Характеристика морфологических изменений зубчатой извилины гиппокампа взрослых крыс при воздействии комбинированного стресса

Автор: Смирнов А.В., Шмидт М.В., Экова М.Р., Медников Д.С., Тюренков И.Н., Бородин Д.Д., Рустанова А.С.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Рубрика: Морфология
Статья в выпуске: 4 (44), 2014 года.
Бесплатный доступ
После воздействия комбинированного стресса со сменой разномодальных стрессоров (шума, вибрации, пульсирующего яркого света) по стохастической схеме на фоне ограничения подвижности и повышения температуры в камере за время стрессирования от 20-22 °С до 25-27 °С у животных после семи ежедневных 30-минутных сеансов в зубчатой извилине гиппокампа взрослых (12 месяцев) крыс отмечены признаки повреждения нейронов, а также нарушения кровообращения в сосудах микроциркуляторного русла.
Зубчатая извилина, гиппокамп, нейрон, крыса, стресс
Короткий адрес: https://sciup.org/142149127
IDR: 142149127 | УДК: 616.091.8:616.-092.9:57.042
Morphological changes in hippocampal dentate gyrus of adult rats exposed to combined stress
The daily exposure of rats to 30 minutes' sessions of combined stress induced by removable multi-modal stressors (noise, vibration, pulsating bright light ) using the stochastic scheme coupled with restricted mobility and a rise in the chamber temperature from 20-22 °С to 25-27 °С for seven days made it possible to reveal the signs of neuronal damage in hippocampal dentate gyrus of adult rats (12 months) as well as circulatory disturbances in the microvasculature.
Текст научной статьи Характеристика морфологических изменений зубчатой извилины гиппокампа взрослых крыс при воздействии комбинированного стресса
Структуры головного мозга активно участвуют в реализации различных проявлений стрессовых реакций. В ответ на стресс происходит повышение секреции глюкокортикоидов – кортизола у человека и кортикостерона у крыс. Нейроны СА1 зоны и зубчатой извилины гиппокампа экспрессируют высокое количество глюкокортикоидных рецепторов и поэтому особенно восприимчивы к гормонам стресса [10]. Воздействие стресса или гормонов стресса влияет на синаптическую пластичность гиппокампа, нейрохимию, нейрогенез, морфологию и апоптоз нейронов [6, 8], а также нарушает гиппокамп-зависимые процессы обучения и памяти [5]. Зубчатая извилина является важным компонентом гиппокампальной цепи, принимает участие в эпизодической и пространственной памяти. Нейроны гранулярного слоя зубчатой извилины получают возбуждающие инпульсы из энтори-альной коры и передают возбуждающие проекции в СА3 зону гиппокампа. Экспериментальные данные показывают, что новообразованные нейроны гранулярного слоя зубчатой извилины образуются в субгранулярном слое при нейрогенезе у взрослых, благодаря своим уникальным свойствам, играют важную роль в определении образов [4]. Острый и хронический стресс, а также лечение кортикостероидами, как правило, подавляют нейрогенез путем воздействия на клеточную пролиферацию, дифференцировку и выживаемость нейронов [14], а лечение антидепрессантами может повысить количество нейронов в гранулярном слое зубчатой извилины [7]. Учитывая, что гиппокамп связан с регулированием гипоталамо-гипофизарно-надпочечникового механизма, (ГГНМ) исследования R. J. Schloesser и соавторов [13] показывают, что подавление нейрогенеза в зубчатой извилине приводит к усилению ответа со стороны ГГНМ после воздействия стрессора, предполагая, что новообразованные нейроны в зубчатой извилине принимают участие в регуляции ГГНМ.
Таким образом, морфофункциональные изменения в зубчатой извилине гиппокампа играют важную роль при стрессовом воздействии.
ЦЕЛЬ РАБОТЫ
Выявить структурные изменения в гранулярном слое зубчатой извилины взрослых 12-месячных крыс на фоне комбинированного стресса.
МЕТОДИКА ИССЛЕДОВАНИЯ
Моделирование комбинированного стресса (пульсирующий свет, громкий звук, вибрация) проводилось по методике [3] на 20 нелинейных крысах-самцах в возрасте 12 месяцев (ФГПУ Питомник лабораторных животных «Рапполово») с соблюдением «Правил проведения работ с использованием экспериментальных животных» (приказ № 755 от 12.08.1977 г. МЗ СССР). Животные были разделены на 2 группы. Первая группа – контрольные крысы (n = 10), 2-я группа – стрессированные крысы (n = 10). Эвтаназию проводили с помощью гильотинного метода с использованием «Гильотины для крыс» (AE0702, производитель «Open Science»). Головной мозг фиксировали в нейтральном забуференном 10%-м формалине. Парафиновые срезы толщиной 5 мкм окрашивали гематоксилином и эозином, тионином по методу Ниссля. Определяли степень повреждения нейронов методом подсчета удельного количества сморщенных нейронов с гиперхроматозом цитоплазмы в гранулярном слое зубчатой извилины.
Статистическую обработку данных проводили с использованием пакетов программ Statistica 6,0 (StatSoft, USA). Обобщенные данные представляли в виде медианы (Me) с указанием интерквартильного интервала [Q1; Q3], где Q1 – 25 процентиль, Q3 – 75 процентиль. Различия между группами оценивали по критерию Манна–Уитни (Mann-Whitney, U-test) и считали статистически значимыми при p < 0,05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
При микроскопическом изучении отделов зубчатой извилины у контрольных животных было выявлено, что гранулярный слой состоял из 5–6 рядов компактно расположенных нейро-цитов, нейроны характеризовались округлой формой ядра и близкой к округлой формой перикариона, равномерным распределением субстанции Ниссля в цитоплазме. Обнаруживались лишь единичные сморщенные гиперхром-ные клетки (рис. 1).
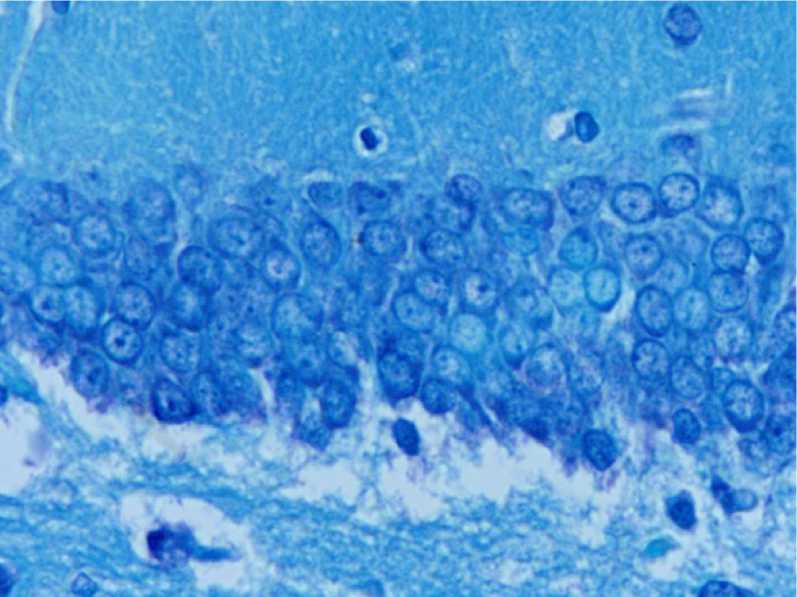
Рис. 1. Гистологическое строение гранулярного слоя зубчатой извилины взрослой крысы в возрасте 12 мес. (контроль).
Окраска тионином по методу Ниссля. Ув. х400
У стрессированных животных часть нейронов гранулярного слоя зубчатой извилины характеризовалась разнообразными неспецифическими изменениями в виде гиперхроматоза, хроматолиза и сморщивания. Наблюдались участки очаговых выпадений нейроцитов, а также уменьшение, по сравнению с контролем, плотности расположения нейронов и ширины слоев.
Кроме того, выявлялись сморщенные нейроны с интенсивной базофилией цитоплазмы перикариона, а в ряде случаев кариопикно-зом, кариолизисом и уменьшением размеров перикариона (рис. 2). Удельное количество сморщенных нейронов с гиперхроматозом цитоплазмы составило 5 [0;15] % (р = 0,0006).
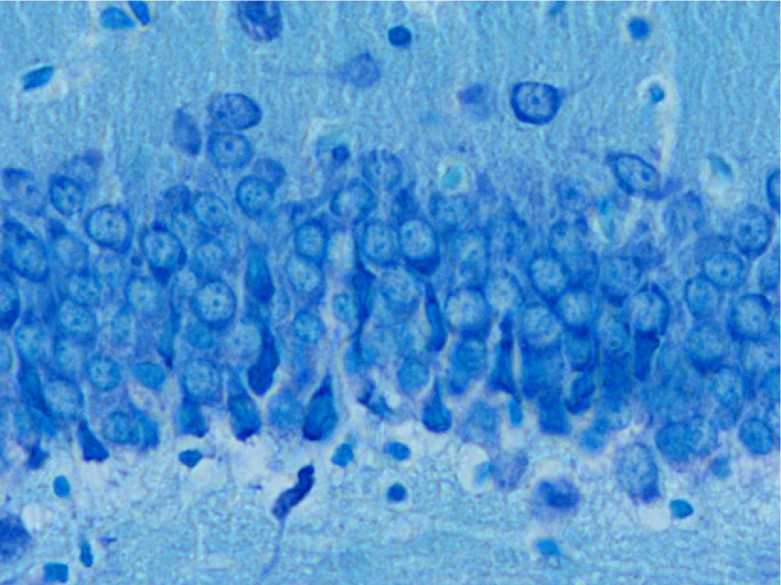
Рис. 2. Гистологическое строение гранулярного слоя зубчатой извилины взрослой крысы в возрасте 12 мес., находившейся в течение 7 суток под влиянием комбинированного стресса.
Окраска тионином по методу Ниссля. Ув. х400
Под воздействием комбинированного стресса в сосудах микроциркуляторного русла наблюдались неспецифические признаки нарушений кровообращения в виде полнокровия капилляров, спазма артериол, стаза эритроцитов.
Обнаруженные нами признаки структурных повреждений в нейронах гранулярного слоя зубчатой извилины гиппокампа косвенно соотносятся с выявленными морфологическими изменениями в СА3 поле гиппокампа при длительном комбинированном стрессовом воздействии [1, 2]. Механизмы повреждения нейронов гиппокампа могут быть связаны с эксайтотокси-ческим действием высокой концентрации глутамата, что ведет к его резкому возрастанию в окружающей синапс внеклеточной среде. Считается, что взаимодействие глутамата с экстра-синаптическими NMDA-рецепторами GluN2B запускает механизмы апоптоза путем активации сигнальных нейрональных комплексов смерти (NDC) [12, 15].
Другой механизм повреждения нейронов при стрессе может быть связан с нарушениями кровообращения в сосудах микроциркуляторного русла гиппокампа, что рассматривается нами как результат ишемии, снижения нейротрофных и нейропротекторных факторов эндотелия сосудов. Пролиферация нейронов в зубчатой извилине гиппокампа происходит в ограниченном кластере, расположенном рядом с сосудами. Нарушение процесса нейрогенеза часто сочетается с изменениями в ангиогенезе. Кроме того, оба этих процесса регулируются схожими модулирующими факторами, например, сосудистым эндотелиальным фактором роста (VEGF) и его рецептором FLK-1. Изменения в концентрации VEGF и экспрессии рецепторов VEGF приводят к изменению клеточной пролиферации и выживаемости как in vitro, так и в естественных условиях [9, 11].
ЗАКЛЮЧЕНИЕ
Таким образом, полученные нами данные свидетельствуют о том, что при длительном воздействии комбинированного стресса у 12-месячных крыс-самцов обнаруживаются признаки обратимых и необратимых повреждений нейронов гранулярного слоя зубчатой извилины гиппокампа, а также определяются признаки нарушений кровообращения в сосудах микроцир-куляторного русла.
Список литературы Характеристика морфологических изменений зубчатой извилины гиппокампа взрослых крыс при воздействии комбинированного стресса
- Смирнов А. В., Тюренков И. Н., Шмидт М. В. и др.//Вестник ВолгГМУ. -2013. -№ 2. -С. 14-17.
- Смирнов А. В., Шмидт М. В., Экова М. Р. и др.//Волгоградский научно-медицинский журнал. -2013. -№ 4. -С. 14-17.
- Тюренков И. Н., Багметова В. В., Чернышева Ю. В. и др.//Российский физиологический журнал им. И. М. Сеченова. -2013. -Т. 99, № 9. -С.1045-1056.
- Benarroch E E//Neurology. -2013. -Vol. 81, № 16. -P. 1443-1452.
- Conrad C. D.//Progress in Neuro-Psychopharmacology and Biological Psychiatry. -2009. -Vol. 34, № 5. -P. 742-755.
- Conrad C. D.//Reviews in the Neurosciences. -2008. -Vol.19, № 6. -P. 395-412.
- David D. J., Samuels B. A, Rainer Q., et al.//Neuron. -2009. -Vol. 62, № 4. -P. 479-493.
- Fa M., Xia L., Anunu R., et al.//Neurobiol. Learn. Mem. -2014. -Vol. 112. -P. 53-60.
- Heine V. M., Zareno J., Maslam S., et al.//European Journal of Neuroscience. -2005. -Vol. 21. -P. 1304-1314.
- Henk K., Marian Joёls M.//J. Neurophysiol. -2003. -Vol. 89. -P. 625-633.
- Jin K., Zhu Y., Sun Y., et al.//Proc. Natl. Acad. Sci. USA. -2002. -Vol. 99. -P. 11946-11950.
- Lee J. M, Zipfel G. J., Choi D. W.//Nature. -1 999. -Vol. 399. -P. A7-14.
- Schloesser R. J., Manji H. K., Martinowich K.//NeuroReport. -2009. -Vol. 20. -P. 553-557.
- Schoenfeld T. J., Gould E.//Experimental Neurology. -2012. -Vol. 233, № 1. -P. 12-21.
- Tu W, Xu X., Peng L., et al.//Cell. -2010. -Vol. 140. -P. 222-234.
