Гистоструктура трахеальной стенки у цыплят-бройлеров в зависимости от условий циркуляции воздуха в закрытых помещениях

Автор: Фисинин В.И., Салеева И.П., Османян А.К., Панов В.П., Малородов В.В., Черепанова Н.Г., Хамитова В.З.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Животные в условиях интенсивных технологий
Статья в выпуске: 4 т.56, 2021 года.
Бесплатный доступ
В настоящее время наблюдается интенсификация производства мяса бройлеров, в связи с чем увеличивается число исследований, посвященных оптимизации и приведению к нормативам микроклимата в помещениях для выращивания птицы. При этом уделяется недостаточно внимания воздействию параметров микроклиимата на дыхательный аппарат птиц. В представленной работе впервые показано, что циркуляция воздуха в закрытых помещениях способствует поддержанию продуктивности, улучшению гистологической структуры и гистохимического статуса трахеальной стенки у цыплят-бройлеров. Цель эксперимента - определение гистологической и гистохимической структуры трахеальной стенки у цыплят-бройлеров ( Gallusgallus domesticus ) кросса Ross 308, которые содержались в вентилируемых помещениях или при отсутствии циркуляции воздуха. Исследование было выполнено в 2020-2021 годах на птицефабрике ООО «Челны-Бройлер» (Республика Татарстан). Цыплят-бройлеров кросса Ross 308 выращивали до 39-суточного возраста в пяти закрытых производственных корпусах, которые различались по равномерности распределения воздушных потоков и циркуляции воздуха (пять групп по 35 гол. в каждой). При морфометрических исследованиях использовали 175 образцов трахей бройлеров всех групп (средняя живая масса 2500 г). Обзорные препараты (по три среза для каждого образца, всего 525 препаратов) получали окрашиванием гематоксилином и эозином. Для гистохимических исследований кислых и нейтральных муцинов срезы окрашивали комбинированным методом выявления полисахаридов с применением реакции Шифф-йодной кислоты (ШИК-реакции) и альцианового синего по методикам, рекомендованным «ООО Лабико» (Россия). В помещениях, где содержались II, III и IV контрольные группы, отсутствовала циркуляция воздуха, в залах выращивания птиц из I и V опытных групп были установлены циркуляционные осевые вентиляторы SF-550-02 («AgroKurs», Россия; производительность 8,5 тыс. м3/ч). Циркуляционные вентиляторы запускали с 10-суточного возраста бройлеров. Недостаточность циркуляции воздушных потоков в залах выращивания птиц вызывала деструктивные изменения слизистой оболочки трахеи, которые выражались в разрастании собственной пластинки слизистой, отеке, уменьшении высоты эпителия, разрушении ресничек. Это приводило к метаплазии эпителия и нарушению мукоцилиарного транспорта. Так, у птицы в I опытной группе толщина слизистой оболочки и входящей в ее состав собственной пластинки оказалась минимальной - соответственно 147,2±3,3 и 129,1±3,1 мкм, в III контрольной группе эти показатели были максимальными - 404,7±9,4 мкм и 395,7±9,4 мкм (p ≤ 0,01). Толщина эпителиального слоя трахеи была достоверно выше в I и V опытных группах (в среднем на 14 %) в сравнении со II, III и IV контрольными группами (p ≤ 0,01). Отсутствие циркуляции воздуха в помещениях привело к достоверному снижению высоты ресничек во II, III и IV группах соответственно на 39,5; 58,1 и 67,5 % по отношению к I и V группам (p ≤ 0,01). Прирост живой массы птицы в 5-недельном возрасте в I опытной группе был выше в сравнении со II, III и IV контрольными группами соответственно на 6,5; 3,2 и 7,1 % (p ≤ 0,05). Полученные гистологические препараты позволяют предположить наличие у птиц в пласте эпителия трахеи простых многоклеточных эндоэпителиальных желез. Таким образом, при обеспечении достаточной циркуляции воздушных потоков в закрытом помещении толщина слизистой оболочки и собственной пластинки уменьшается, а толщина эпителиального слоя и высота ресничек трахеи увеличиваются. Эти показатели состояния птицы могут служить индикаторами благоприятных условий содержания.
Гистоструктура трахеи, слизистая трахеи, реснитчатый эпителий, микроклимат, циркуляция воздушных потоков, респираторная система, гистохимия, шик-реакция, альциановый синий
Короткий адрес: https://sciup.org/142230924
IDR: 142230924 | УДК: 636.52/.58.033:611.21/.23:57.04:697.92 | DOI: 10.15389/agrobiology.2021.4.782rus
Histostructure of the tracheal wall of broiler chickens depending on air circulation conditions in closed poultry houses
Currently, optimization of indoor microclimatic conditions in poultry houses is attracting considerable interest due to the intensification of broiler meat production. However, given the increase in flock sizes of broilers, little research has focused on the effect of microclimate parameters in poultry houses on the bird's respiratory system. Insufficient air exchange in the premises can cause functional respiratory disorders in broiler chickens. This paper is the first to report that air circulation in closed poultry houses contributes to maintaining productivity and improves the histostructure and histochemical properties of the tracheal wall in broiler chicks ( Gallus gallus domesticus ). Our work aimed to study the influence of different air circulation regimes in closed poultry houses on histostructure and histochemical characteristic of the trachea in Ross 308 broiler chicks and their productive performance. The study was conducted in 2020-2021 at the LLC Chelny-Broiler poultry farm (Republic of Tatarstan). Ross 308 cross broiler chicks were raised until 39 days of age in five closed premises under different airflow distribution and air circulation (five groups of 35 birds each). For morphometry, 525 preparations of 175 trachea specimens from of all broilers (2500 g bodyweight) were measured. Trachea sections were stained by hematoxylin and eosin procedure. For histochemical studies of acidic and neutral mucins, sections were stained by a combined method for detecting polysaccharides using the Schiff-iodic acid (PAS-reaction) and alcian blue according to the manufacturer's recommended (LLC Labico, Russia). In the control groups 2, 3, and 4, there was no air circulation; in the experimental groups I and V, circulation was provided by forced ventilation, capacity of 8.5 thousand m3/h (SF-550-02, AgroKurs, Russia). Ventilation was run at the 10 day-age of the broiler chicks. Insufficient air circulation in the poultry rearing rooms caused destructive changes in the tracheal mucous membrane, i.e., its own lamina proliferation, edema, a decrease in the height of the epithelium, and destruction of cilia. This led to metaplasia of the epithelium and disruption of mucociliary transport. The thickness of the mucous membrane and its own lamina was minimum in the experimental group 1 (147.2±3.3 µm and 129.1±3.1 µm, respectively) and maximum in the control group 3 (404.7±9.4 µm and 395.7±9.4 µm) (p ≤ 0.01). The thickness of the tracheal epithelial layer significantly increased in the experimental groups 1 and 5 (by 14 % on average) compared to the control groups 1, 2, and 3 (p ≤ 0.01). The lack of indoor air circulation led to a significant decrease in the height of cilia in the control groups 2, 3, and 4 (by 39.5, 58.1, and 67.5 %, respectively) as compared to the experimental groups 1 and 5. The increase in birds' bodyweight at 5 weeks of age in the experimental group 1 increased compared to the control groups 2, 3, and 4 by 6.5, 3.2, and 7.1 %, respectively (p ≤ 0.05). The histochemical characteristics suggests the presence of simple multicellular endoepithelial glands in the tracheal epithelium layer of birds. Thus, with the provision of proper air circulation in an enclosed space, the thickness of the mucous membrane and its own lamina decreases, and the thickness of the epithelial layer and the height of the tracheal cilia increases. These characteristics are indicative of proper airexchange in the poultry houses.
Текст научной статьи Гистоструктура трахеальной стенки у цыплят-бройлеров в зависимости от условий циркуляции воздуха в закрытых помещениях
Одна из проблем современного индустриального сельского хозяйства — сокращение национальных генетических ресурсов пород сельскохозяйственных животных и птицы, снижение их генетического разнообразия. Подчас создается угроза не только сокращения, но и полной утраты уникальных особо ценных генофондов. Приоритетность снижения потерь генетических ресурсов, сохранения многообразия существующих локальных пород, региональных групп, типов животных и птицы, способных давать продукцию в различных условиях разведения и обеспечивающих устойчивое развитие животноводства, подтверждена международной кон- венцией о биологическом разнообразии (1).
Важный аспектом в разработке программ по сохранению пород сельскохозяйственных животных — изучение их генетических особенностей. Для оценки таких особенностей, а также паспортизации пород широко применяются мульти- и монолокусные ДНК маркеры, или микросателлиты и однонуклеотидные полиморфизмы (ОНП) (2).
Микросателлиты, как правило, высокополиморфны и включают множество аллелей на один локус. Рекомендации ФАО (Food and Agriculture Organization, FAO) по выбору микросателлитных локусов при исследовании различных видов сельскохозяйственных животных опираются на перечень (DAD-IS библиотека, , разработанный группой ISAG-FAO по генетическому разнообразию. Микросателлиты признаются информативными для анализа происхождения и картирования локусов количественных признаков (quantitative trait loci, QTL) (3, 4). В то же время с развитием технологий молекулярного тестирования все большее признание и преимущество перед ними получает анализ одно-нуклеодидных замен (single nucleotide polymorphism, SNP) с использованием микроматриц, или чипов (5).
Для сохранения и рационального использования генетических ресурсов сельскохозяйственных животных и птицы ФАО был осуществлен широкомасштабный проект по анализу их генетического разнообразия (Global Project for the Measurement of Domestic Animal Genetic Diversity, MoDAD) (6). По микросателлитным локусам изучено более 50 популяций разных видов птицы (7-9).
Исследование микросаттелитных локусов индеек ( Meleagris gallopa-vo ) началось в 2000-х годах, при этом использовали микросателлитные панели, разработанные для кур ( Gallus gallus ) . K. Reed с соавт. (10) для работы на индейках применили 520 куриных микросателлитных маркеров. В 280 случаях (54 %) были получены продукты амплификации, большинство из которых оказались или близкими по размеру с фрагментами, амплифи-цированными с ДНК курицы, или полностью совпадали. При оценке информативности для генетического картирования индейки был определен аллельный полиморфизм 57 из 280 амплифицированных участков. В общей сложности 20 из 57 маркеров (35 %) оказались полиморфными (в среднем 1,4 аллель на локус). Сделано заключение, что для картирования генома индейки могут использоваться около 20 % микросателлитных локусов курицы (10).
Куриные микросателлиты применяли для изучения генофонда индеек пород брианзоло, колли евджений и итальянская черная (brianzolo, colli euganei, nero d’Italia). Из 31 локуса информативными оказались 22. При этом 12 локусов (ADL0112, LEI0192, LEI0234, MCW0014, MCW0016, MCW0037, MCW0067, MCW0098, MCW0103, MCW0111, MCW0165, MCW0183) изучали в одиночных ПЦР, 10 локусов — с помощью мультиплексных панелей (Multiplex Master Mix 1 — ADL0268, ADL0278, LEI0094, MCW0216, MCW0248; Master Mix 2 — MCW0034, MCW0069, MCW0081, MCW0222, MCW0295). В девяти маркерах (LEI0166, MCW0020, MCW0078, MCW0080, MCW0104, MCW0123, MCW0248, MCW0284, MCW0330) ам-плифицированных участков ДНК не обнаружили (11). Восемь микроса-теллитных маркеров кур Gallus gallus (MCW0111, MCW0067, LEI0104, MCW0123, MCW0081, MCW0069, MCW0104, MCW0183), из которых семь оказались полиморфными, применялись для изучения генофонда индеек BUT (British United Turkeys) BIG6 и BIG10 (12, 13).
В дальнейшем использовали 772 микросателлитных маркера, раз- работанных для трех видов птиц — курицы, перепелки и индейки. В результате скрининга для исследования аллельного полиморфизма и построения генетической карты индеек отобрали 410 микросателлитных локусов (53,1 %). На специально созданной генетической модели (семье), включающей прямых родственников трех поколений (224 особи), проводили генотипирование по отобранным локусам. Из 410 маркеров 109 (26,6 %) оказались полиморфными (2,3 аллеля на маркер). Более высокий полиморфизм (61,1 %) выявили при использовании специфичных для индеек маркеров. При применении маркеров, специфичных для перепелов и курицы, полиморфность составила соответственно 33,3 и 22,7 %. Авторы пришли к заключению, что перепелиные и куриные микросателлитные локусы могут быть применены для построения сравнительной генетической карты индеек (14).
Следующим этапом стала интеграция данных, полученных в двух научных центрах — Рослинском институте (the Roslin Institute, Эдинбург, Шотландия) и университете Миннесоты (the University of Minnesota, Миннеаполис, США) (15). На индейках экспериментальной фермы Миннесотского университета из 279 микросателлитных маркеров, выявленных и исследованных в Рослинском институте, скрининговым исследованиям подвергли 240. Из них 89 оказались генетически информативными и использовались для генотипирования потомства F 2 . Анализ с применением пакета программ BLAST (Basic Local Alignment Search Tool) позволил унифицировать 483 нуклеотидные последовательности микросателлитов.
Также было выполнено BLAST выравнивание маркерных последовательностей геномов индейки и курицы. Выявлено 263 совпадения и 1700 последовательностей, имеющих высокую гомологию (15).
В совместной работе ученых из США и Турции проводился поиск информативных микросателлитных маркеров для изучения биоразнообразия индеек и создания унифицированной панели. На основе нуклеотидной библиотеки данных был осуществлен дизайн праймеров для 164 фрагментов генома индеек, содержащих микросателлиты. Выявлено 154 информативных генетических маркера, однако, по мнению авторов, этого недостаточно для общей панели (16).
Интерес к изучению генетического разнообразия индеек, в том числе для генетической паспортизации коммерческих пород, прежде всего продиктован тем, что производство мяса индейки в мире постоянно растет. Оно составляет 6,1 млн т в общемировом объеме мяса птицы, занимая второе место. Положительная динамика наблюдается и в России: по данным компания Agrifood Stratedges, рост производства мяса индейки в 2019 году по сравнению с 2007 составил 185 тыс. т (с 37 до 289 тыс. т, или в 7,8 раза), что характеризует этот сегмент рынка как наиболее перспективный и быстро растущий.
Россия имеет собственные генетические ресурсы в индейководстве: в реестре селекционных достижений зарегистрировано семь пород, три кросса и семь линий индеек отечественной селекции. Северо-Кавказская зональная опытная станция по птицеводству — не только правообладатель отечественного генофонда индеек, но и единственное в России предприятие, где ведется планомерная работа по созданию новых селекционных форм.
Впервые генетический профиль российских пород индеек по мик-росателлитным локусам был изучен в 2017 году. Численность в выборках исследованных пород колебалась от 9 до 15 особей, для оценки репрезентативности выборок использовали индекс Хантера-Гастона, кластерный анализ проводили методом UPGMA, дендрограмму строили с помощью компьютерной программы START 2. Было установлено, что наибольшую генетическую близость имели породы бронзовая северокавказская, белая северокавказская и белая широкогрудая, затем следовали черная тихорецкая, серебристая северокавказская и московская белая. Наиболее удаленной оказалась узбекская палевая порода (17). Однако сравнения генофонда отечественных пород индеек с генофондом импортной селекции при этом не проводили.
В настоящей работе на основе микросателлитных маркеров впервые установлены генетические взаимоотношения между породами индеек российской селекции и генофондной популяции университета Миннесоты. Показано, что величина генетических дистанций между породами во многом определяется их происхождением, ареалом разведения, а также вкладом генофонда одних пород при создании и совершенствовании продуктивных качеств других.
Цель работы — изучить генетическое разнообразие и межпородную дифференциацию индеек российской и зарубежной селекции с использованием микросателлитных локусов.
Методика. Работа выполнялась на Северо-Кавказской зональной опытной станции по птицеводству в 2019 году. У 30 особей каждой из семи пород индеек ( Meleagris gallopavo ) отечественной селекции (белой широкогрудой, BSH; бронзовой северокавказской, BrSK; белой северокавказской, BeSK; серебристой северокавказской, SSK; московской белой, MB; черной тихорецкой, CHT; узбекской палевой, UP) отбирали образцы крови из подкрыльцовой вены.
ДНК выделяли в соответствии с протоколом к коммерческому набору АмплиПрайм ДНК-сорб-В («ИнтерЛабСервис», Россия). Количество и качество выделенной ДНК контролировали с использованием спектрофотометра NanoDrop 2000 с («Thermo Scientific», США) стандартным спектрофотометрическим методом, расчет и визуализацию результата проводили в программе NanoDrop 2000, версия 1.4.2. Раствор сравнения — ТЕ-буфер pH 7,8-8,2 (ФБУН Центральный научно-исследовательский институт эпидемиологии, Россия).
Генотипирование проводили по 12 микросателлитным локусам MNT9-MNT20 (18, 19). ПЦР осуществляли на амплификаторе Т 100 («Bio-Rad Laboratories, Inc.», США) в смеси конечного объема 20 мкл, содержащей из расчета на одну реакцию следующие реагенты: по 1 мкл прямого и обратного праймеров (ФКУЗ СтавНИПЧИ Роспотребнадзора, Россия), 2 мкл раствора дНТФ, 4 мкл РНК-элюента, 10 мкл ПЦР-смесь-2red («ИнтерЛабСервис», Россия) и 2 мкл проб ДНК. Режим амплификации был следующим: 15 мин при 95 ° С; 30 с при 95 ° С, 30 с при 58 ° С (для локусов MNT10, MNT11, MNT20 — 56 ° С), 30 с при 72 ° С (35 циклов); 5 мин при 72 ° С.
С использованием станции Experion System («Bio-Rad Laboratories, Inc.», США) и набора реагентов для визуализации фрагментов ДНК Ex-perion DNA 1K Analysis Kit («Bio-Rad Laboratories, Inc.», США) проводили капиллярный электрофорез.
Для сравнения с генотипами индеек отечественных пород использовали описанные генотипы индеек (AM) генофондной фермы университета Миннесоты (Nicholas Turkey Breeding Farms) (18, 19).
Среднее число аллелей и число эффективных аллелей на локус (Na, Ne), степень наблюдаемой и ожидаемой гетерозиготности (Н о , Н е ), индекс 654
1. Последовательности праймеров, использованных в работе для генотипирования индеек ( Meleagris gallopavo ) по микросателлитным локусам
|
Локус |
Номер в GenBank |
Последовательность |
Последовательность праймеров (5´ → 3´) |
Размер аллелей, п.н. |
|
|
прямой |
обратный |
||||
|
MNT9 |
AF482368 |
(CA) 18 |
TGGGAGTGGAAAGGTGAAAG |
TTCTCCTCAGCTCAGCAACC |
164, 168 |
|
MNT10 |
AF482369 |
(TG) 10 + (TTTTG) 5 |
TTCCCAGTGCACTACCTGAAC |
TGAACAGTGATTCCACTGAAGC |
67, 78 |
|
MNT11 |
AF482370 |
(TG) 12 |
TTTCTGACACAGGTACAAGGAAAC |
GCCCTCGAGTATTAGCCACTC |
90 |
|
MNT12 |
AF482371 |
(TG) 14imp |
AGGTGTTTTTGGGCAGTCTC |
TGCAAGCACCATCTGCTAAG |
121, 145 |
|
MNT13 |
AF482372 |
(TG) 20 |
TTAGGGGATGCTGAACTGTG |
GCGTAATTGGTGCTTTCTCC |
183, 185, 187, 235 |
|
MNT14 |
AF482373 |
(CA) 10 |
AAACAGAACAACCTCAAGGACAG |
GAATTGGGTTTGCATTTGAG |
177, 181 |
|
MNT15 |
AF482374 |
(CA) 12 |
TTGTTGCTGTTGTTTTTGTGG |
TTTCTGTGCCTAAGCTTAATGTG |
188 |
|
MNT16 |
AF482375 |
(TG) 13 +(TG) 11 +(TG) 8 +(TG) 5 |
TGTTTGCCTGCAATAAGCTG |
GCACCCTCCCACTGACTG |
219, 226, 234 |
|
MNT17 |
AF482376 |
(TA) 5 +(CA) 29 |
AGGAGCACCCAGCTCAAAG |
GAGTAATACCAAGGAAAAGTGTGC |
181 |
|
MNT18 |
AF482377 |
(TG) 13 |
GCAGGCACAGAGAGCTACG |
CCAATGTTGAAGCAGGTGAG |
158, 159, 161, 162 |
|
MNT19 |
AF482378 |
(TG) 22 |
GCAGGAGGCTCTGAGCTATG |
TTATACGGAAGGCGGTTGAG |
224, 250 |
|
MNT20 |
AF482379 |
(CA) 15 |
TAACTGTCTGCCAGGTGGTG |
GATCTCGGGTGGTGATTGC |
192, 195 |
Шеннона (I) определяли с использованием пакетов программ Microsoft Excel 2007, GenAIEx v 6.5 (20). Вычисляли средние ( M) и стандартные отклонения (±SD). Генетическую структуру популяций оценивали на основании значений F st (21) и генетических дистанций по M. Nei (22). Для построения филогенетического дерева применяли метод ближайшего соседа (Neighbor Joining Method) с использованием программного обеспечения Structure 2.3.4 (23).
Результаты. Для работы отобрали микросателлитные локусы, которые применялись при исследовании индеек генофондной фермы университета Миннесоты (NTBF) (табл. 1).
Анализ полученных данных позволил установить, что индейки российских пород и генофондной фермы университета Миннесоты характеризовались низким генетическим разнообразием. Число выявленных аллелей в локусах микросаттеллитов в целом по породной выборке варьировало от 1 до 4 (табл. 2).
2. Число аллелей в локусах микросателлитов у индеек ( Meleagris gallopavo ) отечественных пород (Северо-Кавказская зональная опытная станция по птицеводству, Ставропольский край, 2019 год) и популяции генофондной фермы университета Миннесоты (Nicholas Turkey Breeding Farms)
|
Локус |
Порода |
|||||||
|
BSH |
BrSK |
BeSK |
SSK |
MB |
CHT |
UP |
АМ |
|
|
MNT9 |
2 |
2 |
2 |
1 |
2 |
2 |
1 |
2 |
|
MNT10 |
1 |
1 |
1 |
2 |
2 |
1 |
1 |
1 |
|
MNT11 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
3 |
|
MNT12 |
2 |
2 |
2 |
1 |
2 |
1 |
1 |
1 |
|
MNT13 |
4 |
3 |
3 |
2 |
2 |
1 |
1 |
2 |
|
MNT14 |
2 |
1 |
1 |
1 |
2 |
1 |
1 |
2 |
|
MNT15 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
2 |
|
MNT16 |
2 |
2 |
2 |
1 |
1 |
2 |
1 |
2 |
|
MNT17 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
2 |
|
MNT18 |
3 |
2 |
2 |
1 |
3 |
1 |
1 |
1 |
|
MNT19 |
1 |
1 |
1 |
1 |
2 |
2 |
2 |
1 |
|
MNT20 |
2 |
1 |
1 |
1 |
2 |
2 |
2 |
2 |
Примечание. BSH — белая широкогрудая, BrSK — бронзовая северокавказская, BeSK — белая северокавказская, SSK — серебристая северокавказская, MB — московская белая, CHT — черная тихорецкая, UP — узбекская палевая породы, АМ — популяция генофондной фермы университета Миннесоты (18, 19).
Среднее число аллелей (Na) на локус колебалось от 1,0 до 1,83, при этом наибольшим числом характеризовались породы BSH (1,83), MB (1,75) и популяция АМ (1,75). По одному аллелю на локус выявили в породах SSK и UP. Аналогичные закономерности наблюдались в отношении числа эффективных аллелей (Ne): наибольшее значение этого показателя было в породе BSH и популяции АМ (1,58 и 1,55), минимальное — в породах SSK и UP (1,0), породы московская белая и черная тихорецкая занимали промежуточное положение (1,30 и 1,16). Выявленное низкое число аллелей на микросателлитный локус у индеек российской селекции согласуется с данными зарубежных исследователей, которые указывают на невысокое генетическое разнообразие коммерческих пород индеек по сравнению с другими видами сельскохозяйственных животных и птицы, а также дикими популяциями индеек (24, 25).
Сопоставление значений ожидаемой и наблюдаемой гетерозиготности (H o - H e ) показало недостаток гетерозигот во всех изученных породах и популяции от 0,2 до 0,6 %. О низком генетическом разнообразии свидетельствовал и информационный индекс Шеннона, который не превышал значения 0,50, а в породах SSK и UP был равен нулю (табл. 3).
Расчет генетический дистанций по M. Nei (табл. 4) и кластерный анализ с применением метода Neighbor Joining tree позволили получить графическое отображение филогенетического родства (рис.) между поро- дами индеек российской селекции и популяцией генофондной фермы университета Миннесоты.
-
3. Генетическое разнообразие индеек ( Meleagris gallopavo ) отечественных пород ( n = 30, Северо-Кавказская зональная опытная станция по птицеводству, Ставропольский край, 2019 год) и популяции генофондной фермы университета Миннесоты (Nicholas Turkey Breeding Farms) по локусам микросателлитов ( M ±SD)
-
4. Генетическая дифференциация индеек ( Meleagris gallopavo ) отечественных пород (Северо-Кавказская зональная опытная станция по птицеводству, Ставропольский край, 2019 год) и популяции генофондной фермы университета Миннесоты (Nicholas Turkey Breeding Farms)
|
Порода |
Na |
Ne |
Ho |
1 He 1 |
H o - H e |
] I |
|
BSH |
1,83±0,27 |
1,58±0,19 |
0,273±0,07 |
0,279±0,07 |
- 0,006 |
0,416±0,12 |
|
BrSK |
1,50±0,19 |
1,07±0,03 |
0,063±0,02 |
0,065±0,02 |
- 0,002 |
0,122±0,04 |
|
BeSK |
1,50±0,19 |
1,43±0,18 |
0,207±0,07 |
0,212±0,07 |
- 0,005 |
0,304±0,11 |
|
SSK |
1,00±0,01 |
1,00±0,00 |
0,000±0,00 |
0,000±0,00 |
0,000 |
0,000±0,00 |
|
MB |
1,75±0,17 |
1,30±0,12 |
0,189±0,05 |
0,194±0,05 |
- 0,005 |
0,312±0,08 |
|
CHT |
1,33±0,14 |
1,16±0,08 |
0,106±0,04 |
0,111±0,04 |
0,005 |
0,162±0,07 |
|
UP |
1,00±0,01 |
1,00±0,00 |
0,000±0,00 |
0,000±0,00 |
0,000 |
0,000±0,00 |
|
АМ |
1,75±0,17 |
1,55±0,15 |
0,286±0,07 |
0,291±0,07 |
0,005 |
0,419±0,14 |
Примечание. BSH — белая широкогрудая, BrSK — бронзовая северокавказская, BeSK — белая северокавказская, SSK — серебристая северокавказская, MB — московская белая, CHT — черная тихорецкая, UP — узбекская палевая породы, АМ — популяция генофондной фермы университета Миннесоты (18, 19); Na и Ne — среднее и эффективное число аллелей на локус, H o и H e — наблюдаемая и ожидаемая гетерозиготность, I — индекс Шеннона.
|
Порода |
BSH |
BrSK |
BeSK |
SSK |
MB |
CHT |
UP |
AM |
|
BSH |
0,000 |
0,250 |
0,127 |
0,594 |
0,344 |
0,457 |
0,162 |
4,872 |
|
BrSK |
0,346 |
0,000 |
0,046 |
0,425 |
0,493 |
0,430 |
0,461 |
- |
|
BeSK |
0,180 |
0,127 |
0,000 |
0,579 |
0,514 |
0,469 |
0,212 |
- |
|
SSK |
0,495 |
0,423 |
0,514 |
0,000 |
0,131 |
0,363 |
1,386 |
- |
|
MB |
0,300 |
0,467 |
0,443 |
0,178 |
0,000 |
0,228 |
0,760 |
- |
|
CHT |
0,392 |
0,431 |
0,415 |
0,502 |
0,267 |
0,000 |
0,762 |
- |
|
UP |
0,241 |
0,612 |
0,339 |
0,870 |
0,672 |
0,672 |
0,000 |
3,684 |
|
AM |
0,600 |
0,728 |
0,643 |
0,735 |
0,633 |
0,705 |
0,723 |
0,000 |
Примечание. BSH — белая широкогрудая, BrSK — бронзовая северокавказская, BeSK — белая северокавказская, SSK — серебристая северокавказская, MB — московская белая, CHT — черная тихорецкая, UP — узбекская палевая породы, АМ — популяция генофондной фермы университета Миннесоты (18, 19). Над диагональю — генетические дистанции по M. Nei (22), под диагональю — значения F st при парном сравнении. Прочерки означают отсутствие показателей.
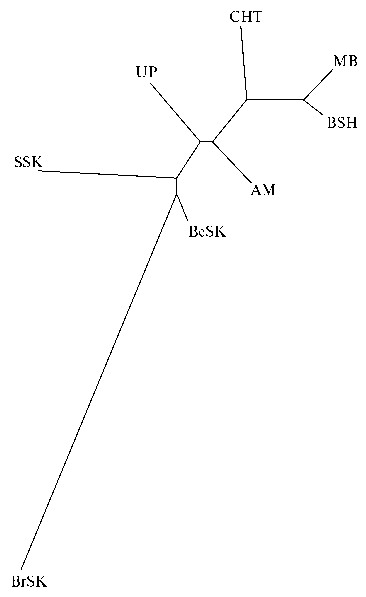
Генетические взаимосвязи индеек ( Meleagris gallopavo ) отечественных пород (Северо-Кавказская зональная опытная станция по птицеводству, Ставропольский край, 2019 год) и популяции генофондной фермы университета Миннесоты (Nicholas Turkey Breeding Farms) на основе генетических дистанций M. Nei (22): BSH — белая широкогрудая, BrSK — бронзовая северокавказская, BeSK — белая северокавказская, SSK — серебристая северокавказская, MB — московская белая, CHT — черная тихорецкая, UP — узбекская палевая породы, АМ — популяция генофондной фермы университета Миннесоты (18, 19).
Дендрограмма генетических дистанций демонстрировала наименьшее генетическое различие между породами MB и BSH. Породы BeSK, SSK и BrSK образовали отдельный узел, при этом наибольшее генетическое удаление проявляла порода BrSK, как в этом узле, так и с другими породами, образуя наибольшую по генетическому расстоянию ветвь. Отдельные ветви на относительно равном удалении образовали по- породы CHT, UP и популяция АМ. Расположение пород индеек на дереве генетических дистанций, по-видимому, было обусловлено историей их создания. Белая широкогрудая — одна из старейших в мире пород, созданных в 1960-х годах в США. Она ведет генетическое происхождение от голландских белых индеек. На Северо-Кавказскую зональную станцию по птицеводству белая широкогрудая порода индеек, а именно четыре линии кросса Hidon (А, В, С, Д), были завезены в 1980 году из Голландии. На базе линий В и D были отселекционированы родительские формы, сохранившиеся в чистоте до настоящего времени. Образцы их ДНК были использованы в настоящем исследовании. Московская белая порода создавалась в Московской области. На первоначальном этапе были использованы местные белые индейки и белтсвиллские индейки, на заключительном — белые голландские индейки. То есть генофонд белых индеек длительно использовался при создании пород белой широкогрудой и белой московской, что и обусловило их близкое расположение на дереве генетических расстояний.
Первую отечественную породу индеек — бронзовую северокавказскую вывели в 1950-1960-х годах скрещиванием местных аборигенных индеек с производителями бронзовой и бронзовой широкогрудой пород. Вторая отечественная порода — белая северокавказская была создана в 1970-1980-е годах посредством скрещивания аборигенных бронзовых северокавказских индеек с самцами белой широкогрудой породы английского происхождения. При выведении серебристой северокавказской породы, которая была официально признана в 2008 году, популяция палевых узбекских индеек на первых этапах улучшалась белой широкогрудой породой, преимущественно самцами линии О4 материнского типа, с целью повышения воспроизводства и мясной продуктивности. На заключительном этапе создания породы особи с разной долей генофонда белой широкогрудой разводились «в себе» при жесткой выбраковке не соответствующих требованиям фенотипов. Следовательно, белая северокавказская и серебристая северокавказская породы создавались при использовании генофонда белой широкогрудой, что, по-видимому, определило их большую генетическую близость между собой и некоторую удаленность от бронзовой северокавказской, которая создавалась исключительно с использованием пород бронзового оперения. Возможно, на образование общего узла бронзовой, белой и серебристой северокавказской породами повлияло и то, что все они создавались в Северо-Кавказском регионе. Однотипные факторы среды обитания, по-видимому, способствовали отбору близких генотипов.
Генетическая удаленность пород черная тихорецкая и узбекская палевая объясняется тем, что они создавались в большей степени при использовании популяций местных индеек. Узбекская палевая порода выводилась на основе узбекских бронзовых индеек, которые точечно улучшались белой широкогрудой породой. Использование белой широкогрудой породы при выведении узбекской палевой должно было повлиять на их генетическую близость. Однако черная тихорецкая порода оказалась ближе к породам белая широкогрудая и московская белая. Можно предположить, что значительная географическая удаленность ареала создания узбекской палевой породы обусловила большее генетическое различие с московской и белой широкогрудой породами, которые, как и черная тихорецкая, создавались в Северо-Кавказском регионе.
Определенную генетическую удаленность от российских пород проявляла популяция индеек генофондной фермы университета Миннесоты. Однако эта удаленность оказалась не столь выраженной, как ожидалось. Вероятно, генофонд популяции АМ включает как генофонд пород с бронзовым оперением, так и генофонд наиболее распространенной во всем мире белой широкогрудой породы, что определяет ее равную удаленность от изученных отечественных пород индеек.
Полученные в настоящем исследовании данные во многом совпадают с результатами В.И. Фисинина с соавт. (17). Построение дендрограммы на основе генетических дистанций c использованием тех же мик-росателлитных локусов, но на меньшей по объему выборке и с применением программы START 2 распределило изученные породы в два кластера. Первый кластер был образован частью генотипов белой широкогрудой и всеми генотипами породы узбекская палевая, второй — двумя крупными подкластерами. Первый подкластер сформировали генотипы пород бронзовая северокавказская, белая северокавказская и белая широкогрудая, второй — генотипами пород черная тихорецкая, серебристая северокавказская и московская белая (17). В указанном выше и в настоящем исследовании наиболее близкими оказались породы белая широкогрудая и московская белая, белая северокавказская и серебристая северокавказская, большее генетическое удаление проявляли черная тихорецкая и узбекская палевая. Использование в представляемой работе большего числа особей и метода ближайшего соседа в программе Structure 2.3.4 позволили выявить более значительную генетическую дифференциацию бронзовой северокавказской породы.
В другом исследовании, также выполненном на Северо-Кавказской зональной опытной станции по птицеводству, с помощью метода ДНК-фингерпринтинга было показано, что наиболее близки породы бронзовая северокавказская и белая северокавказская, затем следовали серебристая северокавказская и узбекская палевая. Значительную генетическую удаленность от пород белой широкогрудой, белой северокавказской, серебристой северокавказской, бронзовой северокавказской и узбекской палевой проявляла черная тихорецкая (26).
Многие авторы указывают, что основными факторами, влияющими на степень генетической дифференциации домашних пород и диких популяций индеек, становятся использование генофонда одних пород при создании других и географическая местность их разведения. При этом ученые единодушны во мнении, что геном индеек значительно менее разнообразен по сравнению с геномом других видов сельскохозяйственных животных и птицы.
E.K. Latch с соавт. (24) исследовали дикую восточную ( M. gallopavo silvestris ) и домашнюю индейку ( M. gallopavo ), используя семь микросател-литных маркеров. Число аллелей на локус варьировало от 5 до 15, при этом домашние индейки по сравнению с восточными дикими характеризовались достоверно меньшими числом аллелей на локус и общей гетерозиготностью.
D. Kamara с соавт. (27) изучали генетическую дифференциацию между коммерческими и некоммерческими породами индеек — нарраган-сетской, бурбонской красной, синевато-серой, испанской черной и королевской пальмовой (Narra-gansett, Bourbon Red, Blue Slate, Spanish Black и Royal Palm) из генофондной коллекции фермы колледжа Вирджиния по 10 микросателлитным локусам (RHT0009, RHT0011, RHT0024, RHT0095, RHT0131, RHT0216, RHT0294, TUM16, TUM20, ADL0023). С помощью филогенетического анализа установлено, что наррагансетская, бурбонская красная и синевато-серая породы имели большее генетическое сходство с коммерческими породами, чем испанская черная и королевская пальмовая (27). Аналогичные данные для этих пород были получены с другими гене- тическими маркерными системами (SNP и фрагменты ДНК случайных последовательностей — random amplification of polymorphic DNA, RAPD) (28). S. Kusza с соавт. (25) провели четкую генетическую дифференциацию между венгерской бронзовой и белой широкогрудой породами индеек на основе 15 микросателлитных локусов. Венгерская бронзовая порода характеризовалась большей полиморфностью (среднее число аллелей на локус 3,20), чем белая широкогрудая (среднее число аллелей на локус 2,77).
K.E. Mock с соавт. (29) использовали микросателлитные маркеры и митохондриальную ДНК в наиболее изменчивой ее части для исследования генетических взаимоотношений между дикими популяциями индеек. Они изучили 24 популяции шести подвидов диких индеек: семь — Рио Гранде (Rio Grande, M. gallopavo intermedia ), шесть — восточной индейки ( M. gallopavo silvestris ), три — Флорида (Florida, M. gallopavo osceola ), пять — Мерриам (Merriam’s, M. gallopavo merriami ), три — Гулда (Gould’s, M. gallopavo mexicana ). Авторы установили соответствие современного разделения подвидов дикой индейки, основанного на морфологическом описании, их генетическим характеристикам, за исключением восточной индейки и Флорида, которые не обнаруживали генетических различий. В популяциях Мерриам и Рио-Гранде выявлена положительная связь между генетическим и географическим расстоянием, тогда как в популяциях восточной индейки такой зависимости не обнаружено.
Для получения более полных сведений о генетическом разнообразии пород и линий индеек M.L. Aslam с соавт. (30) использовали более современный и информативный метод — полногеномное секвенирование. В результате сканирования генома 32 индеек из разных популяций было выявлено 5,49 млн SNP по отношению к описанному референсному геному индейки (UMD 2.01), который составляет 1,1 млрд оснований (31, 32). Гетерозиготность особей варьировала от 0,17 до 2,73 SNP, а по всей выборке колебалась от 0,73 до 1,64 SNP на тысячу пар нуклеотидов. Авторы пришли к выводу, что исследованные коммерческие породы и линии индеек имеют общее происхождение, при этом генетической основой при их выведении были дикие формы индеек, которые характеризуются более высокой гетерозиготностью. Также авторы подчеркивают, что геном индеек в сравнении с геномом других видов сельскохозяйственных животных и птицы характеризуется значительно большей консервативностью.
Таким образом, показано генетическое своеобразие и отличия генотипов пород индеек селекционно-генетического центра по разведению и сохранению генофонда отечественных пород индеек в сравнении с популяцией индеек генофондной фермы университета Миннесоты. Наименьшие генетические различия установлены между породами московская белая и белая широкогрудая, белая северокавказская и серебристая северокавказская. Породы черная тихорецкая и узбекская палевая, а также популяция индеек генофондной фермы университета Миннесоты проявляли большую генетическую удаленность как от указанных выше пород, так и между собой. Наибольшую генетическую дифференциацию демонстрировала порода бронзовая северокавказская. Внутрипородные особенности и межпородная дифференциация семи российских пород индеек по микросател-литным маркерам ДНК в значительной степени отражают историю их создания и совершенствования. Анализ числа аллелей на локус позволил подтвердить выявленную другими исследователями закономерность, согласно которой генофонд пород и популяций домашних индеек характеризуется незначительным генетическим разнообразием. Для дальнейшего рационального использования генофонда отечественных пород индеек, а также получения новой информации об их генетических особенностях и месте в генетической дифференциации среди других пород индеек, разводимых в мире, целесообразно использовать дополнительные методы и современные маркеры генетического анализа, такие как секвенирование SNP и MLST.
