Характеристика популяций Puccinia reconditа Rob. ex Desm. из Среднего Поволжья по частотам вирулентности к LR генам устойчивости пшеницы в 2014 году

Автор: Тырышкин Лев Геннадьевич, Сюков Валерий Владимирович, Захаров Владимир Григорьевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Генетика и селекция
Статья в выпуске: 4-3 т.17, 2015 года.
Бесплатный доступ
По результатам исследований 2014 г. подтверждена принадлежность ульяновской и безенчукской популяций Puccinia recondita к единому ареалу формообразования и распространения листовой ржавчины. В то же время показана значительная схожесть структуры средневолжских субпопуляций со структурой популяции из Северного Кавказа. В 3-х регионах отсутствовали клоны патогена, вирулентные к проросткам линий с генами Lr 9, Lr 19, Lr 24, Lr 28, Lr 47 и Lr Ag#2. К гену Lr 29 был вирулентен только один клон из Безенчука. Показана согласованная динамика концентрации генов вирулентности plr1, plr2a, plr23, plr25, plr35 в течение 2011-2014 гг. в субпопуляциях Среднего Поволжья. По большинству генов существенного изменения концентраций не наблюдалось. По генам plr1, plr25 и plr35 синхронно в обеих субпопуляциях наблюдается снижение концентрации, а по генам plr2а и plr23 - повышение концентрации. Обсуждается возможность расширения фенотипических маркеров возбудителя ржавчины за счет изучения вирулентности монопустульных изолятов на отрезках листьев дифференциаторов, помещенных на различные субстраты (растворы бензимидазола, солей азота и калия).
Листовая бурая ржавчина, яровая пшеница, гены вирулентности, гены устойчивости
Короткий адрес: https://sciup.org/148203912
IDR: 148203912 | УДК: 577.21:633.11:632.937.14
Population characteristics of Puccinia recondita Rob. ex Desm. from the Middle Volga frequency virulence by LR wheat resistance genes in 2014
According to the research in 2014 confirmed the affiliation of Ulyanovsk and Bezenchuk populations of Puccinia recondita in a single areal formation and distribution of leaf rust. At the same time shows a significant similarity between the structure of Middle subpopulations with the structure of the population of the North Caucasus. In 3 regions no clones of the pathogen virulent to seedlings lines with genes Lr 9, Lr 19, Lr 24, Lr 28, Lr 47 and Lr Ag # 2. By gene Lr 29 was virulent Only one clone of Bezenchuk. It is shown that the dynamics of consistent concentration of virulence genes plr1, plr2a, plr23, plr25, plr35 during 2011-2014. subpopulations in the Middle Volga. For most of the genes significantly change the concentrations observed. As genes plr1, plr25 and plr35 synchronously in both subpopulations decrease concentration, and the genes and plr2a plr23 - increasing the concentration. We discuss the possibility of expanding the phenotypic markers rust pathogen by examining the virulence of isolates single-pustuled leaf segments differentiators placed on various substrates (solutions benzimidazole, nitrogen and potassium salts).
Текст научной статьи Характеристика популяций Puccinia reconditа Rob. ex Desm. из Среднего Поволжья по частотам вирулентности к LR генам устойчивости пшеницы в 2014 году
Листовая бурая ржавчина (возбудитель Puccinia reconditа Rob. ex Desm . syn.: Puccinia triticina Erikss) – одна из наиболее распространенных и вредоносных болезней мягкой пшеницы Triticum ae s ti v um L . во многих зонах возделывания культуры, в том числе и в Среднем Поволжье Российской Федерации. Изучение частот вирулентности к конкретным Lr генам устойчивости традиционно проводятся для идентификации эффективных на данный момент генов резистентности к болезни, определения степени сходства субпопуляций из различных частей регионов, что в свою очередь ложится в основу программ рационального территориального размещения имеющихся эффективных генов устойчивости. Подобная работа стабильно проводится в США и
Канаде [1, 2], периодически в странах Западной и Восточной Европы [3-9], Азии [10, 11]. В России в последние десятилетия эту работу ведут в основном в Северокавказском [12, 13], Северо-Западном [14], меньше Нижневолжском [15, 16] регионах.
Проведённое нами изучение [17] полиморфизма средневолжской популяции Puccinia recondita в 2011–2012 гг. показало, что у растений в стадии проростков высокоэффективными являются гены Lr 9 и Lr 41 ( Lr 39+), несколько меньшей эффективностью обладали гены Lr 19, Lr 24, Lr 28, Lr 29 и Lr 47. Сравнение популяций из Безенчука и Ульяновска по частотам вирулентности к 28-и слабоэффективным генам устойчивости показало их крайнюю степень сходства и, скорее всего, принадлежность к одному ареалу формообразования и распространения листовой ржавчины. В то же время полученные данные указывали и на высокое сходство данных популяций с таковыми из Северного Кавказа и Северо-Западного региона России.
Цель настоящего исследования – изучение частот вирулентности Puccinia recondita из двух районов Среднего Поволжья в сравнении с популяцией из Южного Дагестана в 2014 г., а также определение степени сходства в данном году между выборками клонов патогена из 3-х регионов России.
МАТЕРИАЛЫ И МЕТОДЫ
Монопустульные изоляты P. recondita выделяли из популяций патогена, собранных в Среднем Поволжье (пос. Безенчук Самарской области и пос. Тимирязевский Ульяновской области) и Северном Кавказе (г. Дербент, Дагестанская опытная станция ВИР) в 2014 г. и поддерживали на отрезках листьев универсально восприимчивого сорта пшеницы Ленинградка. Всего из каждой популяции было выделено и проанализировано по 100 изолятов возбудителя листовой ржавчины пшеницы.
Семена почти изогенных линий пшеницы сорта Thatcher с генами устойчивости к листовой ржавчине Lr 1, Lr 2, Lr 2a, Lr 2b, Lr 2c, Lr 3, Lr 3ka, Lr 9, Lr 10, Lr 11, Lr 12, Lr 13, Lr 14b, Lr 16, Lr 18, Lr 19, Lr 20, Lr 21, Lr 23, Lr 24, Lr 25, Lr 26, Lr 30, Lr 32, Lr 33, Lr 34, Lr 35, Lr 36, Lr 37, Lr 38, а также образцов с генами Lr 27+31, Lr 28, Lr 29, Lr 45, Lr 46, Lr 47, Lr 48, Lr 49 и Lr Ag#2 высевали в 3 кюветы на смоченную водой вату. После прорастания семян кюветы помещали на светоустановку (20–22°С, постоянное освещение – 2 500 люкс). Растения в стадии одного листа опрыскивали водными суспензиями уредоспор популяций возбудителя из 3-х регионов, на сутки закрывали полиэтиленом и помещали в темноту. Затем полиэтилен снимали и кюветы возвращали на светоустановку. Через 12 дней оценивали типы реакции по шкале Майнса и Джексона [18] 0 – отсутствие симптомов болезни; 0; – некрозы без пустул; 1 – очень мелкие пустулы, окруженные некрозом; 2 – пустулы среднего размера, окруженные некрозом или хлорозом; 3 –крупные пустулы без некроза; X – наличие на одном листе пустул разного типа реакции; р.е.п. – очень редкие пустулы восприимчивого типа 3.
Отрезки первых листьев (длина 0,7–1 см) выращенных на вате растений тех же линий и образцов с Lr генами устойчивости раскладывали в строчку на смоченную водой вату и инокулировали монопустульными изолятами возбудителя листовой ржавчины. Типы реакции на заражение патогеном учитывали на 6–7-е сутки после инокуляции (в данном случае типы Х и е.п. отсутствовали). Типы реакции 0–2 соответствуют авирулентности, тип 3 – вирулентности патогена. Типы реакции отрезков листьев изучаемого набора образцов совпадали с типами реакций на заражение интактных растений теми же клонами (данные не приводятся). Для оценки степени сходства популяций P. recondita из разных регионов использовали метод парных корреляций частот вирулентности [19].
По 15-20 отрезков листьев проростков (длина 2 см) почти изогенных линий и образцов с вышеперечисленными Lr генами раскладывали в кюветы на вату, смоченную водой, водным раствором бензимидазола (60 мг/л), раствором азотнокислого аммония (1,29 г/л) и хлористого калия (0,48 г/л). Отрезки листьев опрыскивали одинаковым объемом водных суспензий уредоспор популяций возбудителя ржавчины из трех регионов (концентрация спор 30 x 103 /мл) с помощью ручного пульверизатора; кюветы закрывали полиэтиленом и на 12 ч помещали в темноту, затем полиэтилен снимали, кюветы закрывали стеклом и помещали на светоустановку. Через 7 суток после инокуляции подсчитывали количество пустул восприимчивого типа на каждом отрезке листа; для дальнейшего анализа рассчитывали среднее число пустул на одном отрезке.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Высокую эффективность против всех изучаемых популяций при заражении интактных проростков в 2014 г. проявили гены устойчивости Lr 9, Lr 19, Lr 24, Lr 28, Lr 47 и Lr Ag#2 . На линии с геном Lr 29 отмечено наличие очень редких вирулентных клонов (табл. 1). Основным отличием данного результата от полученного в аналогичной работе в 2012 г. является отсутствие монопустульных изолятов гриба, вирулентных к Lr 24, Lr 28 и Lr 47.
В основном эти данные подтверждаются и анализом вирулентности монопустульных изо-лятов (табл. 2). При анализе 100 клонов из каждой популяции не выявлено изолятов Puccinia recondita , вирулентных к перечисленным генам. К гену Lr 29 был вирулентен только один клон из Безенчука.
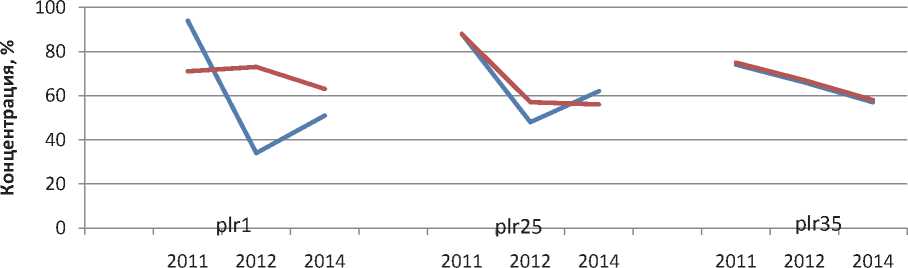
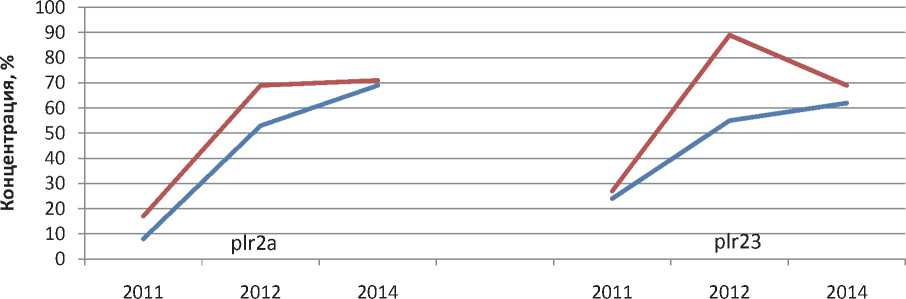
Анализ динамики вирулентностей в 20112014 гг показывает, что по большинству генов существенного изменения концентраций не наблюдалось. По генам plr1, plr25 и plr35 синхронно в обеих субпопуляциях наблюдается снижение концентрации (рис. 1), а по генам plr2а и plr23 – повышение концентрации (рис. 2).
Результаты подсчета количества пустул возбудителя ржавчины 3-х популяций после заражения отрезков листьев образцов пшеницы, помещенных на вату, смоченную различными растворами приведены в табл. 3.
Коэффициенты корреляции между частотой вирулентных клонов к помещенным на воду отрезкам листьев конкретного образца пшеницы и средним количеством пустул на отрезках листьев в воде были близки к 1 (для популяций из Дербента, Ульяновска и Безенчука – 0,987, 0,981 и 0,978, соответственно).
Это связано с тем, что большинство образцов, используемых в работе, являются почти изогенными линиями одного сорта и не различаются по уровню неспецифической устойчивости; следовательно, различия между линиями в количестве пустул при заражении популяцией патогена могут быть связаны только с различиями в частоте встречаемости в данной популяции вирулентных клонов P. recondita . Это в свою очередь указывает на возможность сравнения различных популяций
Таблица 1. Типы реакции линий и сортов пшеницы с Lr генами устойчивости на заражение популяциями возбудителя листовой ржавчины пшеницы. Интактные проростки. 2014 г.
При заражении одной и той же популяцией отрезков листьев, помещенных на вату, смоченную водой и растворами различных веществ, корреляция числа пустул на изучаемых линиях была низкой либо средней. Так, например, для выборки из Ульяновска коэффициенты корреляции числа пустул на отрезках листьев в воде и на раство- рах аммонийной селитры, хлористого калия и бензимидазола (в анализ взяты только линии, обладающие дифференцирующей способностью хотя бы в одном варианте опыта) были равны 0,47, 0,61 и 0,59, соответственно (коэффициенты детерминации – 0,22, 0,38 и 0,35, соответственно). Это подтверждает ранее сделанный вывод [20, 21], что данные вещества влияют не на неспецифическую устойчивость изогенных линий сорта Тэтчер, а вызывают специфическое изменение вирулентности ряда клонов патогена. Это в свою
Таблица 2. Частота клонов возбудителя листовой ржавчины, вирулентных к проросткам (отрезки листьев в воде) образцов пшеницы с Lr генами устойчивости, (%) (2014 г.)
Годы
Ульяновск Безенчук
Рис. 1. Динамика концентрации генов вирулентности plr1, plr25 и plr35 в средневолжских субпопуляциях Puccinia recondita
Годы
Ульяновск Безенчук
Рис. 2. Динамика концентрации генов вирулентности plr2a и plr23 в средневолжских субпопуляциях Puccinia recondita
Коэффициенты корреляции между числом пустул на отрезках листьев линий после заражения 3-мя популяциями возбудителя листовой ржавчины приведены в табл. 4. Высокие, во многих случаях близкие к единице значения коэффициентов указывают на высокую степень сходства выборок из 2-х регионов Среднего Поволжья. Сходство этих популяций с таковой из Дагестана также высоко, однако, слегка ниже согласно коэффициентам корреляции числа пустул возбудителя ржавчины на линиях, проростки которых находились на вате, смоченной растворами бензимидазола и аммиачной селитры (табл. 4).
ЗАКЛЮЧЕНИЕ
Полученные в 2014 г. данные по изучению структуры популяций возбудителя листовой ржавчины пшеницы по частотам вирулентности к конкретным Lr генам устойчивости подтверждают ранее сделанный вывод о принадлежности популяций из Ульяновска и Без-енчука к единому ареалу распространения патогена. Подтвержден также вывод о высокой степени схожести данных популяций с выборкой из Южного Дагестана. Абсолютно эффективными на стадии проростков в 3-х регионах в 2014 г. были гены Lr 9, Lr 19, Lr 28, Lr 24 и Lr Ag#2. Крайне низкими частотами вирулентных клонов характеризовались гены Lr 29 и Lr 47. Полученные данные могут быть полезны при разработке программ рационального территориального размещения известных и новых эффективных генов резистентности пшеницы к листовой ржавчине.
Разработанный подход по сравнению числа пустул с помещенных на различные субстраты
Таблица 3. Среднее количество пустул возбудителя листовой ржавчины на отрезках листьев проростков образцов пшеницы с Lr генами устойчивости (2014 г.)
*- отрезки листьев на вате, смоченной 1. водой; растворами 2. NH4NO3; 3. KCl; 4. бензимидазола.
Таблица 4. Коэффициенты корреляции между средним числом пустул P. triticina на образцах пшеницы, зараженных различными популяциями патогена. 2014
|
Популяции |
Ульяновск |
Дербент |
|
Отрезки листьев в воде |
||
|
Безенчук Ульяновск |
0,985 |
0,991 0,982 |
|
Отрезки листьев в растворе аммиачной селитры |
||
|
Безенчук Ульяновск |
0,935 |
0,969 0,873 |
|
Отрезки листьев в растворе хлористого калия |
||
|
Безенчук Ульяновск |
0,979 |
0,982 0,977 |
|
Отрезки листьев в растворе бензимидазола |
||
|
Безенчук Ульяновск |
0,943 |
0,873 0,900 |
*- отрезки листьев на вате, смоченной 1. водой; растворами 2. NH4NO3; 3. KCl; 4. бензимидазола
Список литературы Характеристика популяций Puccinia reconditа Rob. ex Desm. из Среднего Поволжья по частотам вирулентности к LR генам устойчивости пшеницы в 2014 году
- Kolmer, J.A. Physiologic specialization of Puccinia triticina on wheat in the United States in 2012/J.A.Kolmer, M.E.Hughes//Plant Disease, 2014. Vol.98. №8. P.1145-1150 DOI: 10.1094/PDIS-12-13-1267-SR
- McCallum, B.D. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2009/B.D.McCallum, P.Seto-Goh, A.Xue//Canad.J.Plant Pathology, 2013. Vol.35. Iss.3. P.338-345
- Del Olmo, A.I. Physiologic specialization of Puccinia triticina in Andalusia (Spain) in 2004 and 2005/A.I.del Olmo, J.C.Sillero, D.Rubiales//Options Méditerranéennes, Series A, 2008. №81. P.169-171.
- Elyasi-Gomari, S. Virulence polymorphism of Puccinia recondita f.sp. tritici and effectiveness of Lr-genes for leaf rust resistance of wheat in Ukraine/S.Elyasi-Gomari, V.K.Panteleev//Plant Disease, 2006. Vol.90. №7. P.853-857
- Hanzalova, A. Physiologic specialization of wheat leaf rust (Puccinia triticina Eriks.) in the Slovak Republic in 2009-2011/A.Hanzalova, T.Sumikova, J.Huszar, P.Bartos//Czech.J.Plant Breed., 2012. Vol.48. №3. P.101-107.
- Lin, V. Virulence frequencies of Puccinia triticina in Germany and the European regions of the Russian Federation/V.Lin, E.Gultyaeva//J.Phytopathology, 2007. Vol.155. №1. P.13-21
- Mesterhazy, A. European virulence survey for leaf rust in wheat/A. Mesterhazy, P.Bartos, H.Goyeau et al//Agronomie, 2000. Vol.20. Iss.7. P.793-804
- Park, R.F. Physiologic specialization and pathotype distribution of Puccinia recondita in Western Europe, 1995/R.F.Park, F.G.Felsenstein//Plant Pathology, 1998. Vol.47. Iss.2. P.157-164
- Булойчик, А.А. Частота встречаемости генов вирулентности в Белорусских популяциях Puccinia triticina Erikss/А.А.Булойчик, В.С.Борзяк, Е.А.Волуевич//Молекулярная и прикладная генетика, 2010. Т.11. С.68-73
- Агабаева А.Ч. Патогенные свойства возбудителя листовой ржавчины пшеницы (Puccinia triticina Eriks) в Казахстане//А.Ч.Агабаева, Ш.С.Рсалиев//Новости науки Казахстана, 2013. Вып.1
- Elyasi-Gomari, S. Virulence of Puccinia triticina on wheat in Iran/S.Elyasi-Gomari//African J.Plant Sci., 2010. Vol.4. №2. P.26-31
- Волкова, Г.В. Структура и изменчивость популяций возбудителей бурой и жёлтой ржавчины пшеницы на Северном Кавказе и обоснование приёмов управления внутрипопуляционными процессами: Дисс.. докт. биол. наук. С-Петербург, 2006. 426 с.
- Кудинова, О.А. Взаимосвязь вирулентности и RAPD-полиморфизма Северокавказской популяции возбудителя бурой ржавчины пшеницы: Автореф.. канд. биол. наук. Краснодар, 2012. 23 с.
- Гультяева, Е.И. Структура популяций Puccinia triticina по вирулентности и ДНК-маркёрам в Северо-Западном регионе России в 2007 г./Е.И.Гультяева, Е.Косман, А.П.Дмитриев, О.А.Баранова//Микология и фитопатология, 2011. Т.45. №1. С.70-81
- Иванова, О.В. Источники устойчивости яровой пшеницы к бурой ржавчине и изменчивость структуры популяции возбудителя в условиях Нижнего Поволжья: Автореф.. канд. с.-х. наук. Саратов, 2013. 22 с.
- Маркелова, Т.С. Изучение структуры и изменчивости популяции бурой ржавчины пшеницы (Puccinia recondita f.sp. tritici Rob ex Desm.) в Поволжье/Т.С. Маркелова//Агро XXI, 2007. № 46. С.37-40
- Тырышкин, Л.Г. Сравнительная характеристика вирулентности Puccinia recondita Rob. ex Desm. syn.: Puccinia triticina Erikss. в Среднем Поволжье/Л.Г.Тырышкин, В.Г.Захаров, В.В. Сюков//Вавиловский журнал генетики и селекции. 2014. Том 18, № 2. С. 202-206.
- Mains, E.B. Physiological specialization in leaf rust of wheat, Puccinia triticina Erikss/E.B.Mains, H.S.Jackson//Phytopathology. 1926. Vol. 16. № 1. P. 89-120.
- Большев, Л.Н. Таблицы математической статистики/Л.Н.Большев, Н.В. Смирнов. М., 1983. 416 с.
- Тырышкин, Л.Г. Изменение вирулентности возбудителя листовой ржавчины пшеницы под действием элементов минерального питания/Л.Г.Тырышкин//Известия Санкт-Петербургского государственного аграрного университета. 2014. № 35. С. 85-89.
- Тырышкин, Л.Г. Повышение частичной устойчивости к листовой ржавчине почти-изогенных линий пшеницы с Lr генами под действием бензимидазола -результат изменения вирулентности патогена/Л.Г.Тырышкин//Известия Санкт-Петербургского государственного аграрного университета. 2014. № 34. С. 50-54.
