Характеристика ценопопуляций и особенности мезоструктуры листа Orthilia secunda L. на нарушенных промышленностью землях

Автор: Чукина Надежда Владимировна, Глазырина Маргарита Александровна, Лукина Наталия Валентиновна, Бутырин Кирилл Владимирович, Лихачева Марина Викторовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
В статье представлены результаты изучения возрастной, морфологической и пространственной структур ценопопуляций Orthilia secunda L., мезоструктуры фотосинтетического аппарата и особенностей строения микоризы данного вида, в растительных сообществах на разных техногенных субстратах. Показано, что на нарушенных промышленностью землях, у Orthilia secunda L., происходит изменение мезоструктурных параметров (уменьшение толщины листа, размеров площади ассимилирующей поверхности клеток и хлоропластов) по сравнению с естественными местообитаниями.
Мезоструктура, ценопопуляция, арбутоидная микориза, промышленные отвалы
Короткий адрес: https://sciup.org/148202032
IDR: 148202032 | УДК: 581.55:581.552:581.557.24:502
The characteristics of cenopopulations and peculiarities of leaf mesostructure Orthilia secunda L. on the disturbed industrial lands
The results of study of age, morphological and spatial structure of cenopopulation Orthilia secunda L., mezostructure of the photosynthetic apparatus and type of mycorrhizae in plant communities in various technogenic substrates are presented in this paper. It is shown that in disturbed industrial lands, Orthilia secunda L. changes its morphology and its mezostructure parameters (decreasing in leaf thickness, size of chloroplasts, internal assimilation surface mezophyll and chloroplasts) compared with natural habitats.
Текст научной статьи Характеристика ценопопуляций и особенности мезоструктуры листа Orthilia secunda L. на нарушенных промышленностью землях
В связи с увеличивающимися темпами развития промышленности, усиливается влияние человека на структуру и функции биосферы. Результатом такого воздействия является возникновение техногенных ландшафтов. Среди техногенных ландшафтов особое место по своему отрицательному воздействию на естественные природные комплексы занимают промышленные отвалы. Восстановление растительности на них происходит крайне медленно. В настоящее время проводится изучение структуры, динамики, флористического состава растительных сообществ, изучение ценопопуляций видов, произрастающих на нарушенных промышленностью землях. Вместе с тем, работ по изучению эколого-физиологических механизмов адаптаций растений к техногенным субстратам крайне мало.
Известно, что основным в определении роста и продуктивности растений, является фотосинтетический аппарат, он оказывается наиболее чувствительным к действию неблагоприятных факторов среды, что проявляется в нарушении многих параметров его функционирования [1].
В связи с этим целью наших исследований было изучение возрастной, морфологической и пространственной структур ценопопуляций (ЦП) Or-thilia secunda L., а также мезоструктуры фотосинтетического аппарата и особенностей строения микоризы данного вида, в растительных сообществах на разных техногенных субстратах.
На промышленных отвалах Среднего Урала (таежная зона, подзона южной тайги), на начальных стадиях формирования лесных фитоценозов начинают поселяться типичные лесные виды, в том числе Orthilia secunda L.
Наши исследования проводились в июле 2012 г. на следующих объектах:
– Лесной фитоценоз с доминированием Pinus sylvestris L. (cop 2 –cop 3 ), Betula pendula Roth, B. pubescens Ehrh., Populus tremula L. (cop 1 ), сформировавшийся на золоотвале Верхнетагильской ГРЭС на рекультивированном участке с полосным нанесением потенциально плодородного грунта (ЦП 1 ). Возраст сообщества около 40 лет, сомкнутость крон деревьев – 0,6–0,8.
– Мелколиственный лес с доминированием Populus tremula (cop 2 ), Betula pendula (cop 2 ) и B. pubescens (cop 1 ), Salix caprea L. (sp), сформировавшийся на золоотвале Верхнетагильской ГРЭС на золе (ЦП 2 ). Возраст сообщества около 30 лет, сомкнутость крон – до 0,7–0,8.
– Лесной фитоценоз с доминированием Pinus sylvestris (cop 2 ), Populus tremula (cop 1 ), Betula pubescens и Salix caprea (sp), сформировавшийся на Галкинском отвале мраморизированного известняка (ЦП 3 ), образованного пустыми породами вскрыши и отходами мелочи с дробильнообогатительной фабрики известнякового карьера. Основная масса флюсовых известняков содержит CaO – 50–52 %. Возраст фитоценоза около 50 лет, сомкнутость крон – 0,7–0,8.
– Лесной фитоценоз на дамбе гидроотвала Шу-ралино-Ягодного месторождения россыпного золота (ЦП 4 ), сложенной тяжелосуглинистыми среднекаменистыми вскрышными породами. Возраст растительных сообществ составляет около 18 лет. Древесный ярус представлен: Betula pubescens (sol–sp), Salix caprea (sol–sp), S. viminalis L. (sol– sp), Pinus sylvestris (sol), Picea obovata Ledeb. (sol), Betula pendula (un). Сомкнутость крон достигает 0,3–0,5.
Все объекты характеризуются низким содержанием азота и фосфора. Подробная характеристика субстратов представлена в работах [2, 3].
В качестве контроля (ЦП 5 ) были отобраны образцы растений в сосняке-черничнике в районе биологической станции УрФУ (таежная зона, подзона южной тайги).
При ценопопуляционных исследованиях Orthi-lia secunda за единицу возрастной, морфологической и пространственной структуры ЦП принимали вегетативно размножающуюся особь – рамет [4, 5]. Для исследований мезоструктурных характеристик фотосинтетического аппарата генеративных особей данного вида, отбирали с 10-15 растений по 5–10 сформированных листьев из каждого местообитания.
Анализ показателей мезоструктуры листьев проводили на растительном материале, фиксированном в 3,5 % растворе глутарового альдегида в фосфатном буфере (рН=7,0) согласно методике А. Т. Мокроносова, Р. А. Борзенковой [6]. Поперечные срезы листьев получали с использованием замораживающего микротома МЗ-2 (Россия). Определение количества клеток в единице площади листа проводили после мацерации тканей в 20% растворе КОН при нагревании с помощью счетной камеры Горяева. Измерения толщины листа, размеров клеток мезофилла и хлоропластов проводили с помощью специализированного комплекса для анализа мезоструктуры листьев Simagis Mesoplant (ООО «СИАМС», Россия), используя световой микроскоп Meiji MT 4300L («Meiji Techno», Япония). Изучались следующие параметры: толщина листа, число клеток и число хлоропластов на единицу площади, объем клеток и объем хлоропластов, индексы общей ассимилирующей поверхности клеток и хлоропластов (ИМК и ИМХ), индекс проективной поверхности хлоропластов (ИППХ).
Для исследования микоризы были сделаны поперечные срезы корневых окончаний на замораживающем микротоме, полученные препараты просматривались в поле зрения микроскопа при 120– 200 увеличении [7].
Orthilia secunda L. – представитель подсемейства Pyroloideae (Грушанковые), циркумполярный бореальный вид, широко распространен на Урале, типичный представитель травяно-кустарничкового яруса в лесах различного типа [8]. Хамефит, ползучий зимне-зеленый поликарпик. Мезофит [9]. В естественных местообитаниях грушанковые как правило микотрофны. У Orthilia secunda микориза ар-бутоидного (экто-эндотрофного) типа, характеризующаяся наличием наружного грибного чехла и однослойной сети Гартига, внутриклеточных клубков гиф и продуктов их переваривания в увеличенных клетках коры корня [7, 10].
В результате проведенных исследований установлено, что все ЦП Orthilia secunda являются нормальными неполночленными (рис. 1). Все возрастные спектры имеют две вершины. На нарушенных землях возрастные спектры являются левосторонними, с максимумом в группе виргиниль-ных, либо имматурных особей. В естественном лесном сообществе спектр – правосторонний. Доля генеративных особей выше в более «зрелых» лесных фитоценозах (ЦП 1 – 25,0 %, ЦП 2 – 13,7 %, ЦП 3 – 11,3 %, ЦП 4 – 12,1 %, ЦП 5 – 53,6 %).
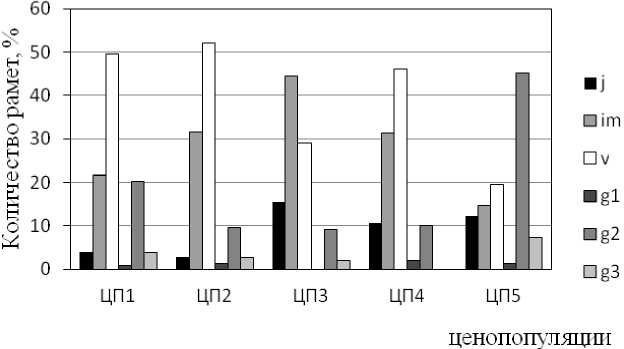
Рис. 1 . Возрастные спектры ценопопуляций Orthilia secunda L.
Тип пространственного распределения всех исследованных ЦП 1-5 Orthilia secunda по отношению дисперсии (S2) к среднему числу особей на площадке является групповым (S2/m=3,22; 2,04; 2,50; 19,73 и 2,13 соответственно).
При исследовании биометрических показателей генеративных особей Orthilia secunda , произрастающих на техногенных субстратах и в контроле, достоверных отличий не выявлено (табл. 1).
Таблица 1 . Некоторые биометрические показатели генеративных особей Orthilia secunda L.
|
Признаки |
ЦП |
N |
Хср.±m |
lim |
Σ |
Cv |
|
Площадь листовой пластинки генеративной особи, см2 |
1 |
23 |
26,6±2,9 |
11,5–69,5 |
13,9384 |
52,4 |
|
2 |
8 |
35,4±4,0 |
14,8–51,5 |
11,4000 |
32,2 |
|
|
3 |
23 |
26,8±2,3 |
11,2–58,0 |
11,1357 |
41,6 |
|
|
4 |
18 |
28,4±3,3 |
6,5–65,5 |
14,1672 |
49,9 |
|
|
5 |
33 |
25,1±1,9 |
6,5–52,8 |
10,9794 |
43,7 |
|
|
Высота генеративного побега, см |
1 |
32 |
16,5±0,41 |
13,0–23,0 |
2,3262 |
14,1 |
|
2 |
10 |
17,6±0,38 |
15,0–19,5 |
1,1891 |
6,8 |
|
|
3 |
25 |
15,7±0,33 |
12,5–19,0 |
1,6503 |
10,5 |
|
|
4 |
14 |
15,5±0,59 |
12,0–19,5 |
2,2160 |
14,3 |
|
|
5 |
49 |
16,7±0,35 |
11,0–22,0 |
2,4213 |
14,5 |
|
|
Масса генеративной особи, г |
1 |
32 |
0,31±0,03 |
0,09–0,81 |
0,1615 |
52,1 |
|
2 |
10 |
0,40±0,04 |
0,22–0,54 |
0,1206 |
30,2 |
|
|
3 |
25 |
0,28±0,02 |
0,10–0,52 |
0,0842 |
30,1 |
|
|
4 |
14 |
0,34±0,05 |
0,17–0,88 |
0,1980 |
58,2 |
|
|
5 |
49 |
0,38±0,02 |
0,17–0,75 |
0,1342 |
35,3 |
Известно, что структурно-функциональные параметры мезофилла листа, такие как размеры клеток и хлоропластов, количество клеток мезофилла и хлоропластов в единице площади могут существенно изменяться под действием различных экологических факторов. Тип и диапазон структурных изменений фотосинтетического аппарата под влиянием факторов среды зависят от функциональных особенностей вида, что в свою очередь позволяет оценить состояние, экологические и функциональные свойства вида, популяции, а также прогнозировать реакцию растений на изменение условий среды. Ряд расчетных параметров мезоструктуры листа, таких как площадь ассимиляционной поверхности клеток и хлоропластов, положительно коррелируют с максимальной интенсивностью фотосинтеза [11, 12].
Результаты исследований структуры фотосинтетического аппарата листьев Orthilia secunda показали, что растения с промышленных отвалов (ЦП 14 ), в сравнении с особями из контрольного местообитания (ЦП 5 ), отличались достоверно меньшими размерами толщины листовой пластинки, что в свою очередь связано со снижением толщины слоя мезофильных клеток. При этом отмечено значительное уменьшение количества клеток мезофилла в единице площади листа у растений с промышленных отвалов. Однако статистически достоверных различий в размерах клеток ассимиляционной ткани листа нами обнаружено не было (табл. 2).
Таблица 2 . Параметры мезоструктуры листа растений Orthilia secunda L., произрастающих на различных техногенных субстратах
|
Параметры |
ЦП 1 |
ЦП 2 |
ЦП 3 |
ЦП 4 |
ЦП 5 |
|
|
Толщина листа, мкм |
Хср.±m |
143,0±3,5 |
142,1±3,9 |
125,9±2,9 |
137,2±1,6 |
161,1±3,0 |
|
lim |
122,7–167,7 |
116,8–162,1 |
105,7–147,9 |
125,3–147,0 |
134,0–184,0 |
|
|
Число клеток, тыс./см2 |
Хср.±m |
295,4±10,5 |
342,8±12,4 |
262,3±7,3 |
269,3±6,3 |
353,4±17,4 |
|
lim |
226,5–382,2 |
245,5–418,9 |
198,2–316,1 |
219,4–322,0 |
198,1–491,7 |
|
|
Число хл., млн./см2 |
Хср.±m |
8,9±0,3 |
12,0±0,4 |
7,6±0,2 |
11,8±0,3 |
11,7±0,6 |
|
lim |
6,8–11,5 |
8,6–14,7 |
5,7–9,2 |
9,7–14,2 |
6,5–16,2 |
|
|
Объем кле- 3 ток, тыс. мкм |
Хср.±m |
31,0±2,0 |
36,8±2,9 |
26,4±1,3 |
23,0±1,3 |
27,2±1,7 |
|
lim |
9,5–56,3 |
17,1–72,4 |
15,4–42,8 |
7,4–37,0 |
14,3–52,2 |
|
|
Объем хлп., мкм3 |
Хср.±m |
55,7±1,9 |
54,3±1,6 |
44,9±1,2 |
44,4±1,2 |
64,8±1,6 |
|
lim |
40,1–79,8 |
41,1–73,3 |
28,3–59,3 |
32,7–57,1 |
50,9–89,4 |
|
|
ИМК, см2/см2 |
Хср.±m |
16,6±0,6 |
20,4±0,7 |
12,6±0,4 |
11,6±0,3 |
17,3±0,9 |
|
lim |
12,8–21,5 |
14,6–24,9 |
9,5–15,2 |
9,4–13,8 |
9,7–24,0 |
|
|
ИМХ, см2/см3 |
Хср.±m |
4,9±0,2 |
6,5±0,2 |
3,3±0,1 |
5,2±0,1 |
7,6±0,4 |
|
lim |
3,7–6,3 |
4,6–7,9 |
2,5–4,0 |
4,2–6,2 |
4,2–10,5 |
|
|
ИППХ, см2/см2 |
Хср.±m |
1,3±0,0 |
1,6±0,1 |
0,8±0,0 |
1,3±0,0 |
1,9±0,1 |
|
lim |
1,0–1,6 |
1,2–2,0 |
0,6–1,0 |
1,1–1,6 |
1,0–2,6 |
|
При сравнении параметров пластид (площади поверхности, площади проекции и объемов хлоропластов) установлено, что особи Orthilia secunda, произрастающие на техногенных объектах, характеризовались достоверно меньшими размерами хлоропластов. В среднем величина объема хлоро- пластов у растений из данных местообитаний составляла 50 мкм3, что в 1,3 раза меньше, чем у растений из естественных местообитаний (ЦП5). Эти различия в итоге выразились в существенном уменьшении (более чем в 1,4 раза) таких важных физиологических параметров как индексы общей ассимилирующей поверхности клеток и хлоропластов (ИМК и ИМХ). Данные интегральные показатели отражают фотосинтетическую функцию, связанную со скоростью и эффективностью ассимиляции углекислого газа и положительно коррелируют со скоростью максимального фотосинтеза [12].
Растения с нарушенных местообитаний отличались также достоверно низкими значениями индекса проективной поверхности хлоропластов (ИППХ), являющегося аналогом листового индекса в фитоценозе.
Исследование микоризы Orthilia secunda показало, что на поверхности корешков во всех ценопо-пуляциях, как на промышленных отвалах, так и в контроле, имеются лишь единичные септированные гифы, редко – рыхлые корневые чехлы, иногда встречаются булавовидные корневые окончания. На поперечных срезах корней плотных мицелиальных чехлов не обнаружено, в единичных коровых клетках наблюдаются клубки мицелия и продукты переваривания гриба. У особей Orthilia secunda из естественного местообитания обнаружены единичные фрагменты сети Гартига.
Таким образом, проведенные исследования показали, что у Orthilia secunda, произрастающей на нарушенных промышленностью землях, происходит изменение структурно-функциональных параметров фотосинтетического аппарата. Эти изменения имеют приспособительный характер, т.к. связаны как с особенностью субстрата (бедность и низкая доступность биогенных элементов), так и с особенностью светового и водного режимов. Данные морфологические и анатомические изменения являются, на наш взгляд, одной из форм адаптации данного вида к техногенным субстратам.
-
1. Мокроносов А.Т . Онтогенетический аспект фотосинтеза. М.: Наука, 1981. 196 с.
-
2. Экологические основы и опыт биологической рекультивации нарушенных промышленностью земель / Т.С. Чибрик, Н.В. Лукина, Е.И. Филимонова, М.А. Глазырина. Екатеринбург: Изд-во Урал. ун-та, 2011. 286 с.
-
3. Махонина Г.И . Экологические аспекты
почвообразова- ния в техногенных экосистемах Урала. Екатеринбург: Изд-во Урал. ун-та, 2003. 356 с.
-
4. Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. 217 с.
-
5. Онтогенетический атлас лекарственных растений: Научное издание. Т. IV Йошкар-Ола: МарГУ, 2004. 240 с.
-
6. Фотосинтез: физиология, онтогенез, экология: Коллективная монография / Под ред. Е.С. Роньжиной. Калининград: Изд-во ФГТУ ВПО КГТУ, 2009. 372 с.
-
7. Селиванов И.А . Микосимбиотрофизм, как форма кон-сортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 230 с.
-
8. Определитель сосудистых растений Среднего Урала / П.Л. Горчаковский, Е.А. Шурова, М.С. Князев и др. М.: Наука, 1994. 525 с.
-
9. Иллюстрированный определитель растений Пермского края / С.А. Овеснов, Е.Г. Ефимик, Т.В. Козьминых и др. / Под ред. д.б.н. С.А. Овеснова. Пермь: Книжный мир, 2007. 743 с.
-
10. Read D.J . The biology of mycorrhiza in the Ericalis // Botany. 1983. V. 61(3). P. 985-1004.
-
11. Nobel P.S . Internal Leaf Area and Cellular CO 2 Resistance: Photosynthetic Implications of Variations with Grow Conditions and Plant Species // Physiol. Plant. 1977. V. 40. P. 137-144.
-
12. A raus J.L., Alegre L., Tapia L., Calafell R., Serret M . Relationships Between Photosynthetic Capacity and Leaf Structure in Several Shade Plants // Amer. J. Bot. 1986. V. 68. P. 1760-1770.
THE CHARACTERISTICS OF CENOPOPULATIONS AND PECULIARITIES OF LEAF MESOSTRUCTURE ORTHILIA SECUNDA L. ON THE DISTURBED INDUSTRIAL LANDS
Ural Federal University named after the first President of Russia B.N. Yeltsin, Ekaterinburg
The results of study of age, morphological and spatial structure of cenopopulation Orthilia secunda L., mezostructure of the photosynthetic apparatus and type of mycorrhizae in plant communities in various technogenic substrates are presented in this paper. It is shown that in disturbed industrial lands, Orthilia secunda L. changes its morphology and its mezostructure parameters (decreasing in leaf thickness, size of chloroplasts, internal assimilation surface mezo-phyll and chloroplasts) compared with natural habitats.
Список литературы Характеристика ценопопуляций и особенности мезоструктуры листа Orthilia secunda L. на нарушенных промышленностью землях
- Мокроносов А.Т. Онтогенетический аспект фотосинтеза. М.: Наука, 1981. 196 с.
- Экологические основы и опыт биологической рекультивации нарушенных промышленностью земель/Т.С. Чибрик, Н.В. Лукина, Е.И. Филимонова, М.А. Глазырина. Екатеринбург: Изд-во Урал. ун-та, 2011. 286 с.
- Махонина Г.И. Экологические аспекты почвообразования в техногенных экосистемах Урала. Екатеринбург: Изд-во Урал. ун-та, 2003. 356 с.
- Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. 217 с.
- Онтогенетический атлас лекарственных растений: Научное издание. Т. IV Йошкар-Ола: МарГУ, 2004. 240 с.
- Фотосинтез: физиология, онтогенез, экология: Коллективная монография/Под ред. Е.С. Роньжиной. Калининград: Изд-во ФГТУ ВПО КГТУ, 2009. 372 с.
- Селиванов И.А. Микосимбиотрофизм, как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 230 с.
- Определитель сосудистых растений Среднего Урала/П.Л. Горчаковский, Е.А. Шурова, М.С. Князев и др. М.: Наука, 1994. 525 с.
- Иллюстрированный определитель растений Пермского края/С.А. Овеснов, Е.Г. Ефимик, Т.В. Козьминых и др./Под ред. д.б.н. С.А. Овеснова. Пермь: Книжный мир, 2007. 743 с.
- Read D.J. The biology of mycorrhiza in the Ericalis//Botany. 1983. V. 61(3). P. 985-1004.
- Nobel P.S. Internal Leaf Area and Cellular CO2 Resistance: Photosynthetic Implications of Variations with Grow Conditions and Plant Species//Physiol. Plant. 1977. V. 40. P. 137-144.
- Araus J.L., Alegre L., Tapia L., Calafell R., Serret M. Relationships Between Photosynthetic Capacity and Leaf Structure in Several Shade Plants//Amer. J. Bot. 1986. V. 68. P. 1760-1770.
