Гистологические характеристики нейронов неокортекса и гиппокампа крыс в позднем онтогенезе

Автор: Бонь Е.И., Аладьева Т.Л.
Журнал: Тюменский медицинский журнал @tmjournal
Статья в выпуске: 1 т.22, 2020 года.
Бесплатный доступ
Цель работы - сравнительное изучение гистологических характеристик пирамидных нейронов гиппокампа, лобной и теменной фронтальной коры головного мозга 2,5-летних крыс. Эксперименты выполнены на 12 самцах беспородных белых крыс с начальной массой 270 ± 20 г. Забор материала осуществлялся спустя 2,5 года после рождения. На поздних этапах постнатального онтогенеза в нейронах происходят деструктивные изменения, проявляющиеся на гистологическом уровне уменьшением размеров и деформацией перикарионов, гиперхроматозом, гипохроматозом и сморщиванием клеток. Наиболее явны выявленные нарушения в неокортексе, наименее - в гиппокампе.
Онтогенез, старение, неокортекс, гиппокамп, нейроны
Короткий адрес: https://sciup.org/140303366
IDR: 140303366 | УДК: 616.89-008.441.13:618.33 | DOI: 10.36361/2307-4698-2020-22-1-46-49
Histological characteristics of neurons of rats neocortex and hippocampus in late ontogenesis
The purpose of the work is a comparative study of the histological characteristics of the pyramidal neurons of the hippocampus, frontal and parietal frontal cortex of 2.5-year-old rats. The experiments were performed on 12 males of outbred white rats with an initial weight of 270 ± 20 g. Material was taken 2.5 years after birth. At the late stages of postnatal ontogenesis, destructive changes occur in neurons, which are manifested at the histological level by a decrease in size and deformation of the pericaryons, hyperchromatosis, hypochromatosis and wrinkling of cells. The most obvious violations in the parietal cortex, the least in the hippocampus.
Текст научной статьи Гистологические характеристики нейронов неокортекса и гиппокампа крыс в позднем онтогенезе
Актуальность. Возрастная перестройка коры больших полушарий головного мозга происходит в течение всей жизни, причем в раннем постнатальном периоде преобладают процессы пролиферации и дифференцировки нервных элементов с усложнением их структуры, а в период старения – инволюционные процессы [4, 9, 13]. Вообще, возрастные изменения нейронов изучены недостаточно. Особенно их старческая инволюция в филогенетически разных отделах коры головного мозга.
Крыса часто используется в качестве объекта экспериментов по изучению головного мозга, что связано со схожестью цитоархитектоники их мозга с человеческим [1, 3]. Средняя продолжительность жизни крысы составляет 2-3 года. Для изучения гистологических особенностей стареющих нейронов избрали срок 2,5 года. В качестве предмета исследования были выбраны такие филогенетически разные отделы коры головного мозга крысы как гиппокамп, относящийся к старой (палеокортекс) коре и различные области не-окортекса (лобная и теменная кора) [1, 3, 7].
Гиппокамп обеспечивает пространственную ориентацию и запоминание определенных мест в пространстве, играет важную роль в обонятельных реакциях, а именно в запоминании запахов, а также в обеспечении процессов обучения и памяти [1, 7].
В области лобной коры находится центральный отдел анализатора кинестетических раздражений, исходящих от костей, суставов, скелетных мышц и их сухожилий. Здесь замыкаются двигательные условные рефлексы, находится ядро двигательного анали- затора, имеющего отношение к сочетанному повороту головы и глаз в противоположную сторону и ядро осязательной, болевой и температурной чувствительности, происходит анализ импульсов, идущих от внутренних органов и сосудов.
Одной из функций лобной коры является формирование доминирующей мотивации, участие в регуляции целенаправленной селекции действий, социальном взаимодействии и выборе линии поведения [1, 3, 7].
Теменная кора снабжена системой эфферентных путей как к другим, корковым формациям рострального полюса больших полушарий, так и к подкорковым образованиям: ядрам таламуса, гипоталамусу, ядрам экстрапирамидной системы, а также к спинному мозгу по путям пирамидного тракта. В ней находится центральная часть двигательного анализатора, который регулирует целенаправленные комбинированные движения, выработанные практикой индивидуальной жизни [1, 3, 7].
Целью настоящей работы было сравнительное изучение гистологических характеристик пирамидных нейронов гиппокампа, лобной и теменной фронтальной коры головного мозга 2,5-летних крыс.
Методика эксперимента. Эксперименты выполнены на 12 самцах беспородных белых крыс с начальной массой 270 ± 20 г. Все опыты проведены с учетом «правил проведения работ с использованием экспериментальных животных». На данное исследование получено разрешение комитета по биомедицинской этике Гродненского государственного медицинского университета. Животные находились на стандартном
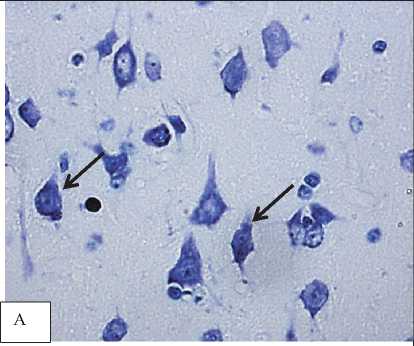
Рис. 1. Гиперхромные сморщенные нейроны в лобной (А) и теменной (Б) коре 2,5-летних крыс. Обозначены стрелками. Окраска по Нисслю. Цифровая микрофотография. Увеличение 400
Изучение гистологических препаратов и их микрофотографирование проводили с помощью микроскопа Axioscop 2 plus (Zeiss, Германия), цифровой видеокамеры (LeicaDFC 320, Германия) и программы анализа изображения ImageWarp (Bitflow, США). Расположение гиппокампа, лобной
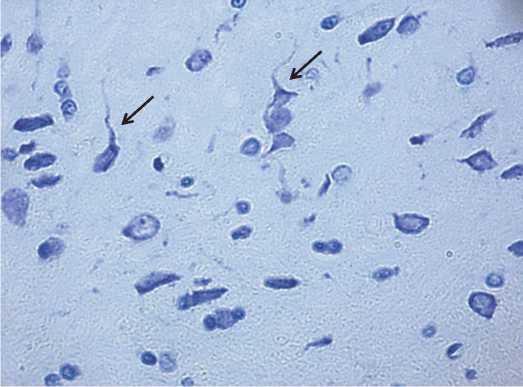
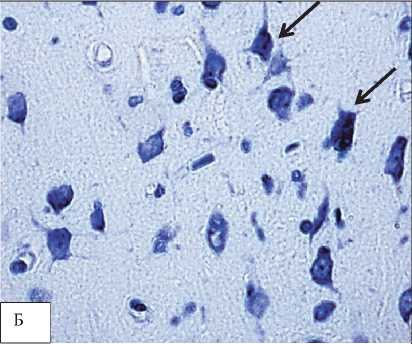
Рис. 2. Деформация перикарионов и нейропиля нейронов теменной коры 2,5-летних крыс. Обозначено стрелками. Окраска по Нисслю. Цифровая микрофотография. Увеличение 400
и теменной коры в гистологических препаратах мозга крыс определяли с помощью стереотаксического атласа [16].
Результаты и обсуждение. В неокортексе 2,5-летних крыс преобладают гиперхромные сморщенные нейроны (рис. 1).
Этот тип нейронов часто встречается при церебральной ишемии. Интенсивная окраска цитоплазмы объясняется конденсацией содержимого в связи со сморщиванием перикариона вследствие водно-электролитных нарушений. Гиперхромные сморщенные нейронырасцениваютсякакдеструктивно-измененные клетки, их функциональная активность значительно снижена. При электронной микроскопии установлено, что их митохондрии лишены крист, а органеллы синтеза, такие как эндоплазматический ретикулум и комплекс Гольджи, фрагментированы и дезорганизованы [2, 5, 6, 10, 11, 12, 14, 15]. Следственно, в гиперхромных нейронах нарушены энергетические и синтетические
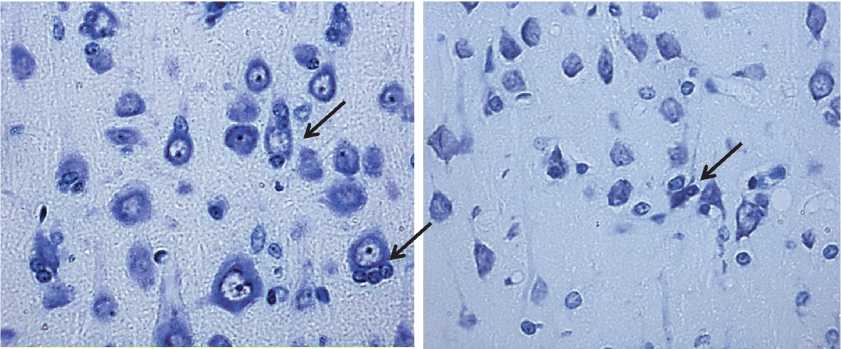
Рис. 3. Сателлитоз и нейронофагия нейронов лобной (А) и теменной (Б) коры 2,5-летних крыс. Обозначено стрелками. Окраска по Нисслю. Цифровая микрофотография. Увеличение 400
процессы, необходимые для образования медиаторов и передачи нервного импульса.
В нейронах не-окортекса также наблюдалась деформация нейропиля, связанная, скорее всего, с нарушением цитоскелета нейронов (рис. 2).
Деструктивные формы нейронов с нарушенным метаболизмом элими- рационе вивария [8]. Забор материала осуществлялся спустя 2,5 года после рождения. После декапитации быстро извлекали головной мозг, кусочки переднего отдела коры больших полушарий фиксировали в жидкости Карнуа. Серийные парафиновые срезы окрашивали 0,1% толуидиновым синим по методу Ниссля.
нируются клетками микроглии. На гистологических препаратах были выявлены признаки сателлитоза, при котором клетки глии располагаются на поверхности нейрона и нейронофагии (проникновение клеток глии в тело погибающего нейрона) [10], рисунок 3.
Кроме гиперхромных сморщенных нейронов, были обнаружены и гипохромные сморщенные клетки (рис. 4).
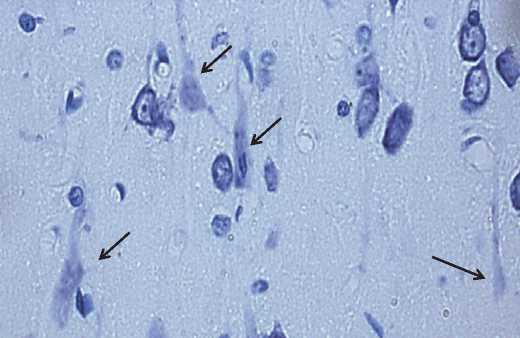
Рис. 4. Гипохромные сморщенные нейроны теменной коры 2,5-летних крыс. Обозначены стрелками. Окраска по Нисслю. Цифровая микрофотография. Увеличение 400
Бледное окрашивание их цитоплазмы тионином по методу Ниссля связано, возможно, с распадом цистерн гранулярной эндоплазматической сети. Данный тип нейронов образуется либо из гиперхромных сморщенных нейронов при их дальнейшей инволюции, либо из гипохромных клеток.
В гиппокампе изменения были минимальны. В популяции преобладали нормохромные нейроны, гиперхромные сморщенные были единичными (указаны стрелками на рисунке 5). В предыдущих наших исследоваях по изучению ишемии головного мозга, также была установлена устойчивость гиппокампа, как филогенетически более древней структуры, к гипоксии [5].
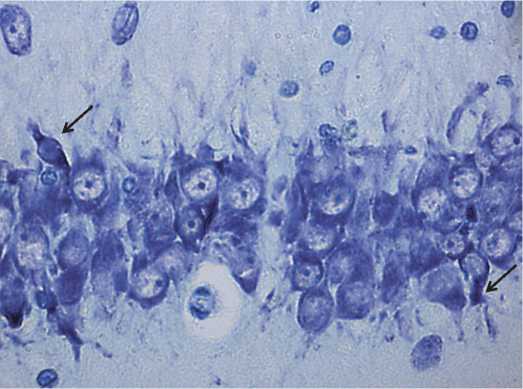
Рис. 5. Нейроны гиппокампа 2,5-летних крыс. Стрелками указаны гиперхромные сморщенные нейроны. Окраска по Нис-слю. Цифровая микрофотография. Увеличение 400
Заключение. Таким образом, на поздних этапах постнатального онтогенеза в нейронах происходят деструктивные изменения, проявляющиеся на гистологическом уровне уменьшением размеров и дефор- мацией перикарионов, гиперхроматозом, гипохрома-тозом и сморщиванием клеток.
Выявленные морфологические признаки лежат в основе ряда неврологических нарушений и снижения когнитивных процессов, наблюдаемых при старении организма. Наиболее явны выявленные нарушения в неокортексе, наименее – в гиппокампе.
Список литературы Гистологические характеристики нейронов неокортекса и гиппокампа крыс в позднем онтогенезе
- Бонь Е. И., Зиматкин С. М. Анатомические особенности коры мозга крысы // Новости медико-биологических наук. – 2016. – Т. 14. – № 4. – С. 49-54.
- Бонь Е. И., Зиматкин С. М. Изменения хроматофилии цитоплазмы больших пирамидных нейронов новой коры мозга крысы в постнатальном онтогенезе // Вестник Смоленской государственной медицинской академии. – 2019. – № 1. – С. 10-16.
- Бонь Е. И., Зиматкин С. М. Микроскопическая организация изокортекса крысы // Новости медико-биологических наук. – 2017. – № 4. – С. 80-88.
- Бонь Е. И., Зиматкин С. М. Онтогенез коры головного мозга крысы // Новости медико-биологических наук. – 2014. – № 4. – С.238-244.
- Бонь Е. И., Максимович Н. Е., Зиматкин С. М. Гистологические изменения нейронов теменной коры головного мозга крыс с субтотальной и тотальной ишемией // Вестник Смоленской государственной медицинской академии. – 2020. – Т. 19. № 2. – С. 23-27.
- Зиматкин С. М. Темные нейроны мозга / С. М. Зиматкин, Е. И. Бонь // Морфология. – 2017. – Т. 152, № 6. – С.81-86.
- Зиматкин С. М., Бонь Е. И. Строение и развитие коры головного мозга крысы: монография. – Гродно, ГрГМУ, 2019. – 155 с.
- Каркищенко Н. Н., Грачева С. В. Руководство по лабораторным животным и альтернативным моделям в биомедицинских исследованиях. – М.: Профиль-2С. – 2010, 241 с.
- Оленев С. Н. Развивающийся мозг. – Л.: Наука. – 1978. – 220 с.
- Попова Э. Н. Морфология приспособительных изменений нервных структур: монография. – Москва. Изд. Медицина. – 1976. – С. 62-82.
- Gallyas F. Novel cell-biological ideas deducible from morphological observations on “dark” neurons revisited // Ideggyogy.Sz. – 2007. – V.78. – P. 212-222.
- Ishida K., Shimizu H., Hida H., Urakawa S. Argyrophilic dark neurons represent various states of neuronal damage in brain insults: some come to die and others survive // Neuroscience. – 2004. – V. 125. – P. 633-644.
- Jellinger K. A. Cell death mechanisms in neurodegeneration // Cell. mol. med. – 2001. – Vol. 5, № 1. – P 1-17.
- Kovesdi E., Pal J, Gallyas F. The fate of “dark” neurons produced by transient focal cerebral ischemia in a nonnecrotic and non-excitotoxic environment: neurobiological aspects // Brain Res. – 2007. – V. 1147. – P. 272-283.
- Ooigawa H., Nawashiro H., Fukui S., Otani N., Osumi A. The fate of Nissl-stained dark neurons following traumatic brain injury in rats: difference between neocortex and hippocampus regarding survival rate // Acta Neuropathol. – 2006. – V. 112. – P. 471-481.
- Paxinos G., Watson C. The Rat Brain in stereotaxic coordinates. – Academic Press, Australia, 1998. – 242 p.
