Глобальная палеогеография и палеобиогеография тоарского века (ранняя юра) по брахиоподам
 по брахиоподам")
Автор: Гриненко В.С., Баранов В.В.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 5 (293), 2019 года.
Бесплатный доступ
Настоящая статья посвящена актуальным проблемам - глобальной палеогеографии и палеобиогеографии в тоарском веке юрского периода. На основании пространственно-временного анализа тоарских брахиопод и материалов глубоководного бурения предложена новая модель. Установлено, что в тоарском веке палеогеография Земли была представлена тремя суперконтинентами - Афалией, Пацифидой и Аразией - и двумя континентами - Гипербореей и Антарктидой, которые были разделены мелководными шельфовыми морскими бассейнами шириной около 2000-3000 км, сообщающимися между собой. Выполненные палеореконструкции показывают, что Бореальный бассейн соединялся с Тетическим бассейном через западный и восточный проливы. На основании анализа климатической зональности и пространственно-временного распространения тоарских брахиопод выделены три палеозоохории высшего ранга: Экваториальная, Бореальная и Нотальная надобласти (superrealms). В Экваториальной надобласти установлены Альпийско-Меланезийско-Китайская и Западно-Южно-Американская биогеографические области (realms), первая подразделяется на Альпийско-Кавказскую, Китайско-Японскую и Меланезийскую провинции (provinces). Альпийско-Кавказская провинция представлена тремя субпровинциями (subprovinces): Европейской, Северо-Африканской и Крымско-Кавказско-Аравийской. Бореальная надобласть представлена Сибирско-Аляскинской областью и Западно-Канадской провинцией Западно-Северо-Американской области. Невадская провинция Западно-Северо-Американской области входит в Экваториальную надобласть. Новозеландско-Новокаледонская область включена в Нотальную надобласть.
Палеогеография, палеобиогеография, пацифида, аразия, афалия, гиперборея, антарктида, ранняя юра, тоар, брахиоподы
Короткий адрес: https://sciup.org/149129376
IDR: 149129376 | УДК: 551.8+56/551:761.3(564.8) | DOI: 10.19110/2221-1381-2019-5-20-26
Global paleogeography and paleobiogeography of the Toarcian (Early Jurassic) on the brachiopods
This article is devoted to the actual problems of the global paleogeography and paleobiogeography in the Toarcian stage of the Jurassic period. The authors proposed a new model on the basis of spatio-temporal analysis of Toarcian brachiopods and materials of deep-sea drilling of the seabed of the World Ocean. It was established that in the Toarcian stage the paleogeography of the Earth was represented by three supercontinents Afalia, Pacifida and Arasa and two continents Hyperborea and Antarctida, which were separated by shallow shelf sea basins, about 2000-3000 km wide. Completed paleoreconstructions showed the Boreal basin was connected to the Tethys West and East straits. Based on the analysis of climatic zonality and the spatial-temporal distribution of the Toarcian brachiopods three paleozoochories of higher rank Equatorial, Boreal, and Notal superrealms were distinguished. The Equatorial Superrealm was represented by the Alpine-Melanesian-Chinese and West-South American biogeographical realms, the first was divided into the Alpine-Caucasian, Sino-Japanese and Melanesian provinces. The Alpine-Caucasian province were represented by three subprovinces: European, North-African and Crimean-Caucasian-Arabian. Boreal superrealms is represented by the Siberian-Alaska realm and the Western Canadian province of the West-North American realms. The Nevadan province of this realm is part of the Equatorial Superealm. The New Zealand-New Caledonian realm in the Notal superrealm is included.
Текст научной статьи Глобальная палеогеография и палеобиогеография тоарского века (ранняя юра) по брахиоподам
Данная работа продолжает серию публикаций по палеогеографии и палеобиогеографии среднего палеозоя — раннего мезозоя [2, 3, 7—9, 18] с использованием материалов глубоководного бурения Deep Sea Drilling Project (DSDP), 1963—1984 гг., Ocean Drilling Project (ODP), 1985—2003 гг. [4—6, 13—16], анализа геохронологического и географического распространения тоарских брахиопод [26, 27].
Зарубежными авторами рассматривались только частные вопросы палеобиогеографии тоарских брахиопод отдельных регионов. М. Манценидо [22] на па-линспатической основе рассматривал палеобиогеографию тоарских брахиопод запада Южной Америки (Андо-Патагонская область) и обосновывал ее связь c западным Тетисом наличием так называемого испанского коридора, Я. Алмирос и М. Бикауд [17] на основании распространения тоарских брахиопод приводи- 20
ли характеристику транзитных зон между биогеографическими провинциями западного Тетиса. В. А. Захаров и др. [10] представили биогеографическое районирование бореальных бассейнов по аммонитам.
В новых тектонических концепциях — тектоники литосферных плит и стадийного развития земной коры — почти не рассматриваются проблемы тектонической эволюции внутренних частей континентов. Следует отметить, что палеогеография морских бассейнов неразрывно связана с тектоническими процессами. Между тем без выяснения основных закономерностей внутриконтинентальной (бассейновой) тектоники вряд ли возможен дальнейший прогресс в развитии тектонической науки и создание общей теории глобального тектогенеза.
На предлагаемых в последнее время палеотекто-нических реконструкциях разных частей Земли в мезозое [24, 25] демонстрируется динамика литосфер- ных плит и океанических бассейнов. Авторы статьи при разработке палеогеографии и палеобиогеографии Земли в тоарском веке используют модель, которую они разработали ранее для среднего палеозоя и раннего мезозоя [3, 8]. Для трех суперконтинентов мы предложили названия Афалия, Аразия и Пацифида, а для двух континентов — Гиперборея и Антарктида. Для обоснования своих реконструкций мы использовали представителей органического мира, шельфовые ассоциации которых трассировались вдоль береговой линии морских бассейнов. Также мы использовали данные по строению и возрасту базальтового и надбазальтового слоев, которые были получены при изучении керна глубоководных скважин [4—6,13—16]. В данной статье тоарский век принимается в объеме и временных рамках согласно Ф. Градстейну и др. [19] с дальнейшими уточнениями Дж. Г. Огга и др. [23].
Основные принципы палеобиогеографического районирования
Брахиоподы всегда были удобной группой фауны для палеобиогеографических реконструкций в палеозое и мезозое, поскольку они доминировали в шельфовых биоценозах и характеризовались ареалами, совпадающими с поясами климатической зональности. Причем расселение брахиопод происходило на личиночной стадии вдоль береговых линий мелководных шельфов господствующими течениями. На пересечение глубоководных океанических бассейнов, которые присутствуют на современных палинспатических реконструкциях, у них не хватило бы времени, поскольку они могли находиться в планктонном состоянии от нескольких суток до двух-трех недель, а затем им были необходимы подходящий субстрат и соответствующая глубина, в противном случае они погибали.
Г. Вестерман [28] предложил следующую иерархию палеозоохорий: надобласть (Superrealm), область (Realm), подобласть (Subrealm), провинция (Province), субпровинция (Subprovince) и регион (Region). Р. Е. Алексеева [1] при палеозоогеографическом районировании по брахиоподам использовала систематические категории, которые применяются при районировании современных морей и океанов, и выделяла надобласть, область и провинцию. Другие [3, 18] при биогеографическом районировании морских бассейнов использовали следующие категории: надобласть, область, провинцию, субпровинцию и район. Надобласть отвечает климатическому поясу Земли и характеризуется специфическим составом подсемейств, семейств и более крупных таксономических категорий. Область входит в состав надобласти и характеризуется однородностью таксономического состава фауны на уровне родов, подсемейств и семейств, ее границы определяются по резкому изменению таксономического состава. По данным Е. Б. Неймарк и А. В. Маркова [11], таксономическое разнообразие характеризуется широтной зональностью и постепенным убыванием от тропической зоны к полюсам. Провинции выделяются в составе областей, для них характерна частичная или полная изоляция акваторий со специфичным родовым и видовым составом. Район ограничен, как правило, локальным рас пространением представителей одного или двух родов и располагается на периферии провинции или области. Для выявления степени биогеографических связей между двумя палеозоохориями использовался индекс общности Чекановского — Сёренсена:
К= 2а K (a+b)+(a+c) ’ где a — общее число родов в обеих палеобиохориях; b — число родов в первой и с — число родов во второй палеобиохории без учета общих родов. Чем выше число К, тем более тесные биогеографические связи между ними. Полная общность палеобиохорий: при K = 100 % (К = 1), при K = 0.1—0.2 наблюдаются слабые связи между палеобиохориями, при K = 0.3— 0.4 — стабильные связи и при значениях K > 0.5 — сильные связи.
Палеогеография и палеобиогеография
Согласно авторским представлениям, в тоарском веке наша планета была представлена тремя суперконтинентами: Пацифидой, Афалией и Аразией — и двумя континентами — Гипербореей и Антарктидой, которые были разделены мелководными шельфовыми морскими бассейнами шириной около 2000—3000 км, сообщающимися между собой и заложенными на континентальной коре (см. рисунок). Бореальный бассейн соединялся с Тетическим через западный и восточный проливы. Связь между Тетическим и Западно-ЮжноАмериканским бассейнами осуществлялась через морские бассейны, расположенные на территории Индонезии, Меланезии, Новой Каледонии и Новой Зеландии. В это время океанический палеобассейн Палеопацифика (= Панталасса) отсутствовал. И. А. Резанов [12] считает, что до начала океанизации (поздний триас — ранний мел) на месте Палеопацифики находилось жесткое основание платформы, при этом эволюция морских мелководных бассейнов в современные океаны всегда происходила на континентальной коре.
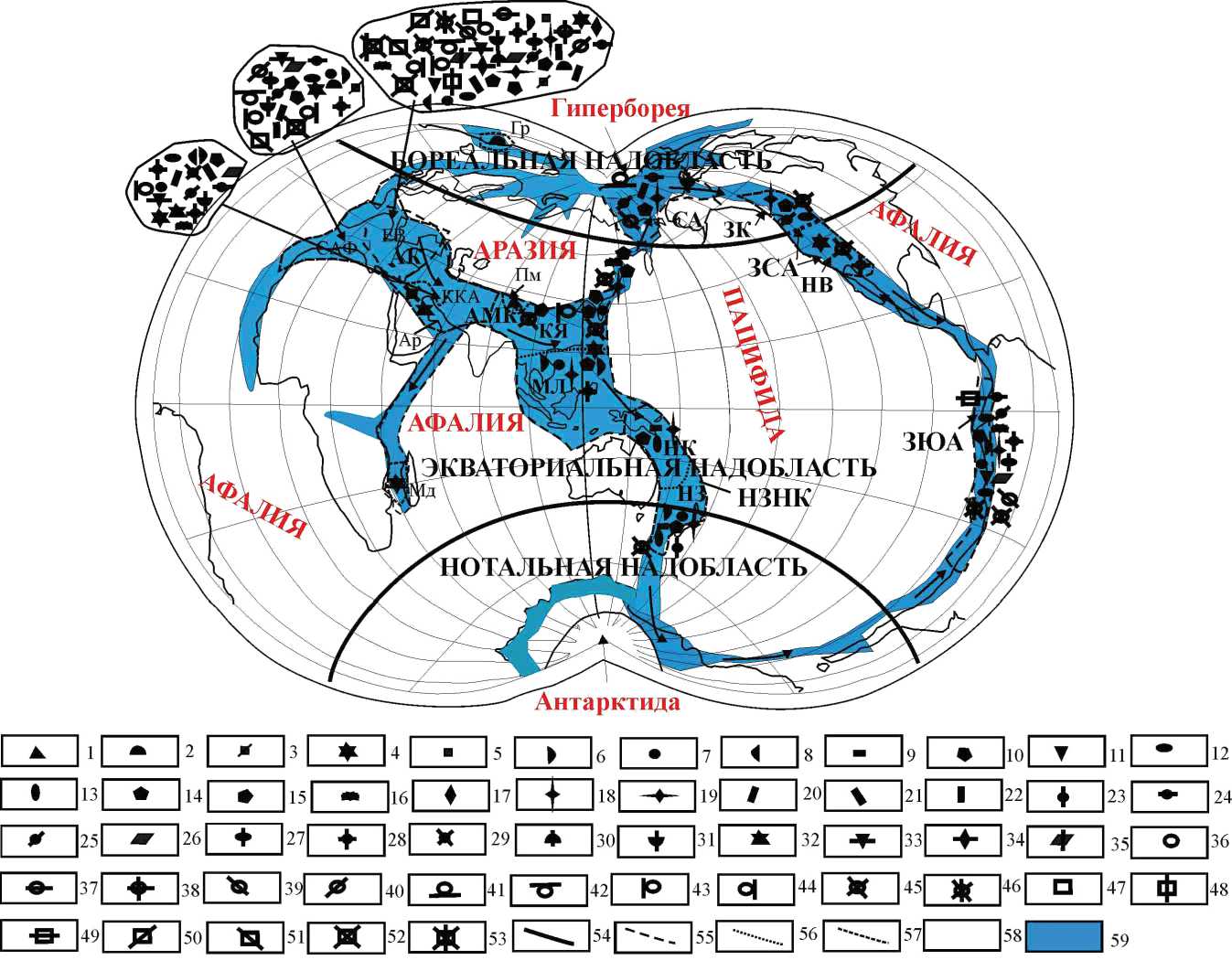
Схема глобальной палеогеографии и палеобиогеографии тоарского века (ранняя юра) по брахиоподам [8]. Области: АМК — Альпийско-Меланезийско-Китайская, ЗСА — Западно-Северо-Американская, ЗЮА — Западно-Южно-Американская, СА — Сибирско-Аляскинская, НЗНК — Новозеландско-Новокаледонская; провинции: АК — Альпийско-Кавказская, ЗК — Западно-Канадская, КЯ — Китайско-Японская, НВ — Невадская, НЗ — Новозеландская, НК — Новокаледонская; субпровинции: ЕВ — Европейская, ККА — Крымско-Кавказско-Аравийская, САФ — Северо-Африканская; районы: Ар — Аравийский, Гр — Гренландский, Мд — Мадагаскарский, Пм — Памирский. Обозначения: 1 — Almorhynchia , 2 — Apringia , 3 — Soaresirhynchia (?), 4 — Pseudogibbirhynchia , 5 — Septocrurella , 6 — Prionorhynchia , 7 — Cirpa , 8 — Bodrakella (?), 9 — Herangirhynchia (?), 10 — R. (Rhynchonelloidea) , 11 — R. (Aalenirhynchia) , 12 — Piarorhynchia , 13 — Caledorhynchia , 14 — Sakawairhynchia , 15 — Ptyctorhynchia , 16 — Rhynchonelloidella , 17 — Cubanirhynchia , 18 — Furcirhynchia , 19 — Praemonticlarella , 20 — Holcorhynchia , 21 — Maxillirhynchia , 22 — Nannirhynchia , 23 — Aucklandirhynchia , 24 — Rudirhynchia (?), 25 — T. (Tetrarhynchia) , 26 — T. (Rostrirhynchia) , 27 — Quadratirhynchia , 28 — Gibbirhynchia , 29 — Conarosia , 30 — Suessia , 31 — Spiriferina , 32 — Calyptoria , 33 — Liospiriferina , 34 — Sinucosta , 35 — Davidsonella , 36 — Eudesella , 37 — Thecidella , 38 — Slavinithyris , 39 — Loboidothyris , 40 — Merophricus , 41 — Omolonothyris , 42 — Hesperithyris , 43 — Monsardithyris , 44 — Stroudithyris , 45 — Lobothyris , 46 — Cuersithyris , 47 — Exceptothyris , 48 — Inaequalis , 49 — Notosia , 50 — Ferrythyris , 51 — Prototegulithyris , 52 — Callospiriferina , 53 — Moorellina ; 54—57 — границы: 54 — надобластей, 55 — областей, 56 — провинций, 57 — субпровинций, 58 — суша, 59 — вода.
Sheme of the global paleogeography and paleobiogeography of the Toarcian Stage (Early Jurassic) on brachiopods [8]: realms: АМС — Alpine-Melanesian-Chinese, WNА — West-North-American, WSА — West-South-American, SА — Siberian-Alaskan, NZNC — New Zealand-New Caledonian; provinces: АС — Alpine-Caucasian, WC — West-Canadian, CJ — Sino-Japanese, NV — Nevadan, NZ — New Zealandian, NC — New Caledonian; subprovinces: ER — European, CCА — Crimean-Caucasian-Arabian, NАF — North African; regions: АЬ — Arabian, Gr — Greenlandic, Мd — Madagascarian, Рм — Pamirian. Legend: 1 — Almorhynchia , 2 — Apringia , 3 — Soaresirhynchia (?) , 4 — Pseudogibbirhynchia , 5 — Septocrurella , 6 — Prionorhynchia , 7 — Cirpa , 8 — Bodrakella (?), 9 — Herangirhynchia (?), 10 — R. (Rhynchonelloidea) , 11 — R. (Aalenirhynchia) , 12 — Piarorhynchia , 13 — Caledorhynchia , 14 — Sakawairhynchia , 15 — Ptyctorhynchia , 16 — Rhynchonelloidella , 17 — Cubanirhynchia , 18 — Furcirhynchia , 19 — Praemonticlarella , 20 — Holcorhynchia , 21 — Maxillirhynchia , 22 — Nannirhynchia , 23 — Aucklandirhynchia , 24 — Rudirhynchia (?), 25 — T. (Tetrarhynchia) , 26 — T. (Rostrirhynchia) , 27 — Quadratirhynchia , 28 — Gibbirhynchia , 29 — Conarosia , 30 — Suessia , 31 — Spiriferina , 32 — Calyptoria , 33 — Liospiriferina , 34 — Sinucosta , 35 — Davidsonella , 36 — Eudesella , 37 — Thecidella , 38 — Slavinithyris , 39 — Loboidothyris , 40 — Merophricus , 41 — Omolonothyris , 42 — Hesperithyris , 43 — Monsardithyri s, 44 — Stroudithyris , 45 — Lobothyris , 46 — Cuersithyris , 47 — Exceptothyris , 48 — Inaequalis , 49 — Notosia , 50 — Ferrythyris , 51 — Prototegulithyris , 52 — Callospiriferina , 53 — Moorellina ; 54—57 — borders: 54 — superrealms, 55 — realms, 56 — provinces, 57 — subprovinces, 58 — land, 59 — water.
ском веке, а представители отряда Koninckinidina были распространены только в пределах данной области.
Европейская субпровинция характеризуется представителями 44 родов: Apringia , Soaresirhynchia(?) , Pseudogibbirhynchia , Septocrurella , Prionorhynchia , Cirpa, Bodrakella (?), R. (Rhynchonelloidea) , R. (Aalenirhynchia),
Piarorhynchia , Cubanirhynchia, Maxillirhynchia,
Ptyctorhynchia, Furcirhynchia, Holcorhynchia,
Rhynchonelloidella, Praemonticlarella, Rudirhynchia(?),
T. (Tetrarhynchia) , T. (Rostrirhynchia) , Quadratirhynchia , Gibbirhynchia , Conorosia , Suessia , Spiriferina , Calyptoria , Liospiriferina , Sinucosta , Davidsonella , Eudesella , Thecidella , Slavinithyris , Loboidothyris , Merophricus , Hesperithyris , Monsardithyris , Stroudithyris , Lobothyris , Cuersithyris , Exceptothyris , Inaequalis , Ferrythyris , Prototegulithyris , Callospiriferina .
Общими для Европейской и Северо-Африканской субпровинций являются Apringia, Soaresirhynchia(?), Pseudogibbirhynchia, Prionorhynchia, Cirpa (Cirpinae), R. (Rhynchonelloidea), Piarorhynchia, Ptyctorhynchia, Holcorhynchia, Nannirhynchia, Ferrythyris, Rudirhynchia(?), T. (Tetrarhynchia), Gibbirhynchia, Spiriferina, Quadra-tirhynchia, Liospiriferina, Callospiriferina, Merophricus, Hesperithyris, Monsardithyris, Stroudithyris, Protote-gulithyris, из них 22 общих рода и подрода, 17 семейств и подсемейств. Европейская субпровинция отличается от Северо-Африканской присутствием представителей 23 родов и подродов: Bodrakella(?), Septocrurella, Rhynchonelloidea (Aalenirhynchia), Sakawairhynchia, Rhynchonelloidella, Cubanirhynchia, Furcirhynchia, Praemonticlarella, Maxillirhynchia, Suessia, Spiriferina, Sinucosta, Moorellina, Davidsonella, Eudesella, Thecidella, Slavinithyris, Loboidothyris, Lobothyris, Cuersithyris, Exceptothyris, Inaequalis, Ferrythyris. На африканском шельфе отсутствуют представители родов бра-хиопод Septocrurella, Bodrakella(?), R. (Aalenirhynchia), Sakawairhynchia, Rhynchonelloidella, Cubanirhynchia, Furcirhynchia, Praemonticlarella, Maxillirhynchia, Suessia, Sinucosta, Davidsonella, Eudesella, Thecidella, Slavinithyris, Lobothyris, Cuersithyris, Exceptothyris, Inaequalis, Notosia, Ferrythyris, распространенных на северном побережье западного Тетиса. Эндемиков здесь не обнаружено, что свидетельствует о тесных биогеографических связях внутри западного Тетиса. Крымско-Кавказско-Аравийская субпровинция характеризуется представителями 19 родов и подродов: Pseudogibbirhynchia, Prionorhynchia, Cirpa, Bodrakella(?), R. (Rhynchonelloidea, Piarorhynchia, Rhynchonelloidella, Cubanirhynchia Holcorhynchia, Maxillirhynchia, Rudirhynchia(?), T. (Tetrarhynchia), Gibbirhynchia, Conarosia, Spiriferina, Calyptoria, Liospiriferina, Monsardithyris. Она отличается от Европейской субпровинции отсутствием представителей 29 родов и подродов: Apringia, Soaresirhynchia(?), Rhynchonelloidea (Aalenirhynchia), Sakawairhynchia, Ptyctorhynchia, Furcirhynchia, Praemon ticlarella, Nannirhynchia, Spiriferina, Quadratirhynchia, Tetrarhynchia (Rostrirhynchia), Suessia, Sinucosta, Moorellina, Eudesella, Thecidella, Slavinithyris, Loboidothyris, Merophricus, Hesperithyris, Stroudithyris, Lobothyris, Cuersithyris, Exceptothyris, Inaequalis, Ferrythyris, Prototegulithyris, Callospiriferina. Крымско-Кавказско-Аравийская субпровинция, так же как и Северо-Африканская, отличается от Европейской субпровинции полным отсутствием представителей отряда Thecideida и подотря да Koninckinidina. Крымско-Кавказско-Аравийская субпровинция имеет 12 общих представителей родов и подродов брахиопод с Северо-Африканской субпровинцией: Pseudogibbirhynchia, Prionorhynchia, Cirpa, R. (Rhynchonelloidea), Piarorhynchia, Holcorhynchia(?), Rudirhynchia, T. (Tetrarhynchia), T. (Rostrirhynchia), Gibbirhynchia, Spiriferina, Liospiriferina и отличается от нее отсутствием представителей 5 родов бра-хиопод: Apringia, Soaresirhynchia(?), Ptyctorhynchia, Nannirhynchia, Quadratirhynchia. В пределах субпровинции можно выделить Аравийский биогеографический район, характеризующийся двумя эндемичными родами брахиопод — Conarosia и Calyptoria. Индекс общности Чекановского — Сёренсена Европейской и Северо-Африканской субпровинций составляет 0.41, Европейской и Крымско-Кавказско-Аравийской — 0.35 и Северо-Африканской и Крымско-Кавказско-Аравийской — 0.27, что свидетельствует о стабильных биогеографических связях между ними.
Китайско-Японская провинция охватывает территорию от Памира и Тибета на западе до Японии и Приморья на востоке. В тоарском веке в состав этой провинции входили представители 8 родов брахиопод: Almorhynchia , Cirpa , Sakawairhynchia , Ptyctorhynchia , Rhynchonelloidella , Furcirhynchia , Monsardithyris , Lobothyris , из них 6 родов: Ptyctorhynchia , Cirpa , Rhynchonelloidella , Furcirhynchia , Monsardithyris , Lobothyris — известны в Европейской провинции. Единственный эндемичный род Almorhynchia , найденный на Памире, ограничен территорией Памирского биогеографического района. Индекс общности Чекановского — Сёренсена Европейской и Китайско-Японской провинций составляет 0.19, что свидетельствует о слабых биогеографических связях между ними.
Меланезийская провинция охватывает территорию Лаоса, Вьетнама, Индонезии и Меланезии. Она, так же как и Китайско-Японская, характеризуется бедным родовым составом брахиопод. Здесь присутствуют представители 7 родов: Pseudogibbirhynchia , Prionorhynchia , Cirpa , Sakawairhynchia , Furcirhynchia , Gibbirhynchia и Lobothyris , которые известны в западных провинциях Альпийско-Меланозийско-Китайской области. Индекс общности Чекановского — Сёренсена Меланезийской и Китайско-Японской провинций составляет 0.35, что свидетельствует о стабильных биогеографических связях между ними.
Западно-Северо-Американская область занимала запад Северной Америки, от Западной Канады на севере до Невады на юге. Она характеризуется присутствием представителей двух отрядов: Rhynchonellida и Terebratulida, надсемейства Rhynchonelloidea, трех семейств: Rhynchonellidae, Tetrarhynchiidae и Lobothyrididae, четырех подсемейств: Rhynchonellinae, Piarorhynchiinae, Davanirhynchiinae и Lobothyridinae, семи родов и подродов брахиопод: Pseudogibbirhynchia , Piarorhynchia , Sakawairhynchia , Furcirhynchia , T . ( Tetrarhynchia ), Gibbirhynchia и Lobothyris . Здесь пока не обнаружены представители отряда Spiriferinida, присутствующие в соседней Сибирско-Аляскинской области. Общими с ней являются представители родов Sakawairhynchia , Furcirhynchia и подрода T . ( Tetrarhynchia ). Индекс общности Чекановского — Сёренсена составляет 0.30, что свидетельствует о стабильных биогеографических связях между ними. Западно-Северо-Американская область разделяется на Западно-Канадскую и Невадскую провинции. Первая представлена Piarorhynchia, Sakawatrhynchia, Gibbirhynchia , Furcirhynchia и Lobothyri s и расположена в Бореальной надобласти, а вторая находится в зо- 24
не экотона между Бореальной и Экваториальной надобластями и характеризуется представителями родов Pseudogibbirhynchia, Gibbirhynchia и Lobothyri s, из которых последний является общим для обеих провинций, что свидетельствует о наличии связи между ними.
Новозеландско-Новокаледонская область расположена вдоль юго-западного обрамления суперконтинента Пацифида. На севере она граничит с Альпийско-Меланезийско-Китайской, а вдоль южного обрамления — с Западно-Южно-Американской областями. Здесь обнаружены представители двух отрядов: Rhynchonellida и Terebratulida, надсемейства Rhynchonelloidea, четырех семейств: Wellerellidae, Rhynchonellidae, Ochotorhynchiidae, и Lobothyrididae, пяти подсемейств: Cirpinae, Rhynchonellinae, Davanirhynchiinae, Cyclothyridinae и Lobothyridinae, семи родов брахиопод: Prionorhynchia , Cirpa , Herangirhynchia (?) , Caledorhynchia Sakawairhynchia , Furcirhynchia и Lobothyris . Отличается от Западно-Южно-Американской области отсутствием представителей надсемейства Thecideoidea. Характеризуется присутствием представителей пяти космополитных и двух эндемичных родов брахиопод — Herangirhynchia и Caledorhynchia . С Западно-Южно-Американской областью ее связывает четыре общих рода: Prionorhynchia , Cirpa , Furcirhynchia и Lobothyris . Индекс общности Чекановского — Сёренсена между ними составляет 0.28, что свидетельствует о существовании стабильных биогеографических связей. Между Новозеландско-Новокаледонской областью и Меланезийской провинцией Альпийско-Меланезийско-Китайской области индекс общности Чекановского — Сёренсена составляет 0.42, что свидетельствует о стабильной биогеографической связи между ними. Внутри Новозеландско-Новокаледонской области выделяются две провинции: Новозеландская и Новокаледонская, первая представлена Priono-rhynchia , Cirpa , Herangirhynchia(?) , Caledorhynchia, Sakawairhynchia , Furcirhynchia и Lobothyris , а вторая — Herangirhynchia(?) , Caledorhynchia, Sakawairhynchia и Furcirhynchia . Новокаледонская провинция отличается от Новозеландской отсутствием представителей трех родов Prionorhynchia , Cirpa и Lobothyris . Индекс общности Чекановского — Сёренсена между ними составляет 0.44, что свидетельствует о тесной биогеографической связи.
Заключение
Основным центром родообразования брахиопод в тоаре, так же как и в геттанге — плинсбахе, оставалась Европейская субпровинция, а нивелировка дифференциации брахиоподовой фауны во всех палеобассейнах Земли увязывается с эвстатическим максимумом в тоарском веке [20, 21]. Эндемичные таксоны в этот отрезок времени составляют лишь незначительный процент от общего родового состава брахиопод: Сибирско-Аляскинская — 12.5 %, Новозеландско-Новокаледонская — 28 %, Западно-Южно-Американская — 7 %. Колебание индекса общности Чекановского — Сёренсена от 0.27 до 0.41 между провинциями и субпровинциями Альпийско-Меланезийско-Китайской области свидетельствует о стабильных биогеографических связях между ними.
Такие же стабильные биогеографические связи существовали между всеми биогеографическими областями, за исключением Альпийско-Меланезийско-Китайской и Сибирско-Аляскинской, в которых индекс общности Чекановского-Сёренсена составляет 0.18, что свидетельствует о наличии между ними каких-то физических (возможно, ландшафтных или батиметрических) барьеров, препятствующих обмену фауной брахиопод.
Исследования проведены в соответствии с планом НИР ИГАБМ СО РАН (проекты 0381-2016-0001 и 0381-2016-0002) и при частичной финансовой поддержке РФФИ (грант РФФИ-Восток № 15-45-05024) .
Список литературы Глобальная палеогеография и палеобиогеография тоарского века (ранняя юра) по брахиоподам
- Алексеева Р. Е. Палеозоогеографические области Евразии раннего девона (по брахиоподам) // Палеонтол. журнал. 1992. № 1. C. 3-13.
- Баранов В. В. Кризис новой глобальной тектоники литосферных плит и палеобиогеография пржидольских брахиопод северных регионов Евразии и Северной Америки // Вестн. Госкомгеологии. 2016. № 1(15). С. 77-90.
- Баранов В. В., Гриненко В. С. Глобальная палеогеография и биогеография среднего палеозоя и позднего триаса по брахиоподам // Геология и минерально-сырьевые ресурсы Северо-Востока России: Материалы VIII Всерос. науч.-практ. конф., 18-20 апреля 2018, г. Якутск, 2018. Т. II. С. 16-20.
- Блюман Б. А. Актуальные вопросы геологии океанов и континентов. СПб.: Изд-во ВСЕГЕИ. 397 c.
- Блюман Б. А. Выветривание базальтов и несогласия в коре океанов: возможные геодинамические следствия // Регионал. геол. и металлогения. CПб.: Изд-во ВСЕГЕИ, 2008. № 35. C. 72-86.
