Гнездящиеся птицы Приморского края: китайский ремез Remiz consobrinus

Автор: Ю.Н. Глущенко, Д.В. Коробов, И.М. Тиунов, В.Н. Сотников, Д.А. Беляев, И.Н. Коробова, В.П. Шохрин
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2596 т.35, 2026 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140314229
IDR: 140314229
Breeding birds of Primorsky Krai: the Chinese penduline tit Remiz consobrinus
Текст статьи Гнездящиеся птицы Приморского края: китайский ремез Remiz consobrinus
Статус . Китайский ремез Remiz consobrinus (Swinhoe, 1870) (рис. 1) является редким локально гнездящимся перелётным видом, однажды встреченным в зимний период. Это активно прогрессирующий недавний вселенец. На первых этапах заселения, когда китайский ремез был очень редким, его включили в Красную книгу Приморского края (Бур-ковский 2005), но позднее, ввиду многократного увеличения численности и расширения гнездового ареала, поступило предложение об исключении этого вида из региональной Красной книги (Глущенко и др. 2022), что и было сделано (Прозорова и др. 2025).
Историческая справка . Первая регистрация китайского ремеза в Приморском крае, согласно наблюдению Е.Э.Стоцкой, состоялась 22 мая 1980 на острове Фуругельма в заливе Петра Великого (рис. 2. 1) . Позднее пролётную группу этих птиц встретил В.Д.Куренков в начале октября второй половины 1980-х годов (точная датировка отсутствует) в пойме реки Раздольная у железнодорожной станции Сиреневка (Глущенко и др. 2014) (рис. 2. 2) .
Появление гнездящихся китайских ремезов в Приморье, вероятно, следует относить к стыку 1980-х и 1990-х годов. Первое гнездо (прошлогоднее) было обнаружено В.Н.Кубаниной в марте 1994 года на юге Ха-санского района в окрестностях сопки Голубиный утёс, а в 1996-1997 годах обследованное здесь поселение не превышало 6-10 пар (Бурковский 1998, 2005; Бурковский и др. 2014; Burkovskiy 1997). В последующие два года численность птиц увеличилась до 20-25 пар (Бурковский 2005). По своей структуре это поселение представляло собой рассеянную колонию, в которой, как показали наблюдения за мечеными птицами, все соседствующие особи регулярно контактировали друг с другом (Бурковский и др. 2014). Важно отметить, что в 1990-е годы китайские ремезы гнездились только вдоль морского побережья от Голубиного утёса через полуостров Краббе до посёлка Зарубино (рис. 2.3), а на побережьях бухты Экспедиции этот вид явно отсутствовал.

Рис. 1. Китайские ремезы Remiz consobrinus . 1-3 – самцы; 4-6 – самки; 7-9 – молодые птицы.
1 – восточное побережье озера Ханка, 27 мая 2012; 2 – Хасанский район, окрестности озера Птичье, 17 мая 2014; 3 – восточное побережье озера Ханка, 27 мая 2012; 4 – Спасский район, окрестности села Степное, 1 июня 2012; 5 – Хасанский район, окрестности озера Птичье, 17 мая 2014;
6 – там же, 22 мая 2014; 7-9 – западное побережье залива Петра Великого, подножье сопки Голубиный утёс, 21 августа 2008. Фото Д.В.Коробова
После 2000 года рост численности китайского ремеза продолжился, что привело к уплотнению гнездовой группировки и заселению побережий внутренних бухт в окрестностях посёлков Посьет и Краскино. С 2007 по 2010 год китайские ремезы заняли побережья ряда других бухт западной части залива Петра Великого, в частности, бухты Троицы, некоторых бухт материковой части восточного участка Дальневосточного государственного морского заповедника, а также бухт Бойсмана, Баклан и Северная (рис. 2.4). Общую численность вида к 2014 году оценивали в 400-450 условных пар, гнездящихся от границы с КНДР до устья реки Брусья на побережье Славянского залива (Глущенко и др. 2014; Gluschenko et al. 2014). К настоящему времени эта гнездовая группировка (назовём её хасанской) прерывисто заняла всё западное побережье залива Петра Великого и прилежащие приморские участки территории вплоть до северного побережья Амурского залива, где ремезы достоверно гнездятся в устьях рек Раздольная и Давыдовка (рис. 2.5).
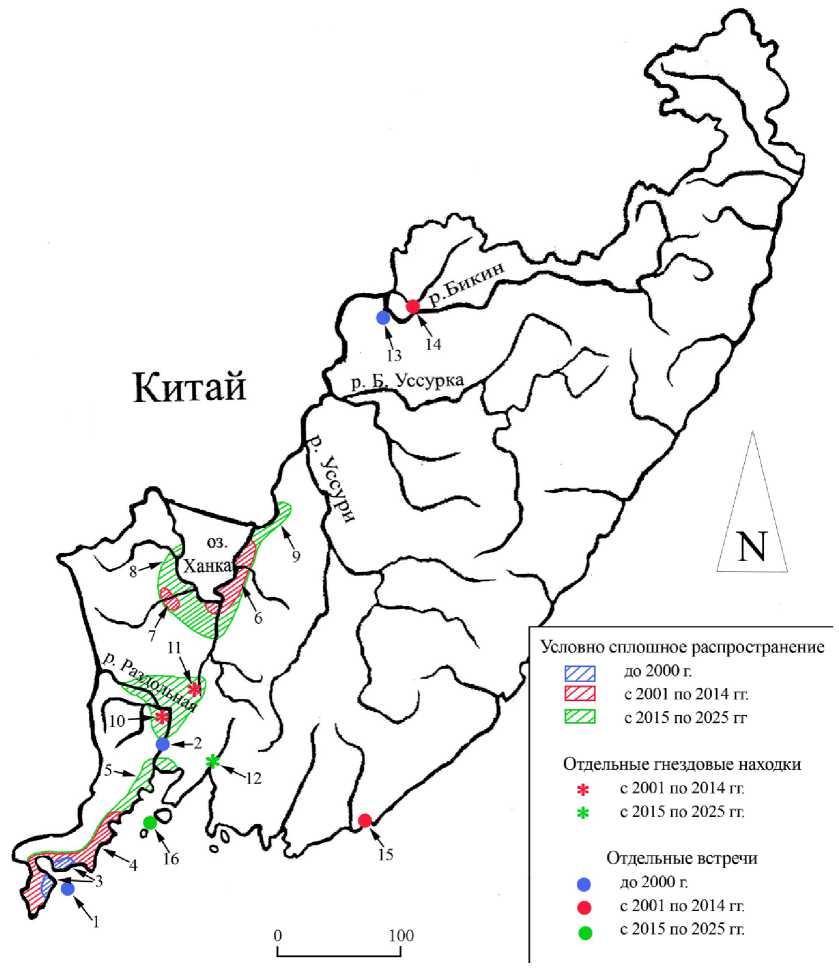
Рис. 2. Распространение китайского ремеза Remiz consobrinus в Приморском крае (пояснения в тексте)
На Приханкайской низменности китайского ремеза впервые отметил В.А.Нечаев 22 июня 2000, когда встретил самца, собиравшего растительный пух для строительства гнезда в окрестностях Берёзовых озёр вблизи юго-восточного побережья озера Ханка. Судя по находкам старых гнёзд, птицы размножались здесь в 2001 году, а в 2002 эту локальную группировку детально обследовали и насчитали до 35 гнездящихся пар (Глущенко и др. 2004). В последующие годы (до 2011 года включительно) численность гнездящихся на этом участке ремезов не претерпевала значительного роста, а в некоторые годы была даже заметно ниже, но район её обитания постепенно расширялся (рис. 2.6). В 2012-2013 годах группировка ремезов, гнездящихся на Приханкайской низменности, значительно увеличилась, и по нашей экспертной оценке составляла 100-150 условных пар. В 2014 году гнёзда ремезов обнаружили в западной части Приханкайской низменности (Волковская-Курдюкова, Курдюков 2014) (рис. 2.7), а к 2025 году ханкайская группировка локально заняла практически все участки российского сегмента Приханкайской низменности до низовий реки Комиссаровка (рис. 2.8).
В долине реки Сунгача активно вокализирующего и широко перемещающегося самца китайского ремеза наблюдали 8 июня 2014 в Кировском районе в окрестностях села Павло-Фёдоровка (Волковская-Кур-дюкова, Курдюков 2019). По нашим данным, с 2021 года на правобережье реки Сунгача, в частности, в окрестностях села Павло-Фёдоровка, а также в междуречье притоков Белая и Шмаковка и по реке Красная (рис. 2. 9) эти птицы регулярно гнездятся.
В 2010 году гнёзда китайских ремезов были обнаружены в китайском секторе Приханкайской низменности у озера Малая Ханка (Глущенко и др. 2010). Дополнительное обследование этой территории, проведённое нами в 2011 году, показало, что здесь этот вид в те годы был явно обильнее и шире распространён, нежели в российском секторе низменности (Глущенко и др. 2012). Это вполне закономерно, поскольку заселение здесь явно проходило в юго-восточном направлении, то есть с китайской территории на российскую.
Судя по находкам единичных гнёзд, с 2012 года китайские ремезы размножаются в низовьях Раздольной к югу от Уссурийска (рис. 2. 10) , а с 2014 года их гнёзда были найдены и несколько севернее – в окрестностях посёлка Новошахтинский Михайловского района (Глущенко и др. 2014; Gluschenko et al . 2014) (рис. 2. 11) . К 2025 году в бассейне вышеупомянутой реки возникла достаточно крупная и ныне широко распространённая уссурийская группировка.
Судя по всему, существующие в настоящее время хасанская, уссурийская и ханкайская гнездовые группировки китайских ремезов, которые изначально возникли независимо друг от друга, ввиду достаточно широкой экологической пластичности (в частности, благодаря переходу к гнездованию на ивах) вскоре территориально объединятся, заселив все подходящие для этого участки юго-западного Приморья. Единичные гнёзда с 2018 года мы находили в долине нижнего течения реки Шко- товка в окрестностях села Штыково (рис. 2.12). На данном этапе заселения ремезами территории края их распространение здесь носит разбро-сано-очаговый характер, при этом из-за недостаточной численности птиц большая часть пригодной для их гнездования территории ещё не заселена. В то же время на отдельных участках их плотность высокая, поскольку вследствие ряда поведенческих особенностей китайские ремезы формируют парцеллы, которые с некоторой натяжкой можно считать колониальными поселениями.
Китайских ремезов встречали и за пределами ныне известных районов гнездования. Так, зимующего самца наблюдали 18 февраля 1997 на крайнем северо-западе Приморского края в окрестностях города Лу-чегорск (Burkovskiy 1997) (рис. 2. 13) . В низовье реки Бикин 27 мая 2003 одну особь встретили в Пожарском районе в окрестностях села Верхний Перевал (Schaumburg et al . 2003) (рис. 2. 14) . В бухте Петрова в паутинную сеть 13 октября 2013 отловили 2 молодых ремезов, которые кочевали в смешанной стае синиц (Шохрин 2014) (рис. 2. 15) . На некоторых участках бассейна реки Уссури от посёлка Кировский до границы с Хабаровским краем, потенциально подходящих для гнездования ремезов, летом 2013 года мы провели рекогносцировочные обследования, но этих птиц не обнаружили. Можно предположить, что в тот период китайские ремезы если и гнездились здесь, то крайне редко и локально (Глущенко и др. 2014). Кроме этого, самца, активно рекламирующего занятую территорию, наблюдали 9 июля 2015 на острове Попова в северной части бухты Пограничная (рис. 2. 16) , однако при повторном посещении этого участка 6-7 июля 2017 ремезов здесь не обнаружили (Волковская-Кур-дюкова, Курдюков 2019).
Для Северной Кореи этот вид считался лишь пролётным, отмеченным в период с 3 апреля по 13 октября (Tomek 2002). Но в настоящее время при достаточно плотном гнездовании на приграничном российском пространстве представляется несомненным его гнездование и на соседней северокорейской территории, однако это требует фактического подтверждения.
Местообитания . На крайнем юго-западе Приморья китайские ремезы изначально гнездились в нижних частях склонов и у подножий невысоких сопок и возвышений, покрытых редколесьем с преобладанием дуба зубчатого Quercus dentata (рис. 3. 3) , населяя сильно разреженные древесные ассоциации, граничащие с открытыми пространствами. В последующие годы при увеличении численности гнездящейся группировки ремезы стали занимать и более возвышенные участки, распространяясь вверх по склонам. В настоящее время многие гнёзда ремезов ха-санской группировки располагаются также среди обширных выровненных сырых лугов и болот на разных деревьях и кустарниках, растущих, в частности, на невысоких гривах и приканальных дамбах.

Рис. 3. Варианты гнездовых биотопов китайского ремеза Remiz consobrinus на крайнем юго-западе Приморья (хасанская группировка). 1 – Хасанский район, окрестности посёлка Зарубино, 28 июля 2023; 2 – Хасанский район, окрестности посёлка Хасан, 26 августа 2023;
3 – Хасанский район, урочище Голубиный утёс, 7 мая 2023, фото Д.В.Коробова
В период становления хасанской гнездовой группировки наблюдалась явная или опосредованная привязанность птиц к морскому побережью, что проявлялось как в общей конфигурации поселения, так и в особенностях размещения гнёзд по мере отдаления от моря. Так, в 2011-

Рис. 4. Варианты гнездовых биотопов китайского ремеза Remiz consobrinus на Приханкайской низменности (ханкайская группировка). 1 – юго-восточное побережье озера Ханка, 10 мая 2016; 2 – северо-восточное побережье озера Ханка, 9 мая 2016; 3 – Спасский район, окрестности села Лебединое, 23 мая 2013. Фото Д.В.Коробова
2014 годах в бухте Алеут колония ремезов была лентообразно вытянута вдоль прибрежной полосы и в некоторой степени повторяла очертания береговой линии; большинство гнёзд было сосредоточено на расстоянии

Рис. 5. Варианты гнездовых биотопов китайского ремеза Remiz consobrinus в бассейне реки Раздольная (уссурийская группировка). 1 – Уссурийский городской округ, окрестности села Утёсное, 29 мая 2021; 2 – Октябрьский район, окрестности села Покровка, 8 июня 2022;
3 – окрестности Уссурийска, 28 октября 2023. Фото Д.А.Беляева.
10-800 м от береговой линии, а в одном случае гнездо находилось непосредственно над поверхностью моря (Бурковский и др. 2014). В настоящее время на крайнем юге Хасанского района удаление гнёзд ремезов от морского побережья может быть заметно больше. В частности, в 2011 году на участке морского побережья около посёлка Гвоздево их гнёзда обнаружили на удалении в 700-4700, в среднем в 1750 м ( n = 7) от берега моря (Волковская-Курдюкова, Курдюков 2019).
На начальном этапе заселения Приханкайской низменности ремезы чаще всего гнездились в сравнительно немногочисленных здесь строчных осинниках, растущих по дамбам каналов, некогда предназначавшихся для осушения этой переувлажнённой территории. Реже птицы устраивали гнёзда в куртинах осин, растущих на небольших возвышениях среди болот (рис. 4. 1) или на одиночных осинах, растущих по дамбам. Кроме того, несколько гнёзд нашли на древних береговых валах озера Ханка, заросших разнообразной древесно-кустарниковой растительностью (рис. 4. 2) и на ивах, растущих по дамбам каналов (Глущенко и др. 2004).

Рис. 6. Гнездо китайского ремеза Remiz consobrinus , расположенное на окраине города Уссурийска. 8 апреля 2023. Фото Ю.Н.Глущенко
В бассейне реки Раздольная китайские ремезы локально заселили главным образом малооблесённые участки речных долин (рис. 5). При размещении своих гнёзд они толерантны к присутствию людей. Так, некоторые их групповые поселения расположены на деревьях придорожных лесопосадок, где по трассам происходит активное движение автомобильного транспорта.
В одном из таких случаев гнездо находилось неподалёку от многоэтажной застройки Уссурийска (рис. 6). Следует отметить, что китайские ремезы мало осторожны и не опасаются заниматься строительством гнезда и кормлением птенцов даже тогда, когда наблюдатель находится открыто в нескольких метрах от гнезда.

Рис. 7. Китайские ремезы Remiz consobrinus в начале весенней миграции.
1 – самец, устье реки Раздольная, озеро Утиное, 29 апреля 2018, фото А.П.Рогаля;
2 – самка, Хасанский район, окрестности посёлка Зарубино, 10 мая 2024, фото О.Н.Васик
Весенний пролёт . Изучен слабо. Первых самцов на крайнем юге Приморья в разные годы наблюдали в начале третьей декады апреля или в последних числах этого месяца (рис. 7. 1) , а первых самок — в конце первой декады мая (рис. 7. 2) , но активная миграция продолжается в течение почти всего мая. Так, пролётную стаю ремезов, состоящую приблизительно из 20 особей, мы встретили 18 мая 2023 у посёлка Хасан, а 20 мая того же года у морского побережья Хасанского района наблюдали перемещение в северном направлении 5 небольших групп общей численностью 28 птиц.
На Приханкайской низменности весной первого самца мы наблюдали 1 мая 2012, хотя возможен и более ранний прилёт, который нами был пропущен из-за крайне нерегулярных посещений мест размножения ремезов. В начале второй декады этого месяца здесь отмечали и самок.
Гнездование . Уже в третьей декаде апреля прилетевшие первыми самцы занимают гнездовые участки. Так, 24 апреля 2023 у подножья
Голубиного утёса (западное побережье залива Петра Великого) встретили 2 самцов, которые, судя по поведению, делили территорию. Большинство самцов появляется на местах размножения в первой половине мая. Они активно поют (рис. 8) на занятом участке, порой вступают в

Рис. 8. Поющие самцы китайского ремеза Remiz consobrinus .
1 – Хасанский район, подножье Голубиного утёса, 18 мая 2014; 2-4 – восточное побережье озера Ханка, 27 мая 2012, фото Д.В.Коробова; 5 – северное побережье Амурского залива, устье реки Давыдовка,
2 июня 2024, фото В.М.Матюшина; 6 – Хасанский район, окрестности станции Рязановка,
1 июля 2018, фото А.В.Вялкова

Рис. 9. Самцы китайского ремеза Remiz consobrinus с материалом для строительства гнёзд.
1 – Хасанский район, подножье Голубиного утёса, 18 мая 2014; 2 – там же, 19 мая 2014; 3 – восточное побережье озера Ханка, 27 мая 2012; 4 – Хасанский район, окрестности посёлка Хасан, 3 июня 2016, фото Д.В.Коробова; 5 – Хасанский район, окрестности посёлка Зарубино, 11 июня 2019, фото В.П.Шохрина; 6 – там же, 19 июня 2020, фото И.А.Малыкиной в стычки с другими самцами. Инициатива начала строительства гнезда всегда принадлежит самцам (Бурковский и др. 2014). Вскоре после прилёта территориальные самцы приступают к сбору строительного материала (рис. 9) и началу строительства гнёзд (рис. 10), продолжая периодически петь, привлекая самок.
Стимулятором выбора гнездового дерева часто служит присутствие остатков прошлогоднего гнезда, при этом удалось выяснить, что старшие

Рис. 10. Самцы китайского ремеза Remiz consobrinus , строящие гнёзда.
1 – Хасанский район, окрестности станции Рязановка, 6 мая 2015, фото А.В.Вялкова; 2 – Хасанский район, окрестности посёлка Зарубино, 10 мая 2024, фото О.Н.Васик; 3 – Хасанский район, подножье Голубиного утёса, 18 мая 2023, фото Д.В.Коробова; 4,5 – Приханкайская низменность, окрестности озера Луповое, 27 мая 2012, фото Д.В.Коробова; 6 – Хасанский район, окрестности посёлка Зарубино, 11 июня 2019, фото В.П.Шохрина

Рис. 11. Гнёзда китайских ремезов Remiz consobrinus , расположенные близко одно к другому.
1 – Спасский район, окрестности села Луговое, 3 апреля 2022, фото О.М.Косолапова; 2 – Хасанский район, окрестности посёлка Хасан, 23 июня 2023, фото Ю.Н.Глущенко; 3 – там же, 31 мая 2016, фото И.И.Уколова особи хорошо помнят местоположение не только своих, но и соседних гнёзд предшествующего сезона и довольно часто строят новое гнездо на месте полностью разрушенного (Бурковский и др. 2014). В любом случае видеть расположенные рядом гнёзда разной давности (рис. 11.1,2) – явление обыденное, а в ряде случаев новое гнездо птицы устраивают как продолжение незаконченной старой постройки (рис. 11.3).
Благодаря индивидуальному мечению птиц удалось выяснить, что многие из гнездовых деревьев используются ремезами из года в год, часто разными парами, при этом нередко они восстанавливают прошлогодние недостроенные гнёзда, хотя многие такие попытки заканчиваются неудачей (Бурковский и др. 2014).
Первые 7 гнёзд китайских ремезов, обнаруженных в Хасанском районе в 1996 году, размещались на липах и 1 на иве (Burkovskiy 1997). В последующие годы птицы стали располагать свои постройки и на других деревьях: на яблоне маньчжурской Malus mandshurica , берёзе даурской Betula davurica , клёне приречном Acer ginnala , ольхе волосистой Alnus hirsuta , но по-прежнему отдавали предпочтение (77.3%) липам Tilia amurensis и T. mandshurica . В дальнейшем спектр выбора пород деревьев для гнездования значительно расширился, но при этом доля гнёзд, построенных на липах, хотя и снизилась, но всё равно оставалась высокой (Глущенко и др. 2014; Gluschenko et al . 2014). Позднее спектр деревьев и кустов, используемых ремезами для расположения гнёзд, продолжал расширяться: из 36 гнёзд, найденных на крайнем юго-западе Приморского края (в непосредственной близости от впервые найденного гнезда) в период с 17 по 25 мая 2014, на липах Tilia sp. располагались 9 гнёзд, на черёмухе азиатской Padus asiatica – 6, на яблонях Malus sp. – 5; по 3 гнезда находилось на клёнах Acer sp. и боярышниках Crataegus sp., по 2 – на бузине корейской Sambucus coreana , ильмах Ulmus sp. и ивах Salix sp. и по 1 гнезду – на леспедеце двухцветной Lespedeza bicolor , дубах монгольском Quercus mongolica и зубчатом Q. dentata и мелкоплоднике ольхолистном Micromeles alnifolia (Глущенко, Коробов 2014). К 2025 году бо ́ льшая часть найденных здесь гнёзд располагалась на ивах.
На Приханкайской низменности в 2002-2004 годах из 34 гнёзд ремезов на ивах были размещены 8.8%, а на осинах 73.5%, но к 2025 году на ивах размещались 31.6%, а на осинах – 53.8%. Переход к гнездованию на ивах имеет хорошие перспективы расселения и увеличения численности (Глущенко и др. 2014), поскольку распространение этих древесных пород среди водно-болотных угодий и низменностей Приморского края гораздо шире, чем, например, осинников, подходящих для гнездования китайских ремезов.
Для птиц уссурийской группировки ремезов бо́льшая часть найденных нами гнёзд ремезов располагалась на ильмах и ивах. Рассматривая всю территорию Приморского края, заселённую этими птицами на 2025 год, максимальное количество гнёзд размещалось на ивах, осинах и липах (табл. 1). Высота расположения гнёзд варьирует в очень широких пределах – от 0.85 до 11.0 м над поверхностью (земли или воды), в среднем (n = 291) 3.22 м (табл. 2).
Таблица 1. Места расположения гнёзд китайского ремеза Remiz consobrinus в Приморском крае (наши данные за 2002-2025 годы / Волковская-Курдюкова, Курдюков 2010, 2017, 2019; Burkovskiy 1997)
|
Место расположения гнезда |
Хасанская группировка |
Уссурийская группировка |
Ханкайская группировка |
Всего |
Доля, % |
|
Ива Salix sp. |
57/1 |
6/0 |
51/6 |
114/7 |
34.47 |
|
Осина Populus trеmula |
– |
5/0 |
85/3 |
90/3 |
26.50 |
|
Липа Tilia sp. |
22/14 |
– |
– |
22/14 |
10.26 |
|
Ильм Ulmus sp. |
5/0 |
7/0 |
12/0 |
24/0 |
6.84 |
|
Берёза Betula sp. |
4/0 |
5/0 |
9/0 |
18/0 |
5.13 |
|
Клён Acer sp. |
11/0 |
– |
– |
11/0 |
3.13 |
|
Яблоня Malus sp. |
8/0 |
3/0 |
– |
11/0 |
3.13 |
|
Черёмуха азиатская Padus asiatica |
8/1 |
1/0 |
– |
9/1 |
2.85 |
|
Ольха Alnus sp. |
4/0 |
1/0 |
– |
5/0 |
1.42 |
|
Боярышник Crataegus sp. |
4/0 |
– |
– |
4/0 |
1.14 |
|
Ясень Fraxinus sp. |
3/0 |
– |
– |
3/0 |
0.85 |
|
Бузина корейская Sambucus coreana |
2/0 |
– |
– |
2/0 |
0.57 |
|
Груша уссурийская Pyrus ussuriensis |
0/2 |
– |
– |
0/2 |
0.57 |
|
Дуб зубчатый Quercus dentata |
2/0 |
– |
– |
2/0 |
0.57 |
|
Дуб монгольский Quercus mongolica |
1/0 |
1/0 |
– |
2/0 |
0.57 |
|
Мелкоплодник ольхолистный Micromeles alnifolia |
2/0 |
– |
– |
2/0 |
0.57 |
|
Тополь Populus sp. |
– |
0/1 |
1/0 |
1/1 |
0.57 |
|
Актинидия коломикта Actinidia kolomikta |
1/0 |
– |
– |
1/0 |
0.28 |
|
Вишня Prunus sp. |
1/0 |
– |
– |
1/0 |
0.28 |
|
Леспедеца двухцветная Lespedeza bicolor |
1/0 |
– |
– |
1/0 |
0.28 |
|
Итого |
136/18 |
29/1 |
158/10 |
323/28 |
100.00 |
Таблица 2. Высота расположения гнёзд китайского ремеза Remiz consobrinus в Приморском крае
|
Число гнёзд |
Высота над землёй или водой |
Источник информации |
|
|
Пределы |
Среднее |
||
|
265 |
85-1100 |
325 |
Наши данные |
|
19 |
130-700 |
312 |
Волковская-Курдюкова, Курдюков 2010, 2017, 2019 |
|
7 |
– |
250 |
Burkovskiy 1997 |
|
291 |
85-1100 |
322.3 |
Всего |
Гнёзда обычно крепятся к кончикам тонких веток дистальных частей крон одиночно стоящих или опушечных деревьев или кустов (рис. 12). В случае, когда деревья растут на склоне, ремезы устраивают гнёзда в той части кроны, которая обращена вниз по склону (Burkovskiy 1997; наши данные).
Территориальные самцы, которые ещё не обзавелись самками, никогда не достраивают гнёзда до их полного завершения: чаще всего они оставляют их на стадии корзиночки («двух дыр»). После этого на разных этапах строительства к ним присоединяются самки, прилетающие с мест

Рис. 12. Типичные варианты расположения гнёзд китайских ремезов Remiz consobrinus .
1 – северное побережье Амурского залива, устье реки Давыдовка, 2 июня 2024, фото В.М.Матюшина;
2 – Хасанский район, окрестности посёлка Хасан, 23 июня 2023, фото Ю.Н.Глущенко; 3 – Уссурийский городской округ, долина реки Раздольная, 28 октября 2023; 4 – там же, 6 июня 2024, фото Д.А.Беляева;
5 – Хасанский район, подножье Голубиного утёса, 10 июня 2023; 6 – там же, 24 июня 2023, фото Д.В.Коробова

Рис. 13. Самки китайского ремеза Remiz consobrinus , участвующие в сборе материала и строительстве гнезда. 1 – залив Петра Великого, полуостров Гамова, 22 мая 2022, фото А.П.Рогаля; 2 – залив Петра Великого, мыс Островок Фальшивый, 18 мая 2014; 3, 4 – там же, 19 мая 2014; 5, 6 – восточный сектор Приханкайской низменности, окрестности озера Луповое, 1 июня 2012, фото Д.В.Коробова

Рис. 14. Самка китайского ремеза Remiz consobrinus , передающая самцу строительный материал для укладки в гнездо. Западное побережье залива Петра Великого, подножье Голубиного Утёса.
19 мая 2023. Фото Д.В.Коробова

Рис. 15. Несогласованное строительство гнезда самцом и самкой китайского ремеза Remiz consobrinus. Западное побережье залива Петра Великого, западное побережье залива Петра Великого, мыс Островок Фальшивый. 20 мая 2014. Фото Д.В.Коробова зимовок в среднем несколько позднее самцов. Они также участвуют в сборе материала (рис. 13.1) и строительстве, особенно активно на его последних стадиях, когда формируется трубка входа в гнездо (рис. 13.5) и натаскивается растительный пух внутрь постройки (рис. 13.6). Однако иногда принесённый материал самка передаёт самцу, который затем укладывает его в формирующуюся постройку (рис. 14).
Часть гнёзд ремезы бросают на разных стадиях строительства, а однажды, по-видимому, из-за несогласованности строительных работ, самка начала строить основу нового гнезда у нижнего края начатой самцом постройки. Таким образом, некоторое время происходило строительство оснований двух гнёзд одновременно (рис. 15).
Наиболее активное строительство гнёзд происходит во второй половине мая и в первой половине июня (Бурковский 2005; наши данные), а строительство самцами вторых гнёзд длится по меньшей мере до середины июля (табл. 3).
Таблица 3. Фенология размножения китайских ремезов Remiz consobrinus на разных участках Приморского края (наши данные за 2002-2024 годы / Бурковский 2005; Волковская-Курдюкова, Курдюков 2010, 2017, 2019; Burkovskiy 1997)
|
Период |
Число наблюдений на разных стадиях размножения |
||||||
|
Строительство гнезда |
Неполная кладка |
Полная кладка |
Голые птенцы |
Оперённые птенцы |
Слётки, выводки |
Всего |
|
|
1-15 мая |
11/0 |
– |
– |
– |
– |
– |
11/0 |
|
16-31 мая |
48/2 |
3/0 |
1/0 |
– |
– |
– |
52/2 |
|
1-15 июня |
25/2 |
3/0 |
10/0 |
– |
– |
– |
38/2 |
|
16-30 июня |
10/3 |
– |
6/0 |
– |
– |
– |
16/3 |
|
1-15 июля |
11/0 |
3/0 |
6/0 |
1/0 |
4/0 |
3/0 |
28/0 |
|
16-31 июля |
– |
– |
– |
– |
1/0 |
5/1 |
6/1 |
|
1-15 августа |
– |
– |
– |
– |
2/1 |
2/2 |
4/3 |
|
15-31 августа |
– |
– |
– |
1/0 |
1/0 |
2/0 |
4/0 |
|
Итого |
105/7 |
9/0 |
23/0 |
2/0 |
8/1 |
12/3 |
159/11 |
Процесс строительства гнезда можно условно разделить на 4 стадии, при этом участие в нём самки может начинаться на любой из них. На первой стадии, кода птицы формируют кольцо (рис. 16.1), крепящееся к дистальной части ветви, ремезы используют главным образом грубые растительные волокна. На втором этапе работ, когда на основе кольца ремезы конструируют «корзинку» (рис. 16.2), они начинают всё больше применять имеющийся поблизости разнообразный по происхождению, окраске и жёсткости «растительный пух». Из этого же материала птицы достраивают гнездо, доводя его сначала до полузакрытой стадии с двумя входами (рис. 16.3). Последняя стадия заключается в заделывании заднего входа (рис. 16.4) и в монтаже входной трубки, которая в окончательном варианте может иметь разную длину (рис. 17). На этой стадии ремезы натаскивают вовнутрь практически готового гнезда значительный объём растительного пуха, включая летучки одуванчика Taraxacum sp. и других растений. В зависимости от имеющегося в окрестностях материала окраска гнезда значительно варьирует от белёсой (особенно на входной трубке) до разных оттенков серого, бурого или рыжего в случае использования птицами «пуха» папоротника осмунды Osmunda sp.

Рис. 16. Стадии строительства гнёзд китайскими ремезами Remiz consobrinus . 1,2 – стадия кольца;
3,4 – стадия «корзинки»; 5 – стадия с двумя входами; 6 – стадия формирования входной трубки.
1 – западное побережье залива Петра Великого, подножье Голубиного утёса, 19 мая 2023; 2,3 – западное побережье залива Петра Великого, мыс Островок Фальшивый, 18-19 мая 2014, фото Д.В.Коробова;
4 – западное побережье залива Петра Великого, полуостров Гамова, 22 мая 2020, фото А.П.Рогаля;
5 – Приханкайская низменность, окрестности озера Луповое, 27 мая 2012, фото Д.В.Коробова;
6 – Хасанский район, окрестности посёлка Хасан, 2 июня 2016, фото Д.Н.Кочеткова

Рис. 17. Полностью готовые гнёзда китайских ремезов Remiz consobrinus .
1 – восточное побережье озера Ханка, 27 мая 2012; 2 – Спасский район, окрестности села Степное, 21 мая 2013, фото Д.В.Коробова; 3 – Хасанский район, окрестности посёлка Хасан, 17 июня 2014, фото Н.Н.Балацкого; 4 – там же, 24 июня 2023, фото Ю.Н.Глущенко; 5 – Хасанский район, около посёлка Зарубино, 19 июня 2022, фото О.Н.Васик; 6 – Уссурийский городской округ, окрестности села Утёсное, фото Д.А.Беляева
По данным О.А.Бурковского (Бурковский 2005; Burkovskiy 1997), при строительстве гнёзд ремезы используют растительные волокна (луб), что делает гнёзда прочными, а также пух семян ивы и сложноцветных Compositae, паутину, а в одной постройке обнаружили шерсть. В стенках одного гнезда, осмотренного на полуострове Песчаный 1 июня 2018, помимо растительного пуха и волокон, выщипанных из стеблей тростника, присутствовали также цветки из корзинок одуванчика и тычиночные нити из цветков уссурийской груши Pyrus ussuriensis (Волков-ская-Курдюкова, Курдюков 2019). По собранным нами сведениям, материал, используемый китайскими ремезами для сооружения гнёзд, имеет почти исключительно растительное происхождение, но в 1 гнезде среди найденных на полуострове Песчаный в значительном количестве присутствовали волокна синтепона. Размеры гнёзд приведены в таблице 4.
Таблица 4. Размеры (мм) гнёзд китайского ремеза Remiz consobrinus в Приморском крае
|
n |
Диаметр гнезда |
Высота гнезда |
Диаметр лотка |
Длина входной трубки (по нижнему краю) |
Диаметр летка |
Источник информации |
|||||
|
Lim |
Среднее |
Lim |
Среднее |
Lim |
Среднее |
Lim |
Среднее |
Lim |
Среднее |
||
|
9 |
70-92 |
80.2 |
120-154 |
134.2 |
56-61 |
58.5 |
12-36 |
21.3 |
21-27 |
23.9 |
Наши данные |
|
3 |
87-100 |
91.3 |
140-147 |
142.7 |
64-73 |
69.7 |
9-29 |
18.0 |
– |
– |
Burkovskiy 1997 |
|
12 |
70-100 |
83.0 |
120-154 |
136.3 |
56-73 |
61.3 |
9-36 |
20.5 |
21-27 |
23.9* |
Всего |
* – рассчитано по 9 промерам.
Массовое индивидуальное мечение позволило выявить, что для этого вида характерна последовательная полигиния, а моногамные отношения хотя и присутствуют, но представлены слабо (Бурковский и др. 2014). После завершения строительства первых гнёзд самцы оставляют самок и приступают к строительству следующих гнёзд сезона (от 1 до 3). Некоторые самцы последовательно перемещаются по территории, строя новые гнёзда, а другие сооружают одно гнездо и держатся около него до затухания гнездостроительной активности; отмечены случаи расположения двух так называемых «холостых» гнёзд одним самцом на отдельном дереве. Довольно часто они строят новое гнездо вблизи постройки «чужой» оставленной самки, иногда – на том же дереве, а в 2014 году было отмечено три случая занятия самцами чужих сооружений. Холостые самцы строят гнёзда не полностью (только до промежуточной стадии, когда они приобретают форму корзинки), а в дальнейшем при условии образовании пары такие постройки доводятся до окончательного состояния партнёрами уже совместно (Бурковский и др. 2014).
Многие недостроенные гнёзда холостых самцов остаются в таком же виде (корзиночки), сохраняясь в течение длительного времени, например зимой и к началу нового гнездового периода (рис. 18). Поскольку старые постройки (как полностью построенные, так и недостроенные)
инициируют пролётных птиц к размножению поблизости, это способствует долгосрочному сохранению поселения и в конечном итоге – к поддержанию мозаичной структуры популяции.

Рис. 18. Старые недостроенные гнёзда китайских ремезов Remiz consobrinus .
1 – восточное побережье озера Ханка, 29 октября 2008; 2 – там же, 27 марта 2010; 3 – Михайловский район, окрестности посёлка Новошахтинский, 4 декабря 2024; 4 – восточное побережье озера Ханка, 27 мая 2012. Фото Д.В.Коробова
Наименьшее расстояние между жилыми гнёздами разных самок составляет около 90 м. Создаётся впечатление, что значительная часть известных гнездовых группировок состоит из отдельных рыхлых поселений, которых условно можно назвать колониями, а не хаотично размещается в подходящих местообитаниях. Птицы таких конгломераций регулярно обмениваются звуковыми сигналами, причём издаваемый ими тонкий пронзительный свист слышен человеком на расстоянии до 400 м (Бурковский и др. 2014).
Сроки откладки яиц растянуты со второй половины мая по июль (табл. 4). В полных кладках китайских ремезов мы находили от 5 до 8 яиц, чаще всего их было 7 (рис. 19-21). Средняя величина кладки соста- вила 6.87 яйца (n = 23). По другим данным, кладки содержат 6-7 яиц (Бурковский 2005), а в одном из гнёзд, обнаруженных в Хасанском районе на берегу озере Птичье, было 10 яиц (сообщение В.А.Нечаева).
Линейные размеры и индекс удлинённости яиц китайских ремезов приведены в таблице 5.
Вес свежих и слабо насиженных яиц, по нашим данным, находился в пределах от 0.8 до 1.2, в среднем составляя 0.99 г ( n = 142). Объём яиц, рассчитанный по формуле: V = 0.51 LB 2, где L – длина яйца, B – максимальный диаметр (Hoyt 1979), колебался от 0.8 до 1.1, в среднем составляя 0.97 см3 ( n = 170).
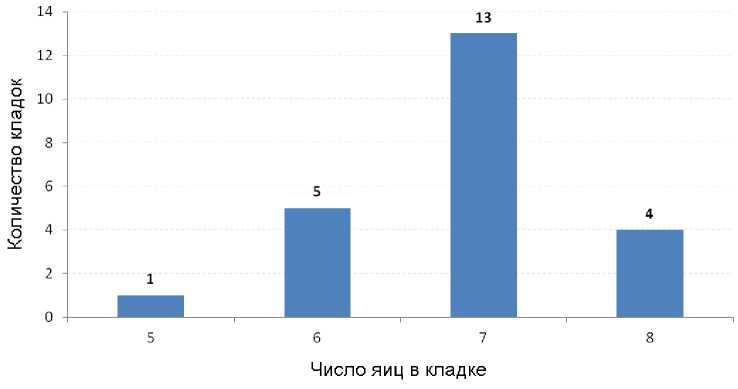
Рис. 19. Число яиц в полных кладках китайского ремеза Remiz consobrinus в Приморском крае (наши данные за 2011-2023 годы)

Рис. 20. Полные кладки китайского ремеза Remiz consobrinus , выложенные на субстрате внутренней части гнезда. 1 – восточное побережье озера Ханка к северу от устья реки Спасовка, 8 июня 2011, фото Д.В.Коробова; 2 – Хасанский район, окрестности посёлка Хасан, 10 июня 2023, фото А.П.Ходакова

Рис. 21. Полные кладки китайского ремеза Remiz consobrinus из Приморского края. Студийная съёмка кладок из оологической коллекции В.Н.Сотникова (город Киров)
Яйца в гнезде бывают погружены в толстый слой рыхлой выстилки лотка, так что сверху они незаметны. Обычно самки закрывают кладку в случае, когда она ещё неполная, а в период насиживания её открывают. Однако при осмотре гнёзд, осуществлённом в зимнее время, в толще такой выстилки мы неоднократно находили испорченные яйца (рис. 22). Нельзя исключить того, что часть закрытых яиц самки попросту не находят, оставляя их в зарытом в выстилку состоянии.
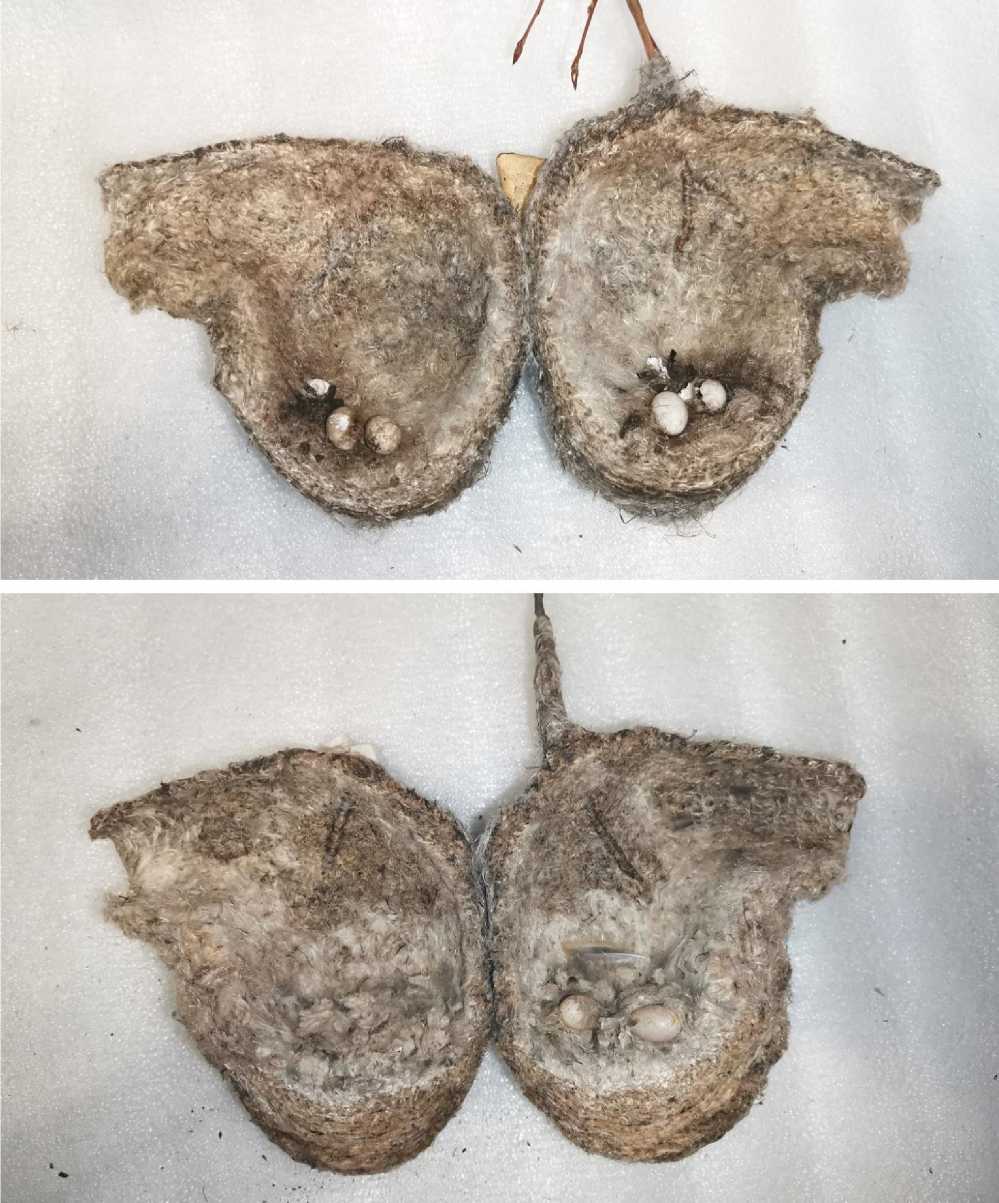
Рис. 22. Вскрытые гнёзда китайского ремеза Remiz consobrinus , содержащие оставшиеся в них яйца. Студийная съёмка гнёзд, собранных на полуострове Песчаный (Амурский залив) 3 января 2026
Насиживанием и последующим выкармливанием птенцов занимается исключительно самка (рис. 23), хотя существует указание на то, что в одном случае, зарегистрированном 5 августа 1996, слётков кормил самец (Burkovskiy 1997). Инкубация яиц длится 11-14 сут, а птенцы покидают гнездо в возрасте до 20 сут, после чего самка в течение некоторого времени ещё продолжает их докармливать.
Таблица 5. Линейные размеры и индекс удлинённости яиц китайского ремеза Remiz consobrinus в Приморском крае
|
n |
Длина (L) , мм |
Максимальный диаметр (B) , мм |
Индекс удлинённости, %* |
Источник информации |
|||
|
Пределы |
Среднее |
Пределы |
Среднее |
Пределы |
Среднее |
||
|
170 |
14.2-18.0 |
16.17 |
10.1-11.4 |
10.81 |
60.2-74.7 |
66.9 |
Наши данные** |
|
12 |
14.4-16.6 |
15.90 |
10.2-10.9 |
10.50 |
– |
– |
Burkovskiy 1997 |
|
182 |
14.2-18.0 |
16.15 |
10.2-11.4 |
10.79 |
60.2-74.7 |
66.9*** |
Всего |
* – рассчитан по формуле: (B / L) × 100% (Романов, Романова 1959); ** – включены данные, опубликованные ранее (Глущенко и др. 2006; Сотников 2023); ***рассчитано по 170 промерам.

Рис. 23. Самка китайского ремеза Remiz consobrinus у гнезда с птенцами. Приханкайская низменность. 6 августа 2012. Фото Д.В.Коробова
У китайского ремеза птенцы покидают гнездо полностью оперёнными (рис. 24) и умеющими хорошо летать. Наиболее поздних гнездовых птенцов мы наблюдали на полуострове Песчаный (Амурский залив) 24 августа 2023 (рис. 25). Слётков регистрировали в течение июля и августа (табл. 4; рис. 26). В выводке насчитывали 4-6 слётков (Бурковский 2005). Существует указание на то, что на ночёвку самки могут приводить слётков в гнездо (Бурковский 2014).
За гнездовой сезон большинство самок успевает вывести только один выводок, но установлено, что в ряде случаев у тех птиц, выводки которых становятся самостоятельными одними из первых, бывает два полных цикла размножения (Бурковский и др. 2014).

Рис. 24. Птенцы китайского ремеза Remiz consobrinus , покидающие гнездо. Приханкайская низменность, 6 августа 2012. Фото Д.В.Коробова

Рис. 25. Самые поздние гнездовые птенцы китайского ремеза Remiz consobrinus . Западное побережье Амурского залива, полуостров Песчаный. 24 августа 2023. Фото Д.В.Коробова
Послегнездовые перемещения, осенний пролёт и зимовка. С июля начинаются кочёвки групп молодых ремезов, переходящие в осенний пролёт, характер которого в условиях Приморского края практиче- ски не изучен. Выяснено, что в ряде случаев тесная связь с территорией колонии у ремезов может сохраняться до начала октября (Бурковский и др. 2014). Вероятно, часть молодых птиц присоединяется к кочующим синичьим и смешанным стаям и иногда вместе с ними они перемещаются довольно далеко от мест гнездования.

Рис. 26. Слётки китайского ремеза Remiz consobrinus . 1 – Хасанский район, окрестности посёлка Хасан, 31 июля 2020, фото А.В.Вялкова; 2 – западное побережье залива Петра Великого, подножье Голубиного утёса, 21 августа 2008, фото Д.В.Коробова

Рис. 27. Самая поздняя из известных осенняя встреча китайского ремеза Remiz consobrinus . Западное побережье Амурского залива, бухта Перевозная, 27 октября 2019. Фото А.П.Рогаля
Двух таких мигрирующих ремезов отловили 13 октября 2013 на юго-востоке края в бухте Петрова Лазовского заповедника (Шохрин 2014). Самая поздняя осенняя встреча датирована 27 октября 2022 (рис. 27). Зимующего самца китайского ремеза наблюдали на крайнем северо-западе Приморского края в окрестностях Лучегорска (Burkovskiy 1997).

Рис. 28. Самка китайского ремеза Remiz consobrinus с кормом для птенцов: 1-4 – с гусеницами; 5-8 – с пауками. Приханкайская низменность. 6 августа 2012. Фото Д.В.Коробова
Питание . В пище взрослых птиц и птенцов доминируют личинки насекомых (Бурковский 2005). Во время наших немногочисленных наблюдений у гнёзд китайских ремезов с птенцами самки приносили им гусениц и пауков (рис. 28).
Судя по пищевым объектам, китайские ремезы в большинстве случаев собирают добычу с поверхности листьев и ветвей, но могут извлекать её и из толщи стеблей растений, например, тростника (рис. 29). Отмечено также кормление птиц на цветках ивы, при этом, судя по имеющимся фотографиям, в таком случае они могли поедать не только насекомых, но и фрагменты цветков (рис. 30).

Рис. 29. Самка китайского ремеза Remiz consobrinus , извлекающая корм из стебля тростника. Западное побережье залива Петра Великого у мыса Островок Фальшивый. 20 мая 2014. Фото Д.В.Коробова

Рис. 30. Самец китайского ремеза Remiz consobrinus , поедающий цветки ивы. Устье реки Раздольная, озеро Утиное. 29 апреля 2018. Фото А.П.Рогаля
В окрестностях Лучегорска в феврале 1997 года в течение нескольких дней самец ремеза время от времени кормился в зарослях рогоза Typha sp. Птица тянула за семенные головки этого растения так же, как это делали замеченные здесь же болотные гаички Poecile palustris . При внимательном рассмотрении семенных головок обнаружили от 1 до 5 личинок насекомых длиной 3-7 мм (Burkovskiy 1997).
Неблагоприятные факторы, враги, гибель . Имеется информация о том, что часть гнёзд ремезов погибает от намокания после продолжительных моросящих дождей и от хищничества врановых птиц (Бур-ковский 2005). Прямых наблюдений разорения гнёзд китайских ремезов хищниками у нас нет, но гнёзда, явно разрушенные ими, мы многократно находили (рис. 31, 32).

Рис. 31. Разорённое гнездо китайского ремеза Remiz consobrinus: 1 – общий вид;
2 – крупный план. Долина реки Раздольная, окрестности села Утёсное. Уссурийский городской округ. 24 декабря 2025. Фото Д.А.Беляева
В разных случаях гнёзда были разорваны с разных сторон: сбоку (рис. 32. 1) , сзади (рис. 32. 3) либо спереди в области летка (рис. 31, 32. 2) . Нельзя исключить того, что отмеченные нами случаи исчезновения поселений китайских ремезов могут быть вызваны регулярным разорением их гнёзд отдельными особями хищников, получивших успешный опыт такого сравнительно простого способа добычи корма.
Пустующие гнёзда ремезов в холодное время года могут использовать мелкие птицы в качестве мест для ночёвки. Судя по скоплению большого количества помёта, явно оставленного не птенцами ремеза, одна из таких особей использовала гнездо, повреждённое неизвестным хищником (рис. 33).

Рис. 32. Разорённые гнёзда китайских ремезов Remiz consobrinus .
Амурский залив, полуостров Песчаный. 3 января 2026. Фото Д.В.Коробова
Условия гнездования китайских ремезов в их типичных биотопах могут ухудшать весьма частые травяные пожары, а также уничтожение древесной растительности (Бурковский 2005).
Исследование выполнено в рамках госзаданий Минобрнауки РФ № 125021302113-3 и № 124012400285-7. За помощь в работе и предоставленные сведения авторы выражают искреннюю благодарность С.Ф.Акулинкину (Киров), Н.Н.Балацкому (Новосибирск), Г.Н.Ба-чурину (Ирбит), О.Н.Васик (Владивосток), В.Д.Ващенко (Кировский район), А.В.Вялкову (Владивосток), О.М.Косолапову (Владивосток), Д.Н.Кочеткову (Читинская область), И.А.Малыкиной (Владивосток), И.В.Масловой (Владивосток), В.М.Матюшину (Владивосток), И.Н.Отставных (Ханкайский р-н), А.П.Рогалю (Владивосток), И.И.Уколову (Москва).

Рис. 33. Разорённое гнездо китайского ремеза Remiz consobrinus , использованное мелкой птицей для ночёвки в холодное время года. Амурский залив, полуостров Песчаный. 3 января 2026. Фото Д.В.Коробова
