Глутаминовая и y-аминомасляная кислоты в камбиальной зоне сосны обыкновенной под влиянием фитоценотического стресса

Автор: Дейч К.О.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Трибуна молодых ученых
Статья в выпуске: 6, 2012 года.
Бесплатный доступ
В статье рассматривается влияние фитоценотического стресса на состав свободных аминокислот в камбиольной зоне сосны обыкновенной с разной густотой насаждения.
Глутаминовая кислота, γ-аминомасляная кислота, сосна, фитоценотический стресс
Короткий адрес: https://sciup.org/14082495
IDR: 14082495 | УДК: 581.524.12
Glutamic and y-amino butyric acids in the scotch pine cambial zone under the phytocenotic stress influence
The phytocenotic stress influence on the free amino acid composition in the Scotch pine cambial zone with various stand density is considered in the article.
Текст научной статьи Глутаминовая и y-аминомасляная кислоты в камбиальной зоне сосны обыкновенной под влиянием фитоценотического стресса
Рост древесных растений в лесу происходит на фоне разнообразных взаимодействий между ними, основными из которых являются конкурентные отношения, в частности внутривидовые. Степень и характер взаимодействий растений в сообществе зависят от густоты ценоза [6]. Густота является определяющим фи-тоценотическим фактором в жизни лесных экосистем и имеет важное хозяйственное значение, она определяет структурный состав и продуктивность молодняков. Взаимное угнетение растений в высокополнотных насаждениях увеличивается в чистых, одновозрастных насаждениях, вследствие чего всегда встречаются господствующие, угнетенные и переходные между ними экземпляры [1, 2]. Реакцию на усиленную конкуренцию деревьев в ценозе можно рассматривать как фитоценотический стресс [3].
Одним из индикаторов стрессового состояния растения при этом может служить изменение аминокислотного состава [4, 5]. Аминокислоты в растениях выполняют важную роль, участвуя в синтезе многих соединений.
Важное место в ответных метаболических реакциях растений на стресс занимают превращения свободных аминокислот, среди которых ключевая роль принадлежит глутаминовой кислоте и её производным.
Так как большая часть азотного цикла растений связана с синтезом глутаминовой кислоты и её производных, то по изменению состава аминокислот глутаминового семейства можно судить о направленности физиологических процессов и накоплении биомассы у разных деревьев в ценозе.
Глутаминовая кислота – важнейшая протеиногенная заменимая аминокислота, входящая в состав практически всех природных белков и других биологически активных веществ (глутатион, фолиевая кислота, фосфатиды). В свободном состоянии присутствует во всех тканях живых организмов, занимает ключевое положение в азотистом обмене растений.
γ-аминомасляная кислота (ГАМК) – непротеиногенная аминокислота, постоянно обнаруживаемая в тканях растений, иногда в концентрациях, значительно превышающих концентрации свободных протеиногенных аминокислот [5, 7]. ГАМК является одним из основных компонентов глутаминового семейства, в состав которого входят глутамин, глутаминовая кислота, аргинин, пролин, цитруллин и орнитин. Из них протеиногенными, которые входят в состав генетического кода и включаются в белки в процессе трансляции, являются – пролин, глутамин, глутаминовая кислота, аргинин.
К непротеиногенным свободным аминокислотам относят ГАМК [7], а также орнитин и цитруллин [8], которые совместно с протеиногенным аргинином входят в орнитиновый цикл – важнейший путь ассимиляции аммиака в растении.
Целью исследования явилось изучение метаболизма γ-аминомасляной и глутаминовой кислот в камбиальной зоне сосны обыкновенной в условиях внутривидовой конкуренции разной степени напряженности.
Объект и методы исследования. Объектом исследования служил 6–7-летний молодняк сосны обыкновенной ( Pinus silvestris L.), растущий на площади, вышедшей из сельскохозяйственного пользования в Красноярской лесостепи. Опытные экземпляры отбирались по положению в биогруппах: свободно растущие (1 экз . м-2), слабо загущенные (10 экз . м-2), сильно загущенные (40 экз . м-2). Для анализа в течение сезона вегетации 4 раза (июнь, июль, август, сентябрь) из разных биогрупп отбирались деревья, со стволиков которых удалялась кора с флоэмой, собирался слой камбия и неодревесневшей ксилемы и немедленно замораживался при -80оС.
Для определения свободных аминокислот 2 г навески тканей экстрагировали 80%-м этанолом, экстракт выпаривали, растворяли в воде, растворимые белки осаждали хлороформом, очищенный экстракт освобождали от углеводов на колонке с катионитом КУ-2, элюат концентрировали и проводили определение на аминокислотном анализаторе ААА-339, результаты рассчитывали по стандартной смеси аминокислот.
Результаты исследования и их обсуждение. Морфометрические характеристики деревьев показали, что с усилением давления внутривидовой конкуренции происходило значительное снижение радиального и линейного годичного прироста, причем различия в диаметре ствола в выделенных группах разной густоты достигали пятикратной величины из-за почти пятикратного уменьшения числа и размеров трахеид в годичном слое. При этом показатели плотности и влажности древесины увеличивались (табл.1).
Таблица 1
Морфометрические параметры модельных деревьев сосны обыкновенной при разной степени загущенности насаждения
|
Параметр |
Густота, шт∙м-2 |
||
|
1 |
10 |
40 |
|
|
Высота, см |
257 ±11 |
230 ± 5 |
95 ± 3 |
|
Диаметр корневой шейки, см |
6,1 ± 0,1 |
4,1 ± 0,3 |
1,2 ± 0,1 |
|
Ширина годичного кольца, мм |
3,3 ± 0,2 |
1,9 ± 0,1 |
0,9 ± 0,1 |
|
Длина верхушечного побега, см |
52 ± 5,0 |
46 ± 1,7 |
26,7 ± 1,6 |
|
Длина хвои, см |
8,4 ± 0,1 |
6,4 ± 0,1 |
5,5 ± 1,1 |
|
Число трахеид в годичном слое, шт. |
122 ± 7,1 |
63 ± 2,7 |
27 ± 2,1 |
|
Влажность древесины, % |
38,7± 0,02 |
38,9± 0,02 |
46,9 ± 0,01 |
|
Плотность древесины, г∙см-3 |
0,32 ± 0,02 |
0,38 ± 0,03 |
0,44 ± 0,09 |
Всего в камбиальной зоне сосны обыкновенной было идентифицировано 28 свободных аминокислот (табл. 2).
Незначительным содержанием (менее 0,1% от суммы всех идентифицированных аминокислот) отмечена α-аминоадипиновая кислота вне зависимости от месяца или биогруппы деревьев (табл. 2).
Таблица 2
Содержание свободных аминокислот в камбиальной зоне ствола сосны обыкновенной за период вегетации при различной густоте насаждения, % от суммы свободных аминокислот
|
Аминокислоты |
1экз∙м-2 |
10экз∙м-2 |
40экз∙м-2 |
|||||||||||
|
03 июн |
23 июн |
13 июл |
04 авг |
26 авг |
03 июн |
23 июн |
13 июл |
04 авг |
26 авг |
23 июн |
13 июл |
04 авг |
26 авг |
|
|
Цистеиновая |
0,25 |
0,31 |
0,38 |
0,25 |
0,28 |
0,19 |
0,30 |
0,13 |
0,21 |
0,25 |
0,22 |
0,12 |
0,35 |
0,59 |
|
Аспарагиновая |
0,32 |
0,35 |
0,13 |
0,15 |
0,24 |
0,30 |
0,18 |
0,42 |
0,12 |
0,21 |
0,29 |
0,17 |
0,13 |
1,55 |
|
Треонин |
1,29 |
2,23 |
1,79 |
0,69 |
0,88 |
2,55 |
1,52 |
0,78 |
0,84 |
1,29 |
2,08 |
2,25 |
1,69 |
2,11 |
|
Серин |
3,85 |
4,50 |
1,95 |
2,16 |
2,23 |
5,06 |
3,21 |
1,98 |
2,32 |
2,57 |
2,76 |
3,00 |
3,67 |
4,64 |
|
Аспарагин |
0,55 |
0,54 |
1,16 |
0,44 |
0,42 |
0,59 |
0,56 |
0,25 |
0,40 |
0,51 |
0,67 |
0,58 |
0,60 |
1,48 |
|
Глутаминовая |
1,50 |
1,98 |
1,09 |
0,93 |
1,93 |
4,55 |
1,78 |
0,87 |
0,31 |
1,02 |
0,59 |
0,53 |
0,95 |
3,28 |
|
Глутамин |
3,56 |
1,60 |
0,84 |
1,10 |
2,34 |
0,86 |
1,04 |
1,55 |
0,82 |
2,17 |
1,15 |
1,77 |
1,14 |
3,78 |
|
α-аминоадипиновая |
0,07 |
0,01 |
0,01 |
0,01 |
0,01 |
0,02 |
0,03 |
0,01 |
0,01 |
0,01 |
0,01 |
0,01 |
0,01 |
0,02 |
|
Гидроксипролин |
1,50 |
0,16 |
0,43 |
0,10 |
0,10 |
1,12 |
0,11 |
0,25 |
0,38 |
0,18 |
0,42 |
1,02 |
0,13 |
0,15 |
|
Пролин |
0,77 |
0,84 |
0,18 |
1,05 |
0,80 |
1,40 |
1,01 |
0,60 |
0,67 |
1,35 |
1,01 |
1,19 |
1,06 |
2,15 |
|
Глицин |
0,93 |
1,54 |
1,64 |
0,92 |
0,94 |
1,93 |
0,90 |
0,51 |
0,81 |
0,83 |
1,09 |
0,82 |
1,21 |
1,32 |
|
Аланин |
4,50 |
1,62 |
0,78 |
1,13 |
2,06 |
0,94 |
1,29 |
0,55 |
0,21 |
0,29 |
0,15 |
0,15 |
0,23 |
2,09 |
|
Цитруллин |
0,28 |
0,54 |
2,43 |
0,46 |
0,25 |
3,02 |
0,36 |
0,18 |
0,20 |
0,06 |
0,59 |
0,13 |
0,30 |
0,54 |
|
α-аминомасляная |
1,83 |
1,65 |
4,77 |
2,24 |
0,79 |
2,02 |
3,98 |
1,44 |
0,78 |
0,23 |
1,21 |
0,24 |
0,36 |
0,02 |
|
Валин |
2,46 |
3,16 |
9,59 |
1,13 |
1,25 |
1,47 |
2,54 |
1,34 |
1,05 |
1,69 |
2,21 |
2,00 |
1,28 |
3,34 |
|
Цистеин |
0,12 |
0,40 |
0,06 |
0,03 |
0,32 |
0,12 |
0,23 |
0,01 |
0,17 |
0,22 |
0,07 |
0,09 |
0,36 |
0,25 |
|
Цистатионин |
0,01 |
0,01 |
0,03 |
0,03 |
0,01 |
0,01 |
0,01 |
0,01 |
0,14 |
0,01 |
0,01 |
0,00 |
0,03 |
0,08 |
|
Метионин |
0,01 |
0,00 |
0,11 |
0,02 |
0,01 |
0,01 |
0,00 |
0,00 |
0,03 |
0,03 |
0,00 |
0,00 |
0,01 |
0,04 |
|
Изолейцин |
0,91 |
1,10 |
0,53 |
0,51 |
0,71 |
1,21 |
0,91 |
0,46 |
0,50 |
0,84 |
0,89 |
0,88 |
0,75 |
1,59 |
|
Лейцин |
2,19 |
1,96 |
0,90 |
0,67 |
1,05 |
2,92 |
1,60 |
0,81 |
0,76 |
1,58 |
2,17 |
1,76 |
1,64 |
2,82 |
|
Тирозин |
0,32 |
1,24 |
0,01 |
0,23 |
0,24 |
0,41 |
0,27 |
0,14 |
0,26 |
0,73 |
0,51 |
0,36 |
0,32 |
1,02 |
|
Фенилаланин |
0,53 |
0,68 |
0,14 |
0,42 |
0,40 |
0,23 |
0,47 |
0,26 |
0,33 |
0,68 |
0,83 |
0,64 |
0,44 |
1,20 |
|
β-аланин |
2,84 |
7,08 |
2,15 |
2,07 |
1,91 |
4,30 |
1,47 |
1,84 |
1,98 |
0,01 |
3,29 |
4,10 |
4,11 |
0,02 |
|
γ-аминомасляная |
66,68 |
60,55 |
67,15 |
81,19 |
79,38 |
57,48 |
72,47 |
83,51 |
85,09 |
80,72 |
73,52 |
73,41 |
76,44 |
60,39 |
|
Орнитин |
0,02 |
0,00 |
0,00 |
0,03 |
0,09 |
0,01 |
0,00 |
0,00 |
0,00 |
0,29 |
0,05 |
0,03 |
0,22 |
0,73 |
|
Лизин |
1,07 |
0,05 |
0,17 |
0,22 |
0,21 |
0,46 |
0,03 |
0,13 |
0,24 |
0,96 |
1,06 |
0,47 |
0,32 |
1,11 |
|
Гистидин |
1,28 |
5,00 |
1,15 |
1,18 |
0,79 |
5,84 |
2,82 |
1,46 |
0,73 |
0,53 |
2,54 |
3,56 |
1,04 |
1,32 |
|
Аргинин |
0,36 |
0,90 |
0,43 |
0,66 |
0,47 |
1,00 |
0,90 |
0,49 |
0,63 |
0,74 |
0,58 |
0,73 |
1,22 |
2,34 |
9°М Л^зреГУьУпншээд)
Серосодержащие аминокислоты (цистеин, цистатионин и метионин) за весь период вегетации не превышали значения 0,5% от суммы независимо от степени загущенности насаждения.
Немного большим содержанием на протяжении периода вегетации в камбиальной зоне сильно угнетенных деревьев отмечены аспарагиновая кислота и пролин (от 0,5 до 0,8–1,5% и от 1 до 2% соответственно). Повышенное содержание α-аминомасляной кислоты в начале и середине (от 2–4% до 1–0,2%) и снижение содержания к концу периода вегетации отмечено для всех выделенных биогрупп деревьев. Падением содержания к концу периода вегетации отмечен и гидроксипролин (от 1,5 до 0,2% для свободно растущей и менее загущенной биогруппы деревьев и от 0,4 до 0,1% для самой угнетенной биогруппы).
Содержание ароматических аминокислот семейства шикимата, которые предшествуют образованию лигнина в древесной ткани, фенилаланина и тирозина, колебалось от 0,2 до 0,6% в течение периода вегетации и повышалось до 1% к сентябрю у самых угнетенных деревьев. Похожая тенденция наблюдалась и среди аминокислот, с дополнительной аминогруппой – аспарагин, глутамин, лизин, повышенное содержание которых отмечено для свободно растущих деревьев в начале периода вегетации. Содержание их также повышалось в 1,5–3 раза (от 0,7 до 1,5%, от 1,2 до 3,8%), но уже к концу периода вегетации в биогруппе самых угнетенных деревьев сосны. Также повышенное содержание в конце периода вегетации у сильно угнетенных деревьев (в среднем 1–3%) наблюдалось для треонина и изолейцина.
Аминокислоты орнитинового цикла: орнитин и аргинин – имели повышенное содержание (от 0,02 до 0,7% от суммы всех свободных аминокислот) к концу периода вегетации для угнетенной группы растений. А содержание цитруллина (около 0,5%) повышалось только у свободно растущих деревьев в середине июля до 2,5 и 3 % к началу июня у среднеугнетенных деревьев.
Среди составляющих гликолатного цикла содержание серина за период вегетации снижалось в среднем с 4 до 2% у менее загущенных и повышалось от 3 до 4,5% у самой угнетенной биогруппы. Тогда как содержание глицина колебалось в пределах 1–2% за период вегетации во всех выделенных биогруппах.
Гетероциклическая аминокислота гистидин отличалась относительно высоким процентным содержанием у свободно растущих деревьев (5–6%) в начале периода активного роста и снижалась к сентябрю (до 1% в среднем от суммы). Также высокое относительное содержание в камбиальной зоне ствола отмечено для аминокислот семейства пирувата: аланин, β-аланин, валин, лейцин (до 4,5; 7,1; 9,6; 2,3% за период вегетации среди всех выделенных биогрупп деревьев), причем для аланина и β-аланина стабильное увеличение в начале и падение к концу периода вегетации для свободно растущих деревьев и накопление в ткани при увеличении степени загущенности в насаждении.
В целом содержание каждой из идентифицированных аминокислот не превышало показателя в 10% от суммы, кроме ГАМК, содержание которой колебалось от 60 до 80% от суммы.
Суммарное содержание свободных аминокислот постепенно снижалось к концу сезона с увеличением давления стресса. В камбиальной зоне свободно растущих деревьев происходило суммарное накопление свободных аминокислот в 2 раза от начала периода вегетации, а у слабо угнетенных накопления свободных аминокислот за весь период не отмечено (рис. 1). Тогда как в камбиальной зоне сильно угнетенных деревьев этот показатель снижался в 3,7 раза в сравнении с началом вегетационного периода.
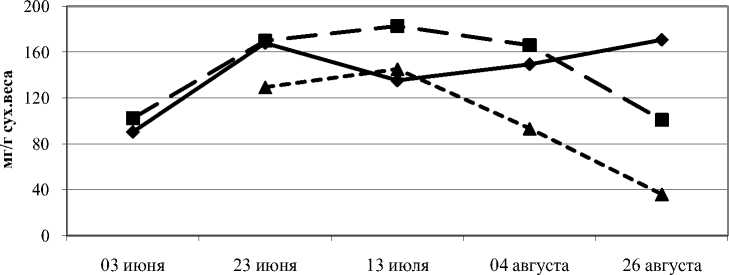
1 экз/м2 — ■-•10 экз/м2 * ^ ^^ ^ ^ 40 экз/м2
Рис. 1. Содержание свободных аминокислот в камбиальной зоне ствола сосны обыкновенной при действии внутривидовой конкуренции за период вегетации, в мг/г сухого веса
Мало менялось по сезону содержание ГАМК у сильно угнетенных деревьев, снижаясь в 1,3 раза к концу периода вегетации, тогда как у свободно растущих и слабо загущенных этот показатель повышался за сезон почти в 1,5 раза (рис. 2).
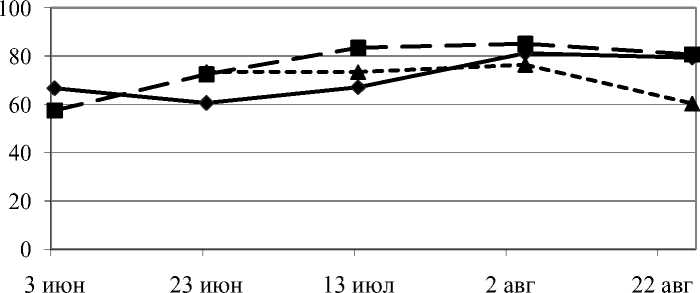
1 экз/м2 10 экз/м2 40 экз/м2
Рис. 2. Динамика Y-аминомасляной кислоты в камбиальной зоне ствола сосны обыкновенной при действии внутривидовой конкуренции за период вегетации, в % от общей суммы свободных аминокислот
Незначительно варьировало содержание глутаминовой кислоты в камбии свободно растущих деревьев по сезону (рис. 3). Однако в камбиальной зоне ствола слабо угнетенных деревьев было отмечено снижение в 4,5 раза этой аминокислоты и, напротив, в сильно угнетенных деревьях отмечено повышение в 5,5 раза содержания глутаминовой кислоты в сравнении с началом периода вегетации.
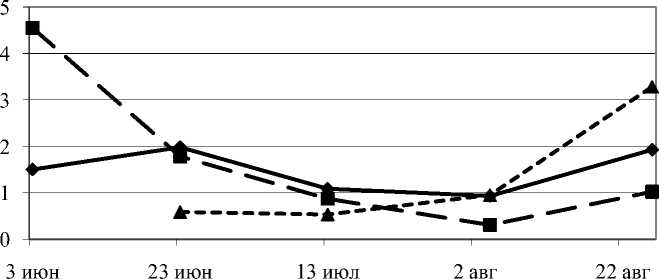
1 экз. м-2 10 экз. м-2 40 экз. м-2
Рис. 3. Динамика глутаминовой кислоты в камбиальной зоне ствола сосны обыкновенной при действии внутривидовой конкуренции за период вегетации, в % от общей суммы свободных аминокислот
На рисунке видно, что к концу сезона у сильно угнетенных деревьев доля ГАМК снижается значительно, притом, что доля семейства глутаминовой кислоты у тех же деревьев остается стабильной (рис.4).
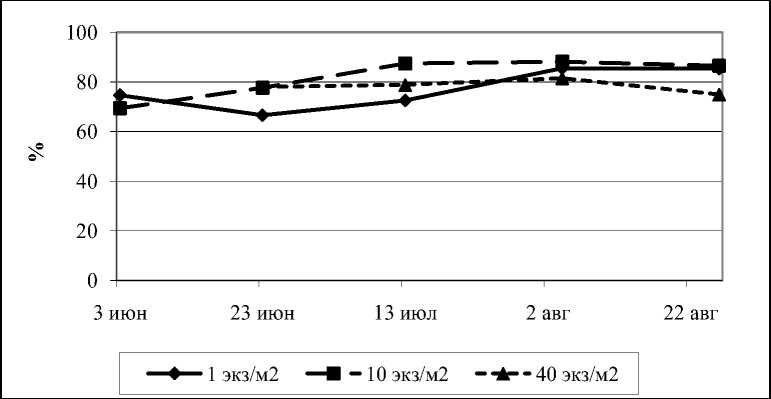
Рис. 4. Динамика аминокислот глутаминового семейства в камбиальной зоне ствола сосны обыкновенной при действии внутривидовой конкуренции за период вегетации, в % от общей суммы свободных аминокислот
Очевидно, что уровень ГАМК за период вегетации изменялся обратно уровню глутаминовой кислоты. Однако к концу сезона у сильно угнетенных деревьев доля ГАМК в общем пуле свободных аминокислот снижается значительнее в сравнении с долей семейства глутаминовой кислоты (рис. 3, 4).
Заключение. За период вегетации не происходило накопления свободных аминокислот среди деревьев самой угнетенной биогруппы. Содержание отдельных аминокислот было значительно повышенным к концу периода вегетации среди деревьев с максимальной густотой насаждения. Такие аминокислоты, как цистеиновая кислота, цистеин, серин, аспарагин, глутамин, пролин, изолейцин, тирозин, увеличились в 2–3 раза; аргинин, аспарагиновая и глутаминовая кислоты увеличились в 4–6 раз; аланин и орнитин увеличились десятикратно за период вегетации, по-видимому, компенсируя снижение содержания ГАМК в 1,2 раза в тех же деревьях. Содержание перечисленных выше аминокислот в биогруппах свободно растущих деревьев, наоборот, снижалось или оставалось относительно стабильным на общем фоне повышения общего содержания свободных аминокислот, тогда как содержание ГАМК в них повышалось в 1,2 раза к концу периода вегетации.
Относительно высокое содержание ГАМК в камбиальной зоне ствола отмечено во всех исследуемых группах деревьев. Вместе с тем для деревьев из биогруппы с максимальной густотой насаждения отмечено снижение доли ГАМК в сравнении с долей семейства глутаминовой кислоты в общем пуле свободных аминокислот к концу периода вегетации, что свидетельствует о перераспределении иона аммония среди других аминокислот семейства глутамата при усилении внутривидовой конкуренции к началу периода покоя.
Предполагается, что снижение относительного содержания ГАМК и повышение уровня глутаминовой кислоты в условиях жесткой внутривидовой конкуренции обусловлено снижением интенсивности ксилогенеза и дефицитом аминогрупп, освобождающихся в процессе лигнификации, в камбиальной зоне ствола деревьев самой загущенной биогруппы. Такое изменение содержания ГАМК в камбиальной зоне сосны обыкновенной может быть использовано в качестве индикатора состояния фитоценотического стресса деревьев сосны обыкновенной в разногустотных естественных насаждениях.
