Характеристика регуляторных Т-клеток у больных раком яичников

Автор: Курганова Е.В., Тихонова М.А., Лебедева В.А., Ласкавая Е.Г., Коваленко В.Ф., Останин А.А., Черных Е.Р.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Экспериментальные исследования
Статья в выпуске: 6 (30), 2008 года.
Бесплатный доступ
Охарактеризованы регуляторные Т-клетки (Treg) у 10 больных раком яичника (РЯ) III-IV стадии. Установлено, что по сравнению с периферической кровью в асцитической жидкости больных РЯ повышено количество CD4-CD25-CD4-CD25highTreg. Выявленный факт может свидетельствовать о направленной миграции и накоплении естественных Treg в зоне опухолевого микроокружения. Увеличение CD4-FOXP3- и CD8-FOXP3- регуляторных Т-клеток имеет более генерализованный характер и регистрируется как в периферической крови, так и в асцитической жидкости. Больные РЯ характеризуются выраженным 14-кратным снижением баланса соотношения цитотоксических CD8-Pfn-Т-лимфоцитов и CD4-FOXP3-Treg - 0,4 ± 0,1 против 5,7 ± 2,0 у здоровых женщин (pU
Регуляторные т-клетки, цитотоксические т-лимфоциты, рак яичников
Короткий адрес: https://sciup.org/14054911
IDR: 14054911 | УДК: :
Regulatory T- cells in ovarian cancer
The aim of the study was to investigate the regulatory T cells (Treg) in the patients with stage III-IV ovarian cancer (OC, n=10). It was revealed that the level of СD4-СD25- /СD4-СD25high Treg in OC patients was increased in ascites compared to that in peripheral blood. These data strongly suggest the directional traffic and accumulation of natural Treg in tumor microenvironment. The level of CD4-FOXP3- and CD8-FOXP3- cells was increased both in ascites and in peripheral blood of patients with OC. Our results indicate that expansion of these Treg subsets is more generalized. It was also shown, that patients with OC are characterized by marked (14-fold) decrease of CD8-Pfn-/CD4-FOXP3- ratio (0,4±0,1 vs 5,7±2,0 in healthy woman, pU
Текст научной статьи Характеристика регуляторных Т-клеток у больных раком яичников
Известно, что опухолевые клетки могут распознаваться и уничтожаться иммунной системой. Однако в условиях клинической манифестации и прогрессии опухоли иммунный ответ оказывается несостоятельным, а эффективность иммунотерапии регистрируется лишь в ограниченных случаях. Одна из причин неадекватного иммунного ответа и низкой эффективности иммунотерапии связана с возрастанием количества регуляторных Т-клеток с супрессорной активностью (Treg) [24].
Популяция регуляторных Т-клеток является достаточно гетерогенной. Наибольший интерес в последние годы привлекают естественные регуляторные СD4+СD25+Т-клетки, которые играют важную роль в поддержании периферической толерантности к собственным антигенам за счет подавления аутореактивных
Т-лимфоцитов [18, 19]. Поскольку опухолевые клетки экспрессируют аутоантигены, рост опухоли сопровождается появлением аутореактивных Т-клеток, что непременно влечет за собой компенсаторное возрастание Treg [1]. Наряду с естественными Treg, имеющими тимическое происхождение, выделяют также индуцированные регуляторные Т-клетки, которые могут генерироваться из СD4+ и СD8+Т-клеток периферической крови [15, 18].
В экспериментальных моделях показано, что опухолевый рост сопровождается увеличением численности регуляторных Т-клеток. Причем их истощение приводит к отторжению или уменьшению массы опухоли у мышей, что демонстрирует ингибирующий эффект Treg на противоопухолевый иммунный ответ [19, 21]. Возрастание уровня регуляторных
СD4+CD25+Т-клеток показано также у больных с различными формами солидных опухолей и гемобластозов. Увеличение этих клеток выявляется как в локальном опухолевом микроокружении, так и в периферической крови, что свидетельствует о генерализованном характере опухоль-индуцированной супрессии [4, 6, 12, 29]. Причем, по данным ряда авторов, количество Treg коррелирует с показателями выживаемости. Поэтому их определение в клинической практике может иметь прогностическое значение [3, 10, 16, 20, 25].
Традиционно Treg оценивают как CD4+CD25+ клетки, поскольку они ко-экспрессируют молекулу СD25. Однако у человека популяция CD4+CD25+ клеток включает не только естественные, но и индуцированные Т reg, а также активированные CD4+Т-лимфоциты [4, 18, 27]. Определенным прогрессом на пути идентификации Treg стало выявление транскрипционного фактора FOXP3. Оказалось, что данный транскрипционный фактор является критическим для развития регуляторных Т-клеток и детерминирует появление у них супрессорной активности [11, 26]. Поэтому экспрессия FOXP3 рассматривается на сегодняшний день в качестве наиболее специфичного маркера Treg [9, 14, 17].
Исследованиями Treg при раке яичников показано, что возрастание CD4+CD25+ клеток выявляется исключительно в зоне опухолевого микроокружения (в асцитической жидкости и среди опухольинфильтрирующих лимфоцитов), но не в периферической крови [7]. Однако популяция CD4+FOXP3+ клеток при раке яичников до сих пор не оценивалась. Отсутствуют данные и о численности регуляторных CD8+FOXP3+ Т-клеток, возрастание которых описано при некоторых других локализациях злокачественных опухолей [13]. Исходя из этого, целью данной работы стало изучение различных субпопуляций регуляторных Т-клеток в периферической крови и асцитической жидкости у больных раком яичников.
Материал и методы
В исследование были включены 10 пациенток, больных раком яичников (РЯ) III–IV стадии. Возраст обследуемых варьировал от 25 до 76 лет (в среднем 56,7 ± 4,5 года). Для иммунологиче- ского исследования у больных перед операцией проводили забор 20 мл периферической крови, во время операции – асцитической жидкости. Все пациентки были обследованы после получения информированного согласия. Контрольную группу составили 20 здоровых женщин, сопоставимых по возрасту.
Мононуклеарные клетки (МНК) выделяли из гепаринизированной венозной крови центрифугированием в градиенте плотности фиколла-верографина. Клетки асцитической жидкости (АЖ) получали путем центрифугирования в течение 10 мин при 1500 об/мин. МНК в концентрации 0,1х106/лунку культивировали в 96-луночных круглодонных планшетах в среде RPMI-1640 (Sigma-Aldrich), дополненной 0,3 мг/мл L-глютамина, 5мМ HEPES-буфера, 100 мкг/мл гентамицина и 10 % инактивированной сыворотки доноров АВ (IV) группы крови при 370С в СО2-инкубаторе. Для стимуляции клеток использовали моноклональные анти-CD3 антитела ICO-90 (анти-CD3, «Медбиоспектр», Москва) в концентрации 1 мкг/мл. Интенсивность пролиферации оценивали через 72 ч по включению 3H-тимидина (1 мкКu/лунку), вносимого за 18 ч до окончания культивирования.
Относительное содержание CD4+CD25+ и CD4+CD25high Т-клеток определяли методом проточной цитофлюориметрии на лазерном клеточном сортере-анализаторе FACSCalibur (Becton Dickinson, США) с использованием фикоэретрин (РЕ)-меченных анти-CD4 ( «Сорбент», Москва) и FITC-меченных анти-CD25 моноклональных антител (BD PharMingen, США). Для оценки экспрессии внутриклеточного транскрипционного белка FOXP3 в CD4+ и CD8+Т-клетках, а также перфорина (Pfr) в CD8+Т-лимфоцитах МНК обрабатывали FITC-меченными aнти-CD4 или анти-CD8-антителами («Сорбент», Москва). Пермеаби-лизацию клеток проводили с использованием 0,2 % раствора Твин-20, после чего клетки культивировали 30 мин с PE-меченными анти-FOXP3-антителами (eBioscience, США) или с РЕ-меченными анти-Pfr-антителами (BD PharMingen, США). Для оценки CD4+FOXP3+CD25+ клеток использовали PerCP-меченные анти-CD4, РЕ-меченные анти-FOXP3 и FITC-меченные анти-CD25-антитела. Математическую обработку полученных результатов проводили с использованием программы «Statistica 6.0».
Результаты и обсуждение
Известно, что увеличение количества/ак-тивности Treg при онкопатологии приводит к развитию иммунной недостаточности, одним из проявлений которой является угнетение пролиферативного потенциала Т-клеток. Оценка уровня анти-CD3-стимулированной пролиферации Т-клеток показала (табл. 1), что у здоровых доноров этот показатель варьировал от 6162 до 68953 имп/мин, составляя в среднем 32700±6990 имп/мин. Интенсивность анти-CD3-стимулированного ответа у больных РЯ была в 1,5 раза ниже (от 686 до 48271 имп/мин). Таким образом, циркулирующие Т-лимфоциты больных характеризовались признаками угнетения функциональной активности, что может быть связано с накоплением Treg.
Таблица 1
Пролиферативная активность Т-клеток доноров и больных РЯ
|
Стимул |
Уровень пролиферации МНК периферической крови (имп/мин) |
|
|
Доноры (n=15) |
Больные (n=10) |
|
|
0 |
270 ± 50 |
240 ± 40 |
|
Анти-CD3 |
32700 ± 6990 |
22110 ± 4570* |
Примечание: * – достоверность различий показателей по сравнению с донорами, рU <0,05 (U – критерий Вилкоксона– Манна–Уитни).
Для анализа Treg в периферической крови на первом этапе была проведена оценка относительного и абсолютного количества Т-клеток с фенотипом CD4+CD25+. У здоровых женщин (рис. 1А) относительное содержание CD4+CD25+ клеток в периферической крови варьировало от 3,0 до 11,0 %, составляя в среднем 5,4 ± 0,5 %, что согласуется с данными литературы [8, 28]. У больных РЯ (рис. 1Б) содержание этих клеток было в 2 раза ниже – 2,7 ± 0,3 % (рU=0,0009) с диапазоном от 1,5 до 4 %. Абсолютное количество CD4+CD25+ клеток в периферической крови больных РЯ составило 46 ± 8 клеток/мкл и было также достоверно меньше, чем у здоровых женщин, – 95 ± 7 клеток/мкл (рU=0,001). В то же время относительное содержание CD4+CD25+ клеток среди лимфоцитов асцитической жидко сти больных (n=5; рис. 1В) в 2 раза превышало содержание таковых в периферической крови.
Известно, что субпопуляция CD4+CD25+ клеток является гетерогенной по интенсивности экспрессии CD25 и функциональной активности. При этом показано, что супрессорной активностью обладают преимущественно CD4 Т-клетки с высокой экспрессией CD25 молекулы – так называемые CD4+CD25high клетки [2].
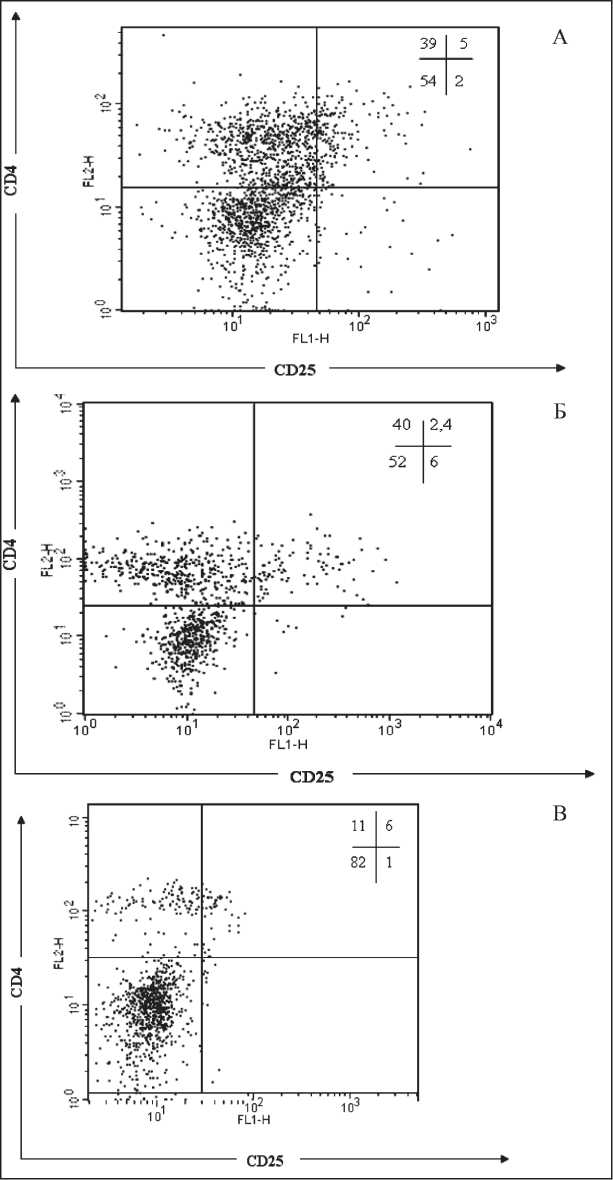
Рис. 1. Индивидуальные значения относительного количества CD4+CD25+ клеток: А – МНК периферической крови донора; Б – МНК периферической крови больной раком яичников;
В – лимфоциты асцитической жидкости этой же пациентки. CD4+CD25+ определяли среди свежевыделенных МНК
Анализ данной субпопуляции при раке яичников ранее не проводился, поэтому параллельно с оценкой CD4+CD25+ Т-клеток, было исследовано относительное содержание CD4 Т-клеток с высокой экспрессией СD25. Относительное количество CD4+CD25high клеток у здоровых доноров (n=15) варьировало от 1 до 2,1 %, в среднем – 1,1 ± 0,1 %. У больных РЯ (n=8) количество CD4+CD25high клеток в периферической крови было достоверно ниже – 0,8 ± 0,3 % (рU=0,02). При этом содержание CD4+CD25high клеток среди лимфоцитов асцитической жидкости почти в 2 раза превышало таковое в периферической крови – 1,4 ± 0,2 % (рU=0,05).
Полученные результаты свидетельствуют о возможном перераспределении популяции CD4+CD25+ у больных РЯ и согласуются с данными Curiel et al., которые впервые продемонстрировали накопление CD4+CD25+ клеток в асцитической жидкости женщин с РЯ [7]. Авторы показали, что направленная миграция CD4+CD25+ клеток в асцитическую жидкость обусловлена взаимодействием хемокина CCL22, продуцируемого опухолевыми клетками и макрофагами, с его рецептором CCR4 на поверхности естественных регуляторных Т-клеток [22]. Выявленное нами более высокое содержание CD4+CD25high клеток в асцитической жидкости по сравнению с периферической кровью является еще одним аргументом в пользу миграции естественных Treg в зону опухолевого микроокружения. Кроме того, данный факт позволяет предполагать, что уменьшение доли CD4+CD25+ Т-клеток периферической крови связано не только с уменьшением в циркуляции активированных Т-лимфоцитов (на фоне опухоль-индуцированной иммуносупрессии), но и естественных регуляторных Т-клеток. С другой стороны, учитывая угнетение пролиферативного потенциала Т-лимфоцитов у больных РЯ, можно полагать, что в циркуляции возрастает число других субпопуляций Treg, в том числе характеризующихся низкой экспрессией или отсутствием на поверхности молекулы CD25. Чтобы проверить это предположение, параллельно с оценкой CD4+CD25+ Т-клеток мы исследовали содержание CD4+Т-клеток с внутриклеточной экспрессией FOXP3, являющегося маркером не только естественных, но и индуцированных Treg.
В периферической крови здоровых доноров (n=20) относительное количество CD4+FOXP3+ клеток варьировало от 1 до 8 %, составляя в среднем 3,3 ± 0,5 % (рис. 2). У больных РЯ (n=10) содержание этих клеток колебалось в пределах от 3 до 16 %, и в среднем более чем в два раза превышало показатель у доноров – 8,5 ± 2,4 % (рU=0,01). Абсолютное количество CD4+FOXP3+ клеток у больных РЯ было также повышенным – 15 ± 57 клеток/мкл против 70 ± 19 клеток/мкл у доноров (рU=0,05). Доля CD4+FOXP3+ клеток среди лимфоцитов асцитической жидкости составила в среднем 6,2 ± 0,9 % (n=5). Таким образом, относительное содержание CD4+FOXP3+ клеток среди лимфоцитов периферической крови и асцитической жидкости больных было сходным и значимо превышало количество этих клеток в периферической крови здоровых женщин.
Увеличение относительного количества CD4+FOXP3+ клеток на фоне снижения доли CD4+CD25+ клеток свидетельствует, что, по-видимому, не все СD4+FOXP3+ клетки несут на своей поверхности молекулу CD25. Действительно, проведенный в отдельной серии экспериментов анализ экспрессии молекулы CD25 в популяции CD4+FOXP3+ клеток периферической крови больных РЯ показал, что лишь небольшая часть этих клеток несла маркер CD25, в то
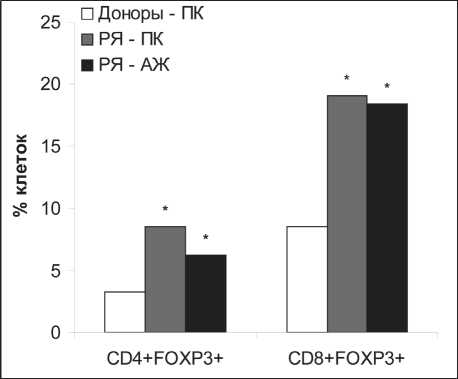
Рис. 2. Относительное количество CD4+FOXP3+ и CD8+FOXP3+ клеток у доноров и больных раком яичников. Примечание: ПК – периферическая кровь, АЖ – асцитическая жидкость, * – различия статистически значимы по сравнению с группой здоровых лиц (рU<0,05)
время как большинство СD4+FOXP3+Т-клеток не экспрессировали CD25 и имели фенотип СD4+FOXP3+CD25- (данные не представлены). Полученные результаты указывают на то, что CD4+CD25+ и CD4+FOXP3+ клетки у больных РЯ являются, по-видимому, различными типами регуляторных Т-клеток. Учитывая, что молекула CD25 является неотъемлемой принадлежностью естественных, но не индуцированных Treg, можно полагать, что у больных РЯ наблюдается увеличение обеих этих субпопуляций. При этом если количество естественных регуляторных CD4+CD25+ Т-клеток возрастает преимущественно в асцитической жидкости (очевидно, за счет их миграции в зону опухолевого микроокружения), то увеличение индуцированных CD4+FOXP3+CD25- Treg регистрируется как в зоне опухолевого роста, так и в циркуляции, то есть имеет более генерализованный характер.
Следует отметить, что, помимо CD4+ Treg, супрессорной способностью обладают и CD8+Т-клетки, экспрессирующие транскрипционный фактор FOXP3+ [5]. Данная популяция клеток является малоизученной и практически не охарактеризованной при онкопатологии. Исследование CD8+FOXP3+ клеток (рис. 2) показало, что в периферической крови относительное количество этих клеток у больных РЯ было повышено более чем в два раза – 19,1 ± 4,4 % против 8,5 ± 1,9 % у доноров (рU=0,04). Такое же количество CD8+FOXP3+ клеток выявлялось среди лимфоцитов асцитической жидкости – 18,4 ± 5,8 %. Увеличение популяции CD8+FOXP3+ было недавно опиисано Y. Kiniwa et al. при раке предстательной железы [13]. Кроме того, S. Wei et al. обнаружили увеличение CD8+Treg, продуцирующих IL-10, в асцитической жидкости больных РЯ [23]. Однако авторы не исследовали экспрессию FOXP3 в указанной популяции клеток.
Изучение регуляторных Т-клеток при онкопатологии представляет особый интерес в том случае, когда данная популяция анализируется в контексте баланса Treg и цитотоксических Т-лимфоцитов (ЦТЛ), играющих важную роль в противоопухолевом ответе [21]. Поскольку одним из механизмов цитотоксической активности ЦТЛ является экспрессия перфорина, была проведена оценка количества CD8+Т-клеток с внутриклеточной экспрессией перфорина. Относительное содержание CD8+Pfr+ клеток в периферической крови здоровых женщин (n=15) и больных РЯ (n=7) было сходным и составляло в среднем 11,1 ± 1,2 % и 11,9 ± 3,8% соответственно. Содержание CD8+Pfr+Т-клеток среди лимфоцитов АЖ было таким же, как в периферической крови – 9,6 ± 2,6 % (n=5). В то же время абсолютное количество CD8+Pfr+ клеток у больных РЯ было достоверно ниже, чем у доноров, – 78 ± 30 и 208 ± 33 клеток/мкл соответственно (PU=0,04). В результате, если у женщин контрольной группы соотношение абсолютного количества ЦТЛ и Treg варьировало от 0,9 до 17, составляя в среднем 5,7 ± 2,0 (n=8), то у больных РЯ этот показатель был снижен в 14 раз – 0,4 ± 0,1 (n=8).
Суммируя полученные данные, можно заключить, что при раке яичников наблюдается увеличение различных субпопуляций Treg. Накопление естественных регуляторных Т-клеток (CD4+CD25+/CD4+CD25high) выявляется преимущественно в зоне опухолевого роста, тогда как увеличение CD4+FOXP3+ и CD8+FOXP3+ регуляторных Т-клеток имеет более генерализованный характер и регистрируется как в периферической крови, так и в асцитической жидкости больных РЯ. При этом отмечается выраженное снижение баланса соотношения ЦТЛ и CD4+FOXP3+ Treg, что может способствовать опухолевому росту.
