Градиентный анализ флористического состава и экологической структуры травяного яруса в пойменных лесах

Автор: Браславская Татьяна Юрьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В статье рассмотрено варьирование видового состава и эколого-ценотической структуры травяного покрова в старовозрастных пойменных лесах в связи с режимом паводкового заливания и освещенности.
Пойменные леса, экотоны, режим паводкового заливания и освещенности, эколого-ценотические группы видов, экологические стратегии видов
Короткий адрес: https://sciup.org/148200786
IDR: 148200786 | УДК: 635.054:556.5
Gradient analysis of herb-layer species composition and functional structure in afforested floodplain
The study deals with spatial variability of herb layer in floodplain conifer-broadleaved old forests (region of Middle Volga). It is shown that light conditions, inundation and species interactions all together influence distribution of herb species realizing whatever ecological strategy.
Текст научной статьи Градиентный анализ флористического состава и экологической структуры травяного яруса в пойменных лесах
шага». Они подбирались таким образом, чтобы в древостое 1) отсутствовали или участвовали в минимальном количестве мелколиственные виды (березы, осина), 2) присутствовали деревья широколиственных или темнохвойных видов возраста 100-140 лет (информация бралась из материалов лесоустройства). В статье использованы материалы с 3-х ключевых участков в центральнопойменной зоне. На этих участках были заложены временные пробные площади размером 100х100 м, в пределах которых было закартировано на миллиметровой бумаге в масштабе 1:500 расположение грив и межгривных понижений пойменного мезорельефа, а также лесных и оконных парцелл. При картировании различались следующие элементы мезорельефа: верхушки грив, склоны грив, днища понижений. Для измерения перепадов высотных отметок использовался гидроуровень, применяемый в строительных работах [7]. Границы между лесными (с сомкнутостью полога древостоя более 30 %) и оконными парцеллами проводились по вертикальным проекциям крон деревьев. Среди оконных парцелл различались мелкие окна (имеющие хотя бы один линейный размер менее 20 м, т.е. меньше высоты примыкающего древостоя) и крупные окна (у которых все линейные размеры более 20 м).
Описания травяного яруса были выполнены в пределах каждой пробной площади на 100-125 площадках размером 2х2 м. Места для их заложения выбирались с учетом приуроченности лесных и оконных парцелл к различным элементам мезорельефа (пропорционально территории, занятой каждым из выявленных сочетаний в пределах пробной площади). Расположение каждой площадки было нанесено на общий план пробной площади и, в соответствии с этим, указано, на каком элементе мезорельефа и в какой парцелле она заложена. Для большинства площадок были также проведены измерения относительных высотных отметок, на основании чего была сделана оценка продолжительности их заливания в период весеннего половодья (см. ниже). Для каждой площадки определялась сомкнутость полога древостоя (яруса А) и полога подлеска (яруса В) непосредственно над ней, общее проективное покрытие травяного покрова
(яруса С) и мохового покрова (яруса D), наличие воды на поверхности и ее покрытие, составлялся полный флористический список сосудистых растений в составе яруса С с указанием баллов проективного покрытия по шкале Браун-Бланке. Видовой состав напочвенных и эпиксильных мхов не исследовался (в исследуемых лесах общее проективное покрытие этих синузий обычно составляет 1-2 %, так что они не играют эдификаторной роли).
В ходе измерения перепадов высотных отметок в мезорельефе местными реперами (точками отсчета) служили деревья, помеченные цветными полиэтиленовыми лентами (на каждой пробной площади – по 8-9 равномерно распределенных реперов). В период максимума весеннего половодья 2003г., когда вся покрывавшая пойму вода имела единую поверхность, возле каждого реперного дерева была измерена глубина воды (22 апреля в 9.00-11.00). В это же время сотрудниками заповедника «Большая Кокшага» было проведено плановое измерение уровня воды на постоянном водомерном посту (23км от пробных площадей). Благодаря этому, в соответствии с рекомендованной методикой [2], можно было приравнять уровень воды на каждом репере к уровню на водомерном посту и пересчитать высотные отметки поверхности поймы, измеренные относительно разных реперов, в единой системе отсчета относительно условной нулевой отметки водомерного поста.
С 2001г. регистрация уровней воды на водомерном посту проводится сотрудниками заповедника ежедневно (кроме периода ледостава). На основе наблюдений 2001-2010гг. была рассчитана продолжительность заливания различных высотных отметок поймы (в диапазоне 10-480см с шагом 10см) в каждом году и теоретическая периодичность (т.н. обеспеченность) [8] каждой вычисленной продолжительности заливания.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В ходе анализа описаний была выполнена их ординация по методу бестрендового анализа соответствий (DCA, в пакете PCOrd v.4). Затем была вычислена корреляция (ранговый коэффициент Спирмена – RSp) между построенными ординаци-онными осями и 1) высотными отметками площадок, 2) суммарными значениями сомкнутости ярусов А и В над площадками, а также корреляция этих показателей между собой. Для каждого описания определено относительное участие (доля в общей сумме баллов покрытия) видов различных эколого-ценотических групп (ЭЦГ) и различных экологических стратегий (распределение видов по ЭЦГ принято по Л.Б. Заугольновой [5]; информация об экологических стратегиях травянистых видов взята из работ А.А. Агафоновой с соавторами [1], Л.А. Жуковой [4], Е.П. Сарычевой [11], О.В. Смирновой [12]). Рассмотрено изменение этих показателей вдоль градиентов относительных высотных отметок и флористического состава, для чего проведено сглаживание методом взвешенных на абсциссу наименьших квадратных отклонений от линейной регрессии (DWLS; в пакете Statistica 6.0).
DCA-ординация описаний выявила в травяном ярусе практически одномерный градиент изменения флористического состава (рис. 1). На одном конце этого градиента сосредоточены описания с верхушек грив и верхних частей склонов, где доминирует обычно Matteuccia struthiopteris (L.) Tod. (ЭЦГ высокотравья), а субдоминантами выступают различные виды неморальной ЭЦГ – Mercurialis perennis L., Aegopodium podagraria L., Lathyrus vernus (L.) Bernh., а также Oxalis acetosella L. (бореальная ЭЦГ). На противоположном конце построенного градиента сосредоточены описания преимущественно с пониженных участков, где доминируют и субдоминируют виды водно-болотной ЭЦГ – Carex vesicaria L., Galium palustre L., Symphytum officinale L., Caltha palustris L., или же виды высокотравья – Filipendula ulmaria (L.) Maxim., Stachys palustris L., Lythrum salicaria L. В средней части градиента оказались описания (с различных местоположений), в которых доминируют или со-доминируют друг с другом виды вы-сокотравья – часто Urtica dioica L. и Rubus caesius L., а также перечисленные выше.
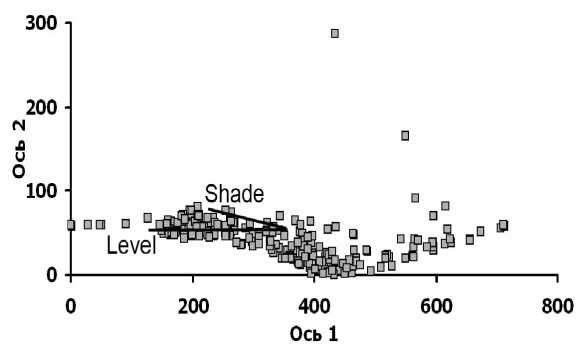
Рис. 1. Ординация описаний травяного яруса в осях флористического сходства и ее сопоставление с относительными высотными отметками и затенением площадок (отображены в виде векторов, направленных в сторону возрастания каждого из показателей). Условные обозначения: Level – относительные высотные отметки площадок, Shade – суммарные значения сомкнутости ярусов А и В над площадками.
С выявленным градиентом флористического состава (координатами описаний на 1-й ординацион-ной оси) наиболее тесно связаны высотные отметки площадок (RSp = -0,722, p<0,01), т.е. показатели режима заливания и увлажнения почвы. Суммарные значения сомкнутости ярусов А и В, т.е. показатели режима затенения напочвенного покрова, связаны с этим градиентом в меньшей степени (RSp = -0,517, p<0,01). Связь между высотными отметками площадок и суммарными значениями сомкнутости выражена еще слабее, но значима (RSp = -0,433, p<0,01). На местности эта связь выражается в том, что верхушки грив и верхние части склонов обычно заняты лесными парцеллами, и окон там мало, а к днищам межгривных понижений обычно приурочены парцеллы крупных или мелких окон, вследствие чего на нижних частях склонов выражены опушки разной ширины и с разной разреженностью древесных растений. Таким образом, в исследованных пойменных лесах заливание и затенение влияют на травяную синузию, взаимодействуя друг с другом, и градиент изменений флористического состава яруса С отражает комплексную реакцию на это влияние.
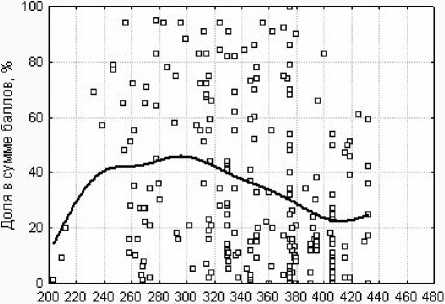
Изменчивость и функциональные связи экологической структуры травяного яруса в исследованных пойменных лесах можно проиллюстрировать относительным участием видов с разными экологическими стратегиями. На диаграммах рассеяния (рис. 2, 1 ) представлено соотношение градиента высотных отметок и общего (суммарного) относительного участия в травяном покрове всех видов, которые в той или иной степени проявляют реак-
Относительные высотные отметки, см
Рис. 2. Относительное участие в составе травяного яруса (доля в общей сумме баллов покрытия) видов, проявляющих реактивные свойства. Линией показан сглаженный тренд распределения вдоль градиента: 1 - высотных отметок; 2 - ординации описаний по флористическому сходству (ось 1).
При сопоставлении относительного участия видов, проявляющих разные стратегии, и положения описаний в градиенте флористического состава гораздо явственнее выражен колоколообразный характер диаграмм рассеяния: на крайних отрезках градиента обычно мало участие видов с реактивными свойствами (рис. 2, 2 ) и велико участие конкурентных и стресс-толерантных видов, а в средней части градиента - наоборот. Вклад видов различных ЭЦГ в это распределение - точно такой же, как описано выше. Видимо, выявленный ординацион-ный градиент флористического состава отражает не только разнообразие экотопов и несходство видов по экологическим амплитудам, но в значительной степени также и результат «раздела территории» в ходе взаимодействий между видами с перекрывающимися широкими амплитудами и различными стратегиями, вследствие чего на этом градиенте более четко, чем на высотном, дифференцирована функциональная структура растительности.
тивные [4, 12] свойства. Прослеживается тенденция, хотя и слабая, что на обоих противоположных концах этого градиента - на высотных отметках менее 240 см и более 300 см - чаще наблюдается малое участие этих видов в травяном ярусе, а на промежуточном высотном уровне 240-300 см - чаще большое, причем оно создается здесь видами высокотравья, черноольховой, водно-болотной и неморальной ЭЦГ. Соответственно, противоположным образом распределено общее относительное участие видов, проявляющих только конкурентные и стресс-толерантные свойства, - часто оно мало на промежуточном отрезке градиента высот, а на краях градиента - чаще большое. В его возрастание на краях градиента существенный вклад обычно вносят: на пониженных участках -виды высокотравья и водно-болотной ЭЦГ, а на повышенных - виды высокотравья, неморальной и бореальной ЭЦГ.
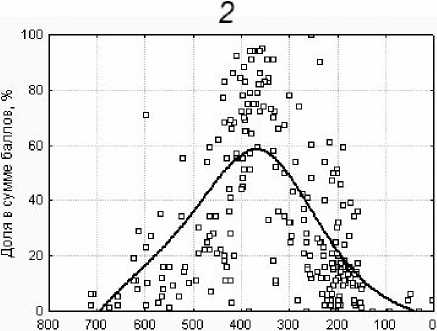
Ось 1
Результат проведенного градиентного анализа демонстрирует, что экотоны в пойменных лесах характеризуются не просто промежуточным экологическим режимом и флористическим составом, а еще и очень мозаичной пространственной структурой травяного яруса. Для формирования этой мозаичности, видимо, особенно благоприятны условия таких пойм, как исследуемая, где в настоящее время абиотические и биотические экологические факторы примерно равнозначны по интенсивности воздействия на лесной покров. Градиент изменений флористического состава травяного яруса отражает его комплексную реакцию на взаимосвязанное влияние паводкового заливания и затенения.
Участие в полевых исследованиях принимали студенты и аспиранты МПГУ и МГУ им. М.В. Ломоносова: Т.С. Проказина, Е.Ю. Бакун, И.В. Соколов, М.В. Гаврилова, О.В. Воронцова, С.Е. Петрова. В период полевых исследований работа была частично профинансирована РФФИ (03-05-6423 8). В период обработки данных работа была поддер- жана Программой фундаментальных исследований Президиума РАН «Биоразнообразие и динамика генофондов».
Список литературы Градиентный анализ флористического состава и экологической структуры травяного яруса в пойменных лесах
- Агафонова А.А. Краткая характеристика популяционной биологии константных видов травяного покрова/Агафонова А.А., Шорина Н.И., Смирнова О.В., Жукова Л.А. и др.//Восточноевропейские леса: история в голоцене и современность. М.: Наука, 2004. Кн. 1. С.224-256.
- Алексеенко Л. Н. К методике использования гидрологических материалов для характеристики условий развития пойменной растительности//Вестн. ЛГУ. Геология. География. 1973. № 18. С.111-122.
- Еленевский Р.А. Вопросы изучения и освоения пойм. М.: Изд. ВАСХНИЛ, 1936. 100 с.
- Жукова Л.А. Популяционная жизнь луговых трав. Йошкар-Ола: Марийский гос. ун-т, 1995. 223 с.
- Заугольнова Л.Б. Южная тайга и подтаежная зона [Электронный ресурс]//Ценофонд лесов Европейской России: сайт. URL: http://mfd.cepl.rssi.ru/flora/.
- Исаев А.В. Формирование почвенного и растительного покрова в поймах речных долин Марийского Полесья (на примере территории заповедника «Большая Кокшага»). Йошкар-Ола: Марийский гос. техн. ун-т, 2008. 240 с.
- Кавунец Д.Н. Гидростатическое нивелирование на строительной площадке. М.: Геодезиздат, 1961. 118 с.
- Клибашев К. П. Гидрологические расчеты/Клибашев К. П., Горошков И. Ф. [под ред. проф. Чеботарева А. И.]. 2-е изд. Л.: Гидрометеоиздат, 1970. 460 с.
- Миркин Б.М. Закономерности развития растительности речных пойм/АН СССР, Башкирский филиал. М.: Наука, 1974. 172 с.
- Раменский Л.Г. Введение в комплексное почвенногеоботаническое изучение земель. М.: Огиз, Сельхозгиз, 1938. 620 с.
- Сарычева Е.П. Структурное и видовое разнообразие черноольховых лесов центра Европейской России (на примере заповедников «Брянский лес» и «Воронинский»). Дисс. на соиск. уч. степ. канд. биол. наук/Пущинский гос. ун-т. М., 2000. 135 с.
- Смирнова О.В. Структура травяного покрова широколиственных лесов. М.: Наука, 1987. 206 с.
- Battaglia L. L., Sharitz R. R. Responses of floodplain forest species to spatially condensed gradients: a test of the flood-shade tolerance tradeoff hypothesis//Oecologia. 2006. V. 147, № 2. P. 108-118.
- Menges E.S., Waller D.M. Plant strategies in relation to elevation and light in floodplain herbs//American Naturalist. 1983. V. 122, № 4. P. 454-473.
- Siebel H.N., Bouwma I.M. The occurrence of herbs and woody juveniles in a hardwood floodplain forest in relation to flooding and light//Journ. Veget. Science. 1998. Vol. 9, № 5. P. 623-630.
