Гистоморфометрические характеристики популяций ганглионарных нейронов в отдаленный период после нейротомии и восстановительной операции у собак

Автор: Шевцов В.И., Щудло Н.А., Борисова И.В., Щудло М.М., Панфилов Р.В., Варсегова Т.Н.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 2, 2005 года.
Бесплатный доступ
У двух взрослых беспородных собак выполнено пересечение седалищного нерва на уровне средней трети бедра и первичная микрохирургическая нейрорафия. Через 12 месяцев после операции методами световой микроскопии и компьютерной морфометрии изучены нейроны их ганглиев L5, L6, L7 в сопоставлении с аналогичными данными по четырем неоперированным собакам. В результате исследований отработана методика типирования А и B-нейронов в криостатных срезах, уточнены светооптические качественные характеристики А1 и А2-подтипов, определены размерные диапазоны типов ганглионарных нейронов и получены данные о соотношении A и B-типов в поясничных ганглиях интактных собак. У оперированных животных в популяции А-нейронов средние диаметры ядра и ядрышка меньше интактных на 4 % и на 2 % соответственно, однако на 19 % (статистически достоверно) уменьшена средняя площадь перикарионов. В популяции B достоверно, но незначительно (на 10 %) уменьшена средняя площадь перикарионов, а средние диаметры ядра и ядрышка уменьшены на 8 % (p
Чувствительные ганглии, a и b-нейроны, аксотомия, регенерация нерва
Короткий адрес: https://sciup.org/142120808
IDR: 142120808
The histomorphometric characteristics of ganglionary neuron populations in the long-term period after canine neurotomy and restorative surgery
Neurotomy of the sciatic nerve at the level of the femoral middle third and primary microsurgical neurorrhaphy were performed in 2 adult mongrel dogs. The neurons of their ganglia L5, L6, L7 were studied 12 months after surgery in comparison with the similar data of 4 non-operated dogs using the methods of light microscopy and computer morphometry. The study resulted in perfecting the technique of neurons A and B typing in cryostat sections, verifying the light-and-optical qualitative characteristics of A1 and A2 subtypes, determining the size ranges of the types of ganglionary neurons and obtaining the data about the ratio of A and B types in the spinal ganglia of intact dogs. As for the animals operated on, in their population of A-neurons the mean diameters of nucleus and nucleolus were 4% and 2% less (respectively) than those of intact animals, however, the mean area of perikaryons was 19% less (statistically reliably). In B population the mean area of perikaryons was reliably but slightly (10%) less and the mean diameters of nucleus and nucleolus were 8% less (p
Текст научной статьи Гистоморфометрические характеристики популяций ганглионарных нейронов в отдаленный период после нейротомии и восстановительной операции у собак
По данным последних лет [5], только в Европе несколько сотен тысяч человек в год получают повреждения периферических нервов. Несмотря на высокий уровень хирургической техники восстановительных операций в европейских странах, результативность их нередко раз- очаровывает врачей и пациентов. Многие авторы считают, что основной причиной неудовлетворительного восстановления чувствительности является посттравматический апоптоз первичных афферентных нейронов. По мнению M.J. Groves et al. [4], после периферической ак- сотомии количество чувствительных нейронов в спинномозговых узлах уменьшается не более, чем на 20 %; другие авторы [12] приводят существенно отличающиеся данные: через 32 недели после повреждения седалищного нерва крыс погибает около 37 % чувствительных нейронов. Независимо от уровня периферической аксото-мии гибель ганглионарных нейронов B-типа начинается раньше и протекает значительно интенсивнее [12, 13]. Сведения о морфофункциональных характеристиках выживших нейронов весьма противоречивы. По мнению M.R. Raffe [11], с 10-го по 20-й день после повреждения нерва регенерационная гипертрофия биосинтетического аппарата приводит к увеличению размеров нейронов. По данным T. Tan-drup et al. [12], нейроны обоих типов уменьшаются в размерах, но только на отдельных сроках наблюдения это изменение статистически значимо.
Сказанное определяет актуальность изучения морфофункциональных характеристик выживших нейронов у разных видов животных в отдалённый период регенерации после периферической аксотомии и восстановительной операции. С этой целью нами выполнено пилотное исследование на шести взрослых беспородных собаках в возрасте от 1 до 3 лет.
МАТЕРИАЛЫ И МЕТОДЫ
У двух собак под внутривенным барбитуровым наркозом выполнено пересечение седалищного нерва на уровне средней трети бедра и первичная микрохирургическая эпиневральная нейрорафия. Материал для морфологического исследования (спинальные ганглии L5-L7 оперированной и контралатеральной стороны) взят через 12 месяцев после операции. В качестве контроля исследованы ганглии четырёх неоперированных собак. Животных выводили из опыта передозировкой барбитуратов, в процессе ламинэктомии применялась префиксация in situ, а после иссечения материала – погружная фиксация охлажденной смесью 2 % растворов глутарового и параформальдегидов на фосфатном буфере (pH 7,4) с добавлением 0,1 % пикриновой кислоты. На следующие сутки после эвтаназии ганглии препарировали микрохирургическими инструментами фирмы Aesculap под бинокулярным стереомикроскопом и острым лезвием вырезали кусочки для криотомирования. Срезы толщиной 10 мкм окрашивали толуидиновым синим по методикам Романовского-Гимза, Живаго и Эйнарсона, небольшую часть материала от интактных животных им-прегнировали серебром и докрашивали гематоксилином. Препараты изучали и фотографировали, используя большие исследовательские микроскопы фирмы «Opton» (ФРГ). Для оцифровки изображений и количественного исследования использовали аппаратно-программный комплекс “ДиаМорф” (Москва). С каждого ганглия исследовали от 6 до 12 ортотропных срезов. Морфо-метрировали изображения всех ядрышкосодержащих профилей нейронов с хорошо выраженным контуром кариолеммы (инструментальное увеличение 500 ). Статистические гипотезы проверяли применением точного критерия Фишера в дольном выражении с использованием преобразования Стирлинга для подсчёта искомой вероятности, двухвыборочного t-теста, предоставляемого программой Microsoft Excel 97, непараметрического W-теста Вилкоксона для независимых выборок с помощью NP-модуля программного обеспечения анализа и обработки данных AtteStat.
РЕЗУЛЬТАТЫ
В процессе микроскопии полученного материала установлено, что наиболее элективное выявление телец Ниссля и структур ядра в криостатных срезах чувствительных нейронов удаётся при окраске толуидиновым синим. Обращая внимание на характер базофильного окрашивания нейроплазмы у интактных и оперированных собак (рис. 1), удаётся уверенно выделить клетки типа А – в соответствии с критериями, известными из литературы [7, 8, 12, 13]; прочие клетки с более интенсивной, пылевидной или гомогенной базофилией мы отнесли к типу B.
У двух интактных животных типы А и B имели чёткое разграничение по размерным характеристикам (например, в ганглии L7 к типу А можно было отнести абсолютно все нейроны более 46 мкм, а в ганглии L5 – более 50 мкм в диаметре). У двух других интактных и у обеих оперированных собак от 3 до 5 % нейронов подвергаются перекалибровке. Как правило, в таких нейронах можно отметить признаки реактивных или компенсаторно-приспособительных изменений: например, некоторые нейроны типа B попадают в размерный диапазон А-клеток при увеличении площади перикариона (внутриклеточной гипертрофии) либо отдельные нейроны типа А с ангулярными контурами и признаками конденсации цитоплазмы оказываются в размерном диапазоне B. В интактном материале среди нейронов типа А обнаруживались клетки с относительно равномерным расположением базофильных телец по площади среза клетки (рис. 2, А) либо с преимущественно перинукле- арным расположением и практически полным отсутствием их по периферии цитона (рис. 2, Б). При импрегнации серебром выявлялись соответствующие им разновидности с равномерным распределением металлофилии (рис. 2, В) и усилением металлофилии по периферии цитона (рис. 2, Г).
У животных опытной группы в ганглиях оперированной стороны практически не встречались нейроны с отсутствием базофилии по периферии клетки.
У интактных собак доли А и В-клеток в ганглии L6 также практически равны, однако в ганглии L7 и особенно L5 выше доля B-клеток (табл. 1). Относительное содержание А и В-клеток в контралатеральных ганглиях оперированных животных мало отличается от интактных. На оперированной стороне в ганглиях L5 и L7 снижена доля B-клеток, а в ганглии L6 – доля A-клеток.
Применение точного критерия Фишера для определения статистической значимости отличий параметров интактных и оперированных собак даёт p>0,05 для ганглиев L5 и L6 и p ≈ 0 для ганглия L7. То есть, только для ганглия L7 статистически высокозначимо отличие относительного содержания A и B-клеток. С большой долей вероятности это можно связать с тем, что у собак именно ганглий L7 содержит нейроны, относящиеся исключительно к пулу седалищного нерва. В L5 и L6 присутствуют нейроны ещё и бедренного нерва, который в данных опытах не был повреждён и оперирован.
Сопоставим морфометрические характеристики А и B-клеток седьмого поясничного ганглия у интактных и оперированных собак (табл. 2). В популяции А средние диаметры ядра и ядрышка меньше интактных всего на 4 % и на 2 % соответственно – статистически незначимо, однако на 19 % (статистически достоверно) уменьшена средняя площадь перикарионов. В популяции B хотя и достоверно, но весьма незначительно (на 10 %) уменьшена средняя площадь перикарионов, а средние диаметры ядра и ядрышка уменьшены на 8 % (p < 0,05).
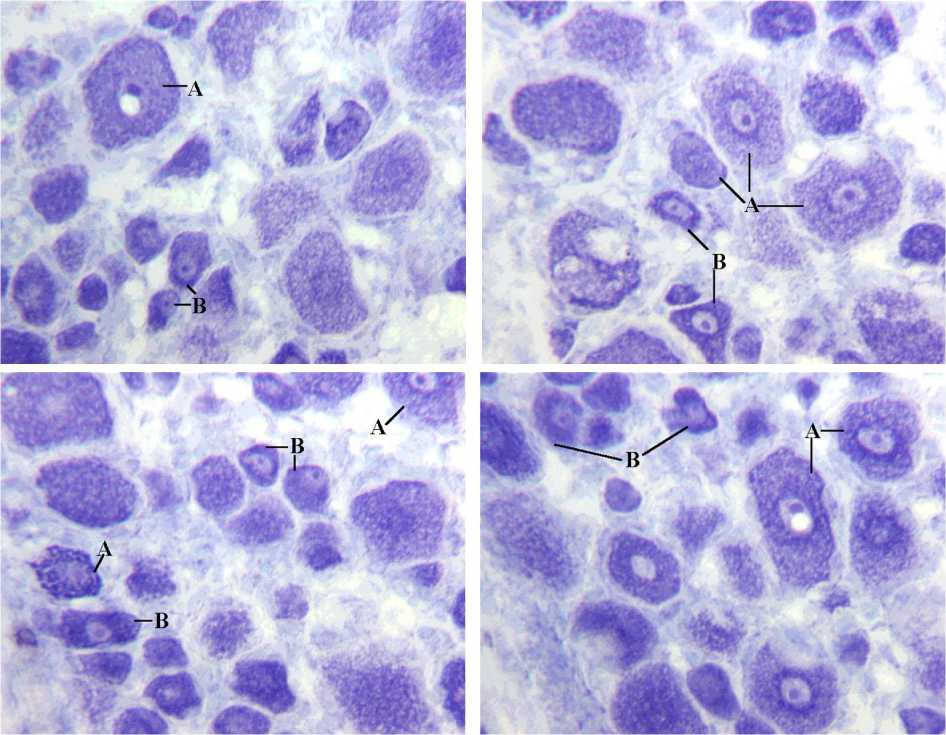
Рис. 1. Типирование нейронов в ганглии L6 оперированной стороны. 12 месяцев после нейротомии и микрохирургической нейрорафии седалищного нерва. Продольный криостатный срез. Окраска толуидиновым синим. А нейроны типа А; B нейроны типа B. Инструментальное увеличение 200
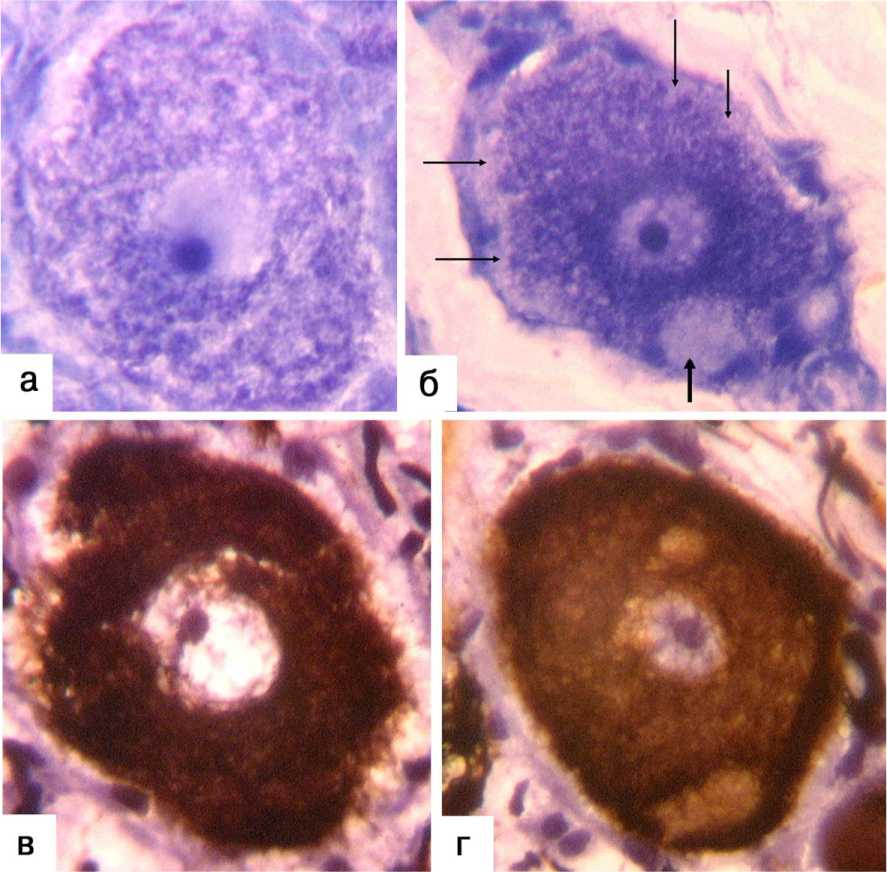
Рис. 2. Варианты распределения базофилии и металлофилии в ганглионарных А-клетках интактных собак. А нейрон с равномерным распределением базофильных телец в цитоплазме и эксцентрично расположенным ядрышком. Б базофилия выражена в основном перинуклеарно и отсутствует по периферии клетки (тонкие стрелки); "очаговый периферический хроматолиз" (толстая стрелка). В нейрон с равномерным распределением металлофилии; Г нейрон с усилением метал-лофилии по периферии цитона. Криостатные срезы, окраска толуидиновым синим ( А, Б ), импрегнация серебром с докра-ской гематоксилином ( В, Г ). Инструментальное увеличение 500
Таблица 1 Относительное содержание нейронов А и B-типов в поясничных ганглиях с репрезентацией афферентов седалищного нерва у интактных и оперированных животных
|
Группа ганглиев |
Ганглий |
|||||
|
L5 |
L6 |
L7 |
||||
|
A |
B |
A |
B |
A |
B |
|
|
Интактных животных |
38,6% |
61,4% |
51,9% |
48,1% |
43,1% |
56,9% |
|
Оперированной стороны |
42,2% |
57,8% |
46,6% |
53,4% |
60,2% |
39,8% |
|
Контралатеральные |
34,5% |
65,5% |
51,2% |
48,8% |
- |
- |
|
Результат тетрахорического тестирования параметров интактных и оперированных собак |
p ≈ 0,100259 |
p ≈ 0,201897 |
p ≈ 0 |
|||
Таблица 2
Планиметрические характеристики нейронов ганглия L7 у оперированных и интактных животных
|
Группа |
Средний диаметр нейрона |
Средний диаметр ядра |
Средний диаметр ядрышка |
Средняя площадь перикариона |
||||
|
А |
B |
A |
B |
A |
B |
A |
B |
|
|
Опер |
61,6±1,3 |
30,9±1,1 |
23,6±0,5 |
15,1±0,4 |
6,2±0,1 |
4,4±0,1 |
2634±120 |
608±52 |
|
Интакт |
67,7±2,2 |
33,3±0,07 |
24,6±0,6 |
16,5±0,3 |
6,3±0,1 |
4,8±0,1 |
3253±232 |
677±31 |
|
Опер/интакт 100% |
91 % |
93 % |
96 % |
92 % |
98 % |
92 % |
81 % |
90 % |
|
Интервал p-значения |
<0,05 |
<0,05 |
>0,05 |
<0,05 |
>0,05 |
<0,05 |
<0,05 |
<0,05 |
Данные таблицы 3 позволяют видеть, что популяция B-клеток по частоте качественных изменений мало отличается от интактной: наиболее значительно отличие по признаку "частота ангулярных контуров цитона", но даже оно оказывается статистически недостоверным (по результатам точного теста Фишера p ≈ 0,135335). Однако повышение, хотя и статистически недостоверное, частоты эктопии ядер и статистически значимое увеличение частоты эксцентричных ядрышек в А-нейронах оперированных животных вероятно следует трактовать как реактивное изменение.
При микроскопии А-нейронов с эктопированными ядрами создаётся впечатление увеличения размеров цитона, декомпозиции базофильных телец в его центре. Эктопированные ядра, как правило, менее конденсированы по сравнению с центрально расположенными, а
Таблица 3
Частота структурных особенностей и реактивных изменений в разных типах нейронов спинального ганглиия L7 у интактных и оперированных собак
|
Признак |
Неоперированные собаки |
Опыт |
||
|
А |
B |
A |
B |
|
|
Эктопия ядра |
0,9 % |
2,7 % |
1,7 % |
1,7 % |
|
Эктопия ядрышка |
5,4 % |
14,4 % |
16,1 % |
16,9 % |
|
Центральный хроматолиз |
0 |
0 |
1,7 % |
0 |
|
"Очаговый периферический хроматолиз" |
10,8 % |
0,9 % |
3,4 % |
0 |
|
Ангулярные контуры цитона |
0,9 % |
5,4 % |
5,9 % |
8,5 % |
|
Вакуолизация ядра |
0 |
0 |
2,0 % |
0,8 % |
|
Вакуолизация цитоплазмы |
1,8 % |
0,9% |
0,8 % |
1,7 % |
|
Цитоплазматические включения (липофусцин) |
2,7 % |
0 |
2,5 % |
0 |
|
Изменения формы ядра |
0 |
0,9 % |
0 |
0,8 % |
ОБСУЖДЕНИЕ
Функционально нейроны спинальных ганглиев гетерогенны, поскольку трансформируют афферентные сигналы разных рецепторных модальностей: тактильной, температурной, ноцицептивной и проприоцептивной [8]. В начале прошлого века делались попытки морфологической типизации нейронов по тинкториальным свойствам. По мнению A.R. Lieberman [7], большинство этих классификаций являются "эзотерическими", а М.В. Войно-Ясенецкий и Ю.М. Жаботинский [1] подчёркивали, что способность нейронов к окрашиванию в значительной мере определяется факторами артефициального характера и посмертными изменениями. Однако приведённое у А.А. Заварзина и С.И. Щелкунова [2] подразделение ганглионарных нейронов на светлые и тёмные было подтверждено в исследованиях J.-H. Scharf (1958) и K.H. Andres (1961), назвавших их типами A и B [цит. по 7, стр. 219]. Эта классификация получила широкое признание [8, 12, 13], поскольку было установлено, что нейроны A и B различаются не только тинкториальными свойствами и размерными характеристиками, но также деталями ультраструктуры, строением отростков, биохими- ядрышко в них часто увеличено. Совокупность перечисленных признаков укладывается в хорошо известную картину центрального хроматолиза.
Локальная декомпозиция базофильных телец ("очаговый периферический хроматолиз"), встречалась чаще в нейронах ганглия L7 интактных собак, чем оперированных, причём чаще в А-, нежели в B-клетках. Следует обратить внимание на то, что в популяции А-нейронов оперированных животных по сравнению с интактными на 5 % (p ≈ 0,033373) увеличена доля клеток с ангулярными контурами. В срезе такие клетки имеют форму треугольников, ромбов, реже шестиугольников; создаётся впечатление конденсации их цитоплазмы, некоторые из них имеют резко базофильное конденсированное ядро. Других характерных морфологических признаков апоптоза [10] встретить не удалось.
ческими свойствами.
В нашем материале на светооптическом уровне наряду с типами A и B у интактных собак различимы разновидности А-нейронов: с равномерным распределением базо- и металло-фильных структур; с ослаблением базофилии и усилением металлофилии по периферии цитона, что, по-видимому, обусловлено сосредоточением в этой зоне главным образом элементов цитоскелета. Данное предположение согласуется с выводами специального исследования О.С. Сотникова и др. [3]. Экстримы распределения базофилии в А-нейронах выявляются у других видов животных в окрашенных толуидиновым синим полутонких срезах [7]. Они соответствуют ультраструктурным подтипам А1 и А2, которые, по данным J.M. Jacobs [цит. по 7], различаются ультраструктурно по представительству и распределению аппарата Гольджи, гранулярного эндоплазматического ретикулума (ГЭР), свободных рибосом, нейрофиламентов. По данным литературы, у мышевидных грызунов [7] численные доли A и B-типов ганглионарных нейронов приблизительно равны. В на- шем материале у всех четырех интактных собак выявлены незначительные различия ганглиев L5, L6, L7 по этому параметру. Его отличия от интактного материала у двух оперированных собак однонаправленны, невелики и статистически значимы только для ганглия L7. В комплексе с данными планиметрических исследований и результатами полуколичественной оценки качественных изменений нейронов типа A и B они заставляют предположить, что, несмотря на некоторое преобладание утраты нейронов типа B, к 12 месяцам после операции этот тип нейронов обладает меньшими отличиями от интактной нормы, чем популяция А. Анализ причин изменений размерных характеристик ядер и ядрышек в B-нейронах требует дополнительных исследований на большем количестве животных. Следует также отметить, что через 12 месяцев после повреждения нерва и восстановительной операции представительство клеток, которые находятся в состоянии денервационной атрофии и необратимых реактивных изменений, более значительно в популяции А по сравнению с популяцией B. Отсутствие сопоставления с интактным материалом считается одним из главных источников неправильной интерпретации морфологических изменений нейронов [1], поэтому мы анализировали структурные особенности и реактивные изменения нейронов путём сопоставления частоты качественных признаков у оперированных и интактных собак. Это позволило дифференцированно оценить структурные особенности и реактивные изменения. О том, что тёмные нейроны с ангулярными контурами довольно-таки часто встречаются в нормальных интактных ганглиях разных видов животных, писали многие авторы [цит. по 7]. Высокая частота эктопии ядрышек в B-клетках седьмого поясничного ганглия у неоперирован-ных и оперированных собак, по-видимому, также должна интерпретироваться как их структурная особенность, а не патологическое изменение, поскольку, по данным литературы [7], в ганглиях грудного отдела мышевидных грызунов, кошек и зайцеобразных частота этого признака во всех нейронах варьирует от 50 до 100 %. В единичных работах в качестве реактивного изменения нейронов спинальных ганглиев рассматривается периферический хроматолиз. Так, частота "глобулярных и сфероидальных" хроматолитических образований, которые ультраструктурно соответствуют расширениям цистерн ГЭР, аккумуляциям нейрофиламентов и липидным каплям, выше у животных, перенесших субпериневральную инъекцию алюминийсодержащего раствора, по сравнению с интактными животными [6]. В нашем материале этот признак преобладал в популяции А у интактных собак и практически не встречался у оперированных. Вероятно, он отражает структурнофункциональную активность нормальных нейронов, их участие в самовосстановлении мембранных структур, цитоскелетных протеинов, органелл, синтезе и секреции нейротрансмиттеров. Это предположение хорошо подтверждается тем, что у нейронов с "периферическим хроматолизом", как правило, отмечается звёздообразная конфигурация хроматина в ядре, что считается [1] признаком высокой функциональной активности ядры-шка. Следовательно, снижение частоты рассматриваемого признака у оперированных животных можно трактовать как показатель сниженной структурно-функциональной активности нейронов, о чём также свидетельствует уже рассмотренное достоверное уменьшение средней площади перикариона.
ВЫВОДЫ
-
1. Изменения относительного содержания нейронов A и B типов в спинальных ганглиях L5-L7 у собак через 12 месяцев после нейротомии и восстановительной операции по сравнению с неопе-рированными животными указывают на то, что преобладание потери нейронов B-типа характерно лишь для определённого этапа регенерации.
-
2. Сопоставление частоты реактивных изменений и морфометрических характеристик нейронов A и B-типов с интактным материалом свидетельствует, что через 12 месяцев после повреждения и первичной восстановительной операции снижение структурно-функциональной активности более выражено в популяции А-клеток. Наряду с увеличением частоты премортальных изменений нейронов популяции А это заставляет предположить, что в отдалённые сроки после операции гибель B-нейронов замедляется и до-
- минирует потеря А-нейронов.
-
3. Полученные данные об изменениях выживших нейронов в отдалённые сроки после повреждения нерва и восстановительной операции формируют представление о необходимости разработки и применения более специфической и длительной, чем это принято в настоящее время, нейротропной терапии у пациентов с травмами периферических нервов.
-
4. Разработанный в данном исследовании алгоритм оценки гистоморфометрических характеристик популяций нейронов, сочетающий количественные и полуколичественные параметры, может быть эффективно использован для сравнительных исследований результативности различных экспериментально моделируемых хирургических и терапевтических стратегий в лечении травм нервных стволов.
