троицк, Челябинская обл., 2009 |
|
А59 |
S |
10.5 ± 0.5 |
0.593 ± 0.096 |
Там же |
|
|
А60 |
R |
10.3 ± 1.1 |
1.120 ± 0.088 |
Там же |
|
|
А61 |
S |
4.5 ± 0.8 |
1.401 ± 0.303 |
Там же |
|
|
А66 |
M |
5.7 ± 0.6 |
0.898 ± 0.061 |
Там же |
|
|
H7 |
R |
13.7 ± 1.5 |
1.031 ± 0.075 |
Каштановая почва, Саратовская обл. |
|
|
H8 |
S |
2.7 ± 1.1 |
0.974 ± 0.078 |
Там же |
|
|
H12 |
M |
2.9 ± 0.6 |
0.473 ± 0.078 |
Там же |
|
|
H13 |
M |
2.0 ± 0.5 |
0.532 ± 0.069 |
Там же |
|
|
H15 |
S |
6.0 ± 0.9 |
0.464 ± 0.069 |
Там же |
|
|
H16 |
S |
4.4 ± 0.4 |
0.654 ± 0.080 |
Там же |
|
|
Н17 |
R |
10.0 ± 1.2 |
1.004 ± 0.098 |
Там же |
|
|
Н18 |
S |
4.1 ± 0.4 |
0.700 ± 0.046 |
Там же |
|
|
Н19 |
S |
0.0 ± 0 |
0.691 ± 0.084 |
Там же |
|
|
Н20 |
М |
6.6 ± 0.7 |
1.056 ± 0.126 |
Там же |
|
|
Н31 |
М |
1.2 ± 0.3 |
1.037 ± 0.275 |
Там же |
|
|
II |
3-4 |
M |
0.0 ± 0 |
2.269 ± 0.106 |
ГКБ, Пермь, 2009 |
|
3-19 |
M |
0.6 ± 0.3 |
1.916 ± 0.087 |
Там же |
|
|
3-24 |
S |
0.0 ± 0 |
0.642 ± 0.059 |
Там же |
|
|
8-4 |
M |
5.3 ± 0.5 |
1.199 ± 0.044 |
Там же |
|
|
2пл |
S |
5.7 ± 0.8 |
0.992 ± 0.129 |
ЦРБ, п. Лобаново, 2010 |
|
|
31пл |
S |
7.5 ± 0.8 |
0.381 ± 0.015 |
Там же |
|
|
60пл |
S |
14.8 ± 1.6 |
1.739 ± 0.089 |
Там же |
|
|
9 |
S |
21.2 ± 1.9 |
1.113 ± 0.087 |
ГКБ, Чайковский, 2010 |
|
|
17 |
M |
8.2 ± 0.7 |
0.904 ± 0.093 |
Там же |
|
|
18 |
S |
17.3 ± 2.7 |
1.108 ± 0.074 |
Там же |
|
|
К-7 |
S |
24.2 ± 1.8 |
1.171 ± 0.104 |
ККБ, Пермь, 2009 |
Штаммоспецифичность показателя гидрофобности и независимо от этого высокая пленкообразующая способность клинических штаммов (особенно М-типа) еще раз указывает на высочайшую пластичность P. aeruginosae, способных устойчиво сохранять наиболее выгодный в конкретной экологической нише признак. Вместе с тем, можно полагать, что у природных штаммов доминируют механизмы неспецифической адгезии, а у клинических – фактором, определяющим колонизацию и пленкообразование в биотопах макроорганизма, служит специфическая адгезия. Сочетание этих двух механизмов обеспечивает выживание псевдомонад в нозокомиальных условиях.
Список литературы Гидрофобные свойства и пленкообразующая способность штаммов рода Pseudomonas, изолированных из разных экологических ниш
- Бухарин, О.В., Механизмы выживания бактерий/О.В. Бухарин, А.Л. Гинцбург, Ю.М. Романова [и др.]. М.: Медицина, 2005. 367 с.
- Ильина, Т.С. Системы коммуникаций у бактерий и их роль в патогенности/Т.С. Ильина, Ю.М. Романова, А.Л Гинцбург//Микробиология. 2006. № 3. С. 22-29.
- Коваленко, Г.А. Углеродминеральные носители для адсорбционной иммобилизации нерастущих бактериальных клеток/Г.А. Коваленко Е.В. Кузнецова, В.М Ленская//Биотехнология. 1998. № 1. С. 47-56.
- Милько, Е.С. Гидрофильно-гидрофобные и адгезивные свойства диссоциантов Rhodococcus rubropertinctus/Е.С. Милько, Н.С. Егоров//Микробиология. 1994. Т. 64, Вып. 2. С. 382-284.
- Милько, Е.С. Морфологические и физиолого-биохимические особенности диссоциантов Pseudomonas aeruginosae/Е.С. Милько, Л.П. Мартынкина//Микробиология. 1996. Т. 65, № 3. С. 352-356.
- Николаев, Ю.А. Биопленка -«город микробов» или аналог многоклеточного организма?/Ю.А. Николаев, В.К. Плакунов//Микробиология. 2007. Т. 76, № 2. С. 149-163.
- Радионович, А.М. Биофильм и его потенциальная роль в патогенезе хронического бронхолегочного процесса у больных муковисцедозом, инфицированных Pseudomonas aeruginosae/А.М. Радионович, Н.Ю. Каширская, Н.И. Капранов//Пульмонология. Прил. по муковисцидозу. 2006. С. 106-112.
- Сидоренко, С.В. Роль бактериальных биопленок в патологии человека//Инф. хир. 2004. Т. 2, № 3. С. 16-20.
- Шагинян, И.А., Формирование биопленок клиническими штаммами бактерий комплекса Burkholderia cepacia в зависимости от их фенотипических и генотипических характеристик/И.А. Шагинян, Г.А. Данилина, М.Ю. Чернуха [и др.]//Журн. микробиол. 2007. № 1. С. 3-9.
- Costerton, J.W. Bacterial biofilms: a common cause of persistent infections/J.W. Costerton, P.S. Stewart, E.P. Greenberg//Science. 1999. Vol. 284. P. 1318-1322.
- Rosenberg, M. Adherence of bacteria to hydrocarbons: A simple method for measuring cell surface hydrophobicity/M. Rosenberg, D. Gutnik, E. Rosenberg//FEMS Microbiol. Lett. 1980. Vol. 9. P. 29-33.
