Глутаматэргическая модуляция кардиореспираторных эффектов гипоксии

Автор: Буй Тхи Хыонг, Александров Вячеслав Георгиевич, Александрова Нина Павловна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 29, 2013 года.
Бесплатный доступ
В экспериментах на крысах, анестезированных уретаном, исследовалось влияние церебрального уровня глутамата на кардиореспираторные эффекты гипоксической стимуляции. Предъявление газовой смеси содержащей 10% О2 в N2 приводило к рефлекторному снижению среднего артериального давления, увеличению частоты сердечных сокращений, учащению дыхания и увеличению дыхательного объёма. Повышение церебрального уровня глутамата при его церебровентрикулярном введении резко ослабляло или устраняло эти эффекты. Таким образом, установлено, что церебральный уровень глутамата является фактором, который может оказывать модулирующее влияние на рефлекторные механизмы хеморецепторной регуляции дыхания и кровообращения.
Глутамат, гипоксия, дыхание, кровообращение, хеморецепторная регуляция
Короткий адрес: https://sciup.org/146116430
IDR: 146116430 | УДК: 612.217+612.766
Glutamatergic modulation of the cardiorespiratory effects of hypoxia
The influence of a glutamate cerebral level on cardiorespiratory effects of hypoxic stimulation had been performed in experiments on the anesthetized rats. Presentation of a gas mix containing 10% of O2 in N2 led to decrease in average arterial pressure, and increase in heart rate, and elevation both in breathing rate and in tidal volume. These effects disappeared after cerebroventicular microinjection of glutamate. Thus, it has been established that cerebral level of glutamate is a factor which can modulate ventilatory and cardiocirculatory chemoreflex.
Текст научной статьи Глутаматэргическая модуляция кардиореспираторных эффектов гипоксии
Введение. Изучение рефлекторных механизмов, реализующих эффекты гипоксии на функции дыхания и кровообращения, является перспективным направлением исследований в области нейрофизиологии висцеральных систем. Известно, что понижение напряжения кислорода в артериальной крови приводит к рефлекторным реакциям кардиоваскулярной и респираторной систем [7]. С другой стороны в состоянии гипоксии наблюдается повышение церебрального уровня глутамат-аниона L-глутаминовой кислоты, который является одним из важнейших нейромедиаторов центральной нервной системы [4]. Вместе с тем установлено, что величина вентиляторного ответа на гипоксию зависит от уровня эндогенного глутамата в ядре солитарного тракта, в котором происходит замыкание рефлекторных дуг ряда кардиореспираторных рефлексов [5]. Это позволяет предположить, что церебральный уровень глутамата является фактором, оказывающим модулирующее влияние на реализацию кардиореспираторных хеморефлекторных механизмов.
Наши предыдущие исследования показали, что повышение церебрального уровня глутамата приводит к усилению инспираторнотормозящего рефлекса Геринга-Брейера [2], ослабляет барорефлекторную чувствительность [3]. Тем самым было установлено, что церебральный уровень глутамата является фактором, оказывающим модулирующее влияние на механорецепторные механизмы регуляции кардиореспираторной системы. Цель настоящего исследования заключалась в изучении возможного влияния повышенного церебрального уровня глутамата на хеморецепторные механизмы регуляции, которые обеспечивают реакцию кардиореспираторной системы на гипоксический стимул.
Материал и методика . Эксперименты были проведены на трахеостомированных спонтанно дышащих крысах линии Wistar (n=7, самцы, вес 250-300 г), анестезированных уретаном (1250 мг/кг, в/б). В ходе эксперимента регистриовалась пневмотахограмма и артериальное давление, производилась микроинъекция раствора глутамата в боковые желудочки мозга. При подготовке животного к эксперименту производилась трахеостомия. В б едренную артерию устанавливался катетер, который представлял собой полиэтиленовую трубку, заполненную физиологическим раствором, содержащим гепарин в концентрации 2500 ЕД/мл. На черепе размечалось и рассверливалось отверстие для введения микроинъектора. В прямой кишке устанавливался термодатчик и включалась система термостатирования животного.
К трахеостомической трубке последовательно присоединялась пневмометрическая трубка и клапанная коробка. Коробка обеспечивала вдох из мешка, заполненного воздухом или газовой смесью, и свободный выдох. Пневмометрическая трубка подключалась к пневмотахометру ML141 (ADInstruments, Австралия). Артериальный катетер присоединялся к переходной камере, установленной на датчике давления. Камера, также как и катетер, предварительно заполнялась раствором гепарина. Датчик давления присоединялся к входу мостового усилителя ML224 (ADInstruments, Австралия) и калибровался в мм рт. ст. при помощи манометра . Сигналы пневмотахограммы и артериального давления подавались на вход устройства сбора данных PowerLab8/30 ( производство ADInstruments, Австралия) и обрабатывались в ходе эксперимента с помощью пакета программ LabChart 6.0. По сигналу пневмотахограммы рассчитывался дыхательный объём (ДО) и частота дыхательных движений (ЧДД), по сигналу артериального давления – среднее артериальное давление (АД ср ) и частота сердечных сокращений (ЧСС).
Микроинъекции раствора глутамата производились при помощи шприца Гамильтона, соединённого с микроинъектором, который представлял собой стальную трубку с наружным диаметром 150 мкм. Микроинъектор погружался в правый боковой желудочек при помощи стереотаксического аппарата. Координаты трепанационного отверстия и глубина погружения инъектора определялись по атласу мозга крысы и составляли 0,5 мм ростральнее уровня bregma, 1,5 мм латеральнее
Вестник ТвГУ. Серия "Биология и экология". 2013. Выпуск 29. № 2 средней линии, 3,5-4,0 мм от поверхности черепа. Через микроинъектор, в правый боковой желудочек мозга со скоростью 1 мкл/мин вводилось 5 мкл раствора, содержащего 6 мкМ глутамата.
После того, как температура тела животного стабилизировалась на уровне 36,8-37,0°С, производили пробную запись регистрируемых процессов и приступали к эксперименту. Общая длительность эксперимента составляла 90 мин, время дыхания гипоксической смесью при одном предъявлении равнялось 2,5 мин. Первые 40 минэксперимента животное дышало гипоксической смесью каждые 10 минут. На 40-й минуте приступали к введению глутамата, и после этого гипоксическая смесь предъявлялась на 3-й, 5-й, 10-й минуте, а затем вновь каждые 10 минут до конца эксперимента. Для количественной оценки эффектов действия гипоксической смеси величина всех учитывавшихся параметров, зарегистрированных на фоне действия смеси, выражалась в процентах к величине тех же параметров, зарегистрированных непосредственно перед её подачей. Статистическая обработка полученных данных проводилась средствами пакета Microsoft Excel. Вычислялась средняя величина регистрируемых параметров и ошибка среднего. Для оценки достоверности выявленных различий использовался однофакторный дисперсионный анализ. Различия считались достоверными при Р<0,05.
Результаты и обсуждение. Эксперименты показали, что дыхание гипоксической смесью приводит к характерным изменениям активности кардиореспираторной системы. Так, АДср уменьшалось, в среднем на 37±3,5 мм рт. ст., а ЧСС возрастало на 37±7,2 уд/мин. Происходило увеличение ДО на 0,5±0,05 мл, и ЧДД - на 26±4,0 ц/мин. На 41-й минуте начиналось введение глутамата, который оказывал возбуждающее действие на кардиореспираторную систему. Значения всех параметров достигали максимальных величин на 50-й минуте эксперимента, то есть через 10 минут после начала введения (Табл. 1). Затем происходило постепенное снижение активности кардиореспираторной системы.
Таблица 1
Изменение параметров активности кардиореспираторной системы под влиянием церебровентрикулярных микроинъекций глутамата
|
Параметр, размерность |
Время прошедшее от начала опыта (мин) |
||
|
40 |
50 |
90 |
|
|
АД ср , мм рт. ст. |
92±6,6 |
154±17,0* |
94±9,50 |
|
ЧСС, уд/мин |
402±2,10 |
492±15,5* |
421±24,4 |
|
ДО, мл |
02,0±0,09 |
4,3±0,11* |
2,5±0,11 |
|
ЧДД, цикл/мин |
0140±13,8 |
161±19,9* |
127±21,2 |
Примечание. * – достоверные отличия от исходных значений, зарегистрированных на 40-й мин непосредственно перед введением глутамата (Р<0,05).
I 70
20 40 мин 60 80 100
S?
о 120
В
40 60 80 100
мин
мин
мин
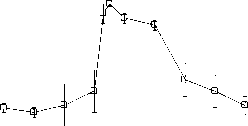
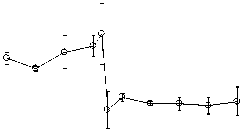
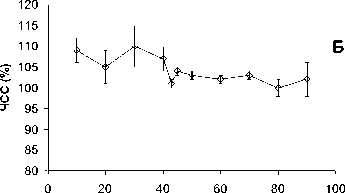
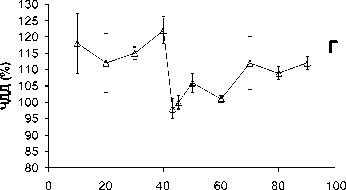
Рис . 1. Ослабление гипоксической реакции при повышении церебрального уровня глутамата.
Объяснения в тексте
Дыхание гипоксической смесью обычно приводило к падению АД ср до величин, составлявших 55-65% от его текущего значения (рис. 1А). Повышение церебрального уровня глутамата приводило к тому, что гипоксия переставала влиять на величину АД ср (рис. 1А). В течение первых 20 минут после начала введения глутамата дыхание гипоксической смесью не вызывало снижения АД ср . Затем этот эффект постепенно ослабевал. Реакция на гипоксию восстанавливалась и к концу эксперимента была такой же, как в начале эксперимента, до введения глутамата (рис. 1А). Гипоксия вызывала также небольшое, но статистически достоверное увеличение ЧСС, которого не происходило на фоне действия глутамата (Рис. 1Б). Повышение церебрального уровня глутамата влияло и на гипоксическую реакцию респираторной системы. До введения глутамата предъявление гипоксической смеси вызывало увеличение ДО на 20-30% по сравнению с текущими величинами. Повышение церебрального уровня глутамата устраняло этот эффект, причём необратимо (рис. 1В). Гипоксия вызывала также увеличение ЧДД на 15-25% по сравнению с текущим значением этого параметра. Этот эффект также исчезал после введения глутамата, постепенно восстанавливаясь к концу эксперимента (рис. 1Г).
Таким образом, результаты наших экспериментов свидетельствуют о том, что повышение церебрального уровня глутамата устраняло или резко ослабляло хеморецепторную реакцию кардиореспираторной системы анестезированной крысы на гипоксический стимул.
В наших предыдущих исследованиях было показано, что повышение церебрального уровня глутамата оказывает активирующее влияние на состояние кардиореспираторной системы, увеличивая частоту дыхания, электрическую активность диафрагмы, повышая артериальное давление и частоту сердечных сокращений [2; 3]. В тех же экспериментах было впервые установлено, что церебральный глутамат может оказывать модулирующее влияние на некоторые механорефлекторные механизмы регуляции дыхания и кровообращения. В частности, повышение церебрального уровня глутамата приводило к усилению инспираторно-тормозящего рефлекса Геринга-Брейера [2] и ослаблению барорефлекса [3]. Сходное действие на параметры активности респираторной системы оказывал миметик глутамата N-метил-D-аспартат (NMDA), который увеличивает частоту дыхания и дыхательный объём, а также усиливает инспираторнотормозящий и экспираторно-облегчающий рефлексы Геринга-Брейера [1]. Как известно, афферентным звеном рефлексов Геринга-Брейера являются первичные афференты, происходящие от медленноадаптирующихся рецепторов растяжения лёгких [6], а барорефлекс запускается рецепторами растяжения, расположенными в дуге аорты [9]. Поэтому полученные нами ранее результаты показали, что церебральный уровень глутамата является фактором, оказывающим модулирующее влияние на механорецепторные механизмы регуляции кардиореспираторной системы. Результаты настоящего исследования доказывают, что тот же фактор оказывает модулирующее влияние и на хеморецепторные рефлекторные механизмы, регулирующие активность кардиореспираторной системы. Это доказывается тем, что повышение церебрального уровня глутамата резко ослабляло все исследованные нами компоненты гипоксического ответа. Вместе с тем, следует отметить, что способность кардиореспираторной системы реагировать на гипоксию понижением артериального давления и учащением дыхания постепенно восстанавливалась, а реакции в виде увеличения дыхательного объёма и учащения дыхания не проявлялись до конца эксперимента. По-видимому, это связано с тем, что при повышении церебрального уровня глутамата происходит активация глутаматных рецепторов разных типов. Вероятно, что рецепторы NMDA-типа обеспечивают быструю реакцию рефлекторных механизмов кардиореспираторной системы на повышение уровня церебрального глутамата. В наших экспериментах эта реакция развивалась в течение 3-5 минут после начала микроинъекции [1]. Вместе с тем, активация NMDA-рецепторов может оказывать косвенное влияние на механизмы рефлекторной регуляции кардиореспираторной системы. Известно, в частности, что NMDA вызывает усиление экспрессии нейронами циклооксигеназы-2, что стимулирует выработку простагландинов [10],
Вестник ТвГУ. Серия "Биология и экология". 2013. Выпуск 29. № 2
которые, в свою очередь обладают широким спектром действия, в том числе и на механизмы центральной регуляции висцеральных систем [8].
