Хищные млекопитающие как разорители гнёзд обыкновенного зимородка

Автор: Котюков Юрий Валентинович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
Прослежена судьба 1352 нор обыкновенного зимородка, из которых 253 (18,7%) были разорены дикими хищными млекопитающими. Лисицей уничтожено более 73% разорённых дикими Carnivora гнёзд, представителями сем. Куньи - 2,4%. Частота разорения гнёзд зимородка лисицей не зависит ни от плотности населения зимородка, ни от численности лисицы, ни от обилия её основного корма - мелких млекопитающих. Разорение лисицей гнёзд зимородка обусловлено обилием и доступностью других кормовых объектов.
Обыкновенный зимородок, гнездование, обыкновенная лисица, пищевые объекты, разорение гнёзд
Короткий адрес: https://sciup.org/146116664
IDR: 146116664 | УДК: 591.5:598.278:599.742.1:599.742.4
Carnivorous mammals as predators on common kingfisher nests
We traced the fate of the 1352 burrows of the common kingfisher, of which 253 (18,7%) were destroyed by wild predatory mammals. Red fox was responsible for the destruction of more than 73% nests destroyed by wild carnivorans; mustelids destroyed 2,4% of nests. Rate of the nest predation depends neither on the densisty of the kingfishers’ population, nor on the number of red foxes, nor on the number of micromammals as food items for the fox. Availability of the other food items is related to the nest predation by the red fox.
Текст научной статьи Хищные млекопитающие как разорители гнёзд обыкновенного зимородка
Введение. Репродуктивные потери наиболее важный регулятор численности популяций птиц. Из множества факторов вызывающих частичную или полную потерю кладок или выводков есть два, действие которых постоянно почти в любой популяции и в любой сезон размножения: хищничество и эмбриональная смертность (Паевский, 1985). Практически все виды птиц являются объектом нападения хищников, по крайней мере, на стадии яйца или птенца (Newton, 1998). Тем не менее, разорение гнёзд, частным случаем которого является хищничество, редко отмечается у крупных видов птиц и у птиц гнездящихся колониально (Ricklefs, 1969). У мелких видов птиц хищники разоряют в среднем 30-40% гнёзд, а у отдельных видов бывает разорено более 80% гнёзд (Ковшарь, 1981; Martin, 1993). Наименее всего подвержены разорению гнёзда тех видов, которые гнездятся в норах и дуплах (Ковшарь, 1981; Martin, Li, 1992). Вместе с тем частота разорения гнёзд может быть обусловлена локальной плотностью населения птиц и численностью хищников (Schmidt, Whelan, 1999; Ackerman et al., 2004) и многими другими факторами.
В Окском заповеднике обыкновенный зимородок Alcedo atthis L. является обычным гнездящимся видом. Численность гнездящихся птиц и число жилых нор подвержены значительным (3-10-кратным)
межгодовым колебаниям. Практически ежегодно часть нор зимородка погибает по различным причинам, при этом значительную долю составляют норы, погибшие в результате разорения. В качестве обычных разорителей гнёзд обыкновенного зимородка в Окском заповеднике установлены представители сем Псовые (Canidae): обыкновенная лисица Vulpes vulpes L., енотовидная собака Nyctereutes procyonoides Gray, домашняя собака Cants lupus familiaris L. и сем Куньи (Mustelidae): горностай Mustela erminea L., лесная куница Martes martes L. Цель настоящей работы - обобщить случаи разорения хищными млекопитающими нор обыкновенного зимородка в Окском заповеднике и установить факторы определяющие частоту разорения.
Материал и методика. Исследования проводили в весеннелетние сезоны 1976-2012 гг. на участке р. Пра от западной границы Окского заповедника до устья протяжённостью 54 км (контрольный участок) и на прилегающих участках рек Пра и Ока. Учёт и картирование нор A. atthis проводили во время лодочных маршрутов: на контрольном участке р. Пра до 12 раз в течение сезона, за его пределами - до 4 раз. При каждом посещении норы отмечали её сохранность и производили другую необходимую работу, используя известную методику (Гуревич и др., 1978). При обнаружении погибшей норы пытались определить причину её гибели. В тех случаях, когда нора была разорена, пытались определить вид или группу (семейство) млекопитающих разоривших гнездо. Для этого использовали следующие признаки: характер разорения норы, отпечатки лап, клочки шерсти, экскременты и т.п., оставленные у разорённой норы. Всего за годы исследования на контрольном участке р. Пра найдены 1352 жилых норы зимородка. В 517 случаях гнездование было неуспешным, в том числе 264 гнезда погибли по вине млекопитающих. Так как лисица является основным разорителем гнёзд зимородка, то велика вероятность того, что увеличение численности хищника отрицательно сказывается на успешности размножения зимородка. При статистическом анализе в качестве переменной, характеризующей плотность населения V. vulpes, мы использовали сведения о числе выводков, зарегистрированных на территории Центрального лесничества Окского заповедника и об общей численности по данным зимнего маршрутного учёта (ЗМУ). Эти сведения были почерпнуты из статьи Лавровского (2005) и ежегодников «Летопись природы Окского заповедника». Из «Летописи природы» были также взяты сведения о численности мелких млекопитающих. Учёт мелких млекопитающих с помощью давилок Геро проводили два раза в год на 3 пробных площадках: в пойме Пры, в пойме Оки и на надпойменной террасе, по методике, описанной Формозовым (1937). Показатель обилия мелких млекопитающих - число особей на 100 ловушко-суток (ос./100 л-с).
Статистическую обработку данных проводили с помощью универсального статистического пакета STADIA 8.0/prof, разработанного Кулаичевым (2006). При обработке количественных данных вычисляли параметрический коэффициент корреляции Пирсона (г), коэффициент ранговой корреляции Спирмена (R), коэффициент конкордации Кенделла (1), а также критерии Краскела-Уоллиса (Н) и Джонкхиера (J). Предварительно каждую выборку проверяли на соответствие её распределения нормальному закону.
Результаты и обсуждение. На контрольном участке р. Пра прослежена судьба 1352 жилых нор обыкновенного зимородка - от 5 до 79 нор в течение сезона. Частичные потери - гибель или исчезновение отдельных яиц или птенцов в некоторых гнёздах — наблюдаются постоянно. Вместе с тем практически ежегодно, за исключением сезона 2009 г., отмечаются норы, в которых установлены полная гибель или исчезновение потомства. Из 517 неуспешных нор 40 (7,7%) погибли по вине человека и чуть более половины по вине диких и домашних животных (рис. 1). Вероятно также, что часть гнёзд отнесённых в категорию «прочее» (рис. 1) погибла в результате нападения диких животных. Чаще всего норы зимородка разоряли лисица и енотовидная собака, значительно реже домашняя собака и представители сем. Mustelidae - горностай и лесная куница (рис. 2). Другие млекопитающие становились виновниками гибели нор зимородка исключительно редко.
Одну нору раздавила домашняя корова Bos taurus taurus L. и в 3 случаях норы погибли после того, как были разрыты кротом Talpa europaea L., водяной полёвкой Arvicola terrestris L. и кабаном Sus scrofa L. Раскапывая норы зимородка V. vulpes и N. procyonoides поедают яйца и птенцов; изредка их добычей становится взрослая птица. Не всегда удаётся установить видовую принадлежность хищника, но, судя по бесспорным находкам, из этих двух видов на долю лисицы приходится не менее 75-80% случаев разорения гнёзд зимородка. В отличие от других представителей сем. Canidae домашняя собака не поедает содержимое разрытой норы и зачастую раскапывает её лишь частично, вынуждая птиц бросить гнездо (7 случаев). Горностай, как правило, проникает в гнездо через леток норы, съедает яйца, убивает птенцов и взрослую птицу, оставляя при этом характерные следы (4 случая). Лесная куница, в отличие от горностая, раскапывает сверху расположенную близко к поверхности земли нору (единственный достоверно установленный случай) и через небольшое отверстие достаёт содержимое гнездовой камеры. В одном случае, в нору зимородка, устроенную в грунте среди корней вывороченного ветром дерева, кто-то из представителей сем. Mustelidae (горностай или лесная куница) проник через заднюю стенку гнездовой камеры и уничтожил полную кладку.
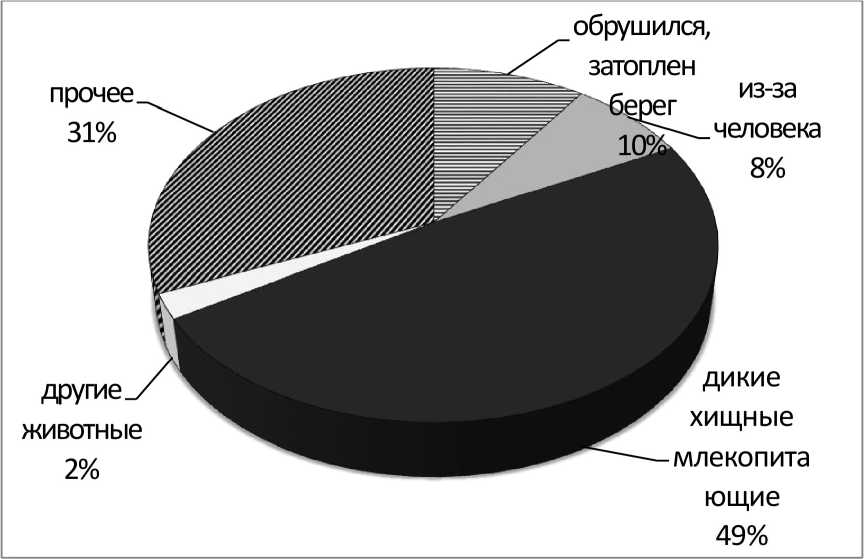
Рис. 1 . Причины гибели гнёзд обыкновенного зимородка.
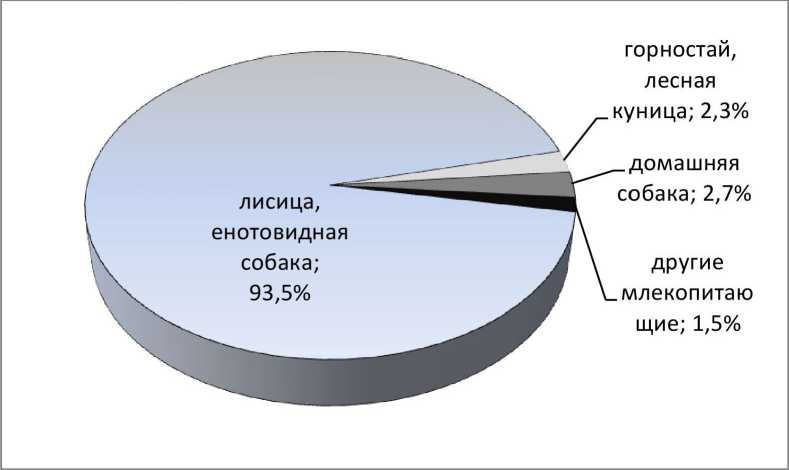
Рис. 2 . Видовой состав млекопитающих и степень их участия в разорении гнёзд обыкновенного зимородка.
Таким образом, лисицей уничтожено не менее 185 гнёзд зимородка, что составляет 71,1% гнёзд разорённых Carnivora (N=260). Можно предположить, что в годы высокой численности V. wipes увеличивается и число нор зимородка, разорённых хищниками. Однако статистические расчёты показали, что и численность по данным ЗМУ, и число выводковых нор лисицы не коррелируют с числом разорённых гнёзд зимородка и их долей среди всех гнёзд: г<0,04 при р>0,81; t<0,124 при р>0,16; R<0,175 при р>0,16.
В рационе У. vulpes в Окском заповеднике во все сезоны года преобладают мышевидные грызуны (Бородина, 1940; Лавровский, 2005). Численность грызунов также, как и других мелких млекопитающих сем. Soricidae, изменяется в значительных пределах даже в течение коротких временных интервалов. Например, в Окском заповеднике обилие мелких млекопитающих может увеличиться от весны к осени в 20-30 раз и вновь сократиться в 50 раз к следующей весне. Такие значительные колебания численности основных пищевых объектов должны вынуждать лисицу искать другие корма, одним из которых в весенне-летний период могут стать яйца и птенцы зимородка. Тем не менее, корреляционный анализ показал, что число разорённых гнёзд зимородка и их доля среди жилых гнёзд варьируют независимо от обилия мелких млекопитающих.
Испробовав все возможные варианты компоновки данных (численность мелких млекопитающих по отдельным площадкам и сезонам, совокупная за год по площадкам и т.д.) мы обнаружили ничтожно малые по абсолютной величине значения коэффициентов t<0,135 при р>0,12 и R<0,167 при р>0,16. Условный показатель прироста численности мелких млекопитающих, вычисляемый как разность между осенней и весенней численностью одного года, также не оказывал влияния на количество разорённых гнёзд зимородка (рис. 31
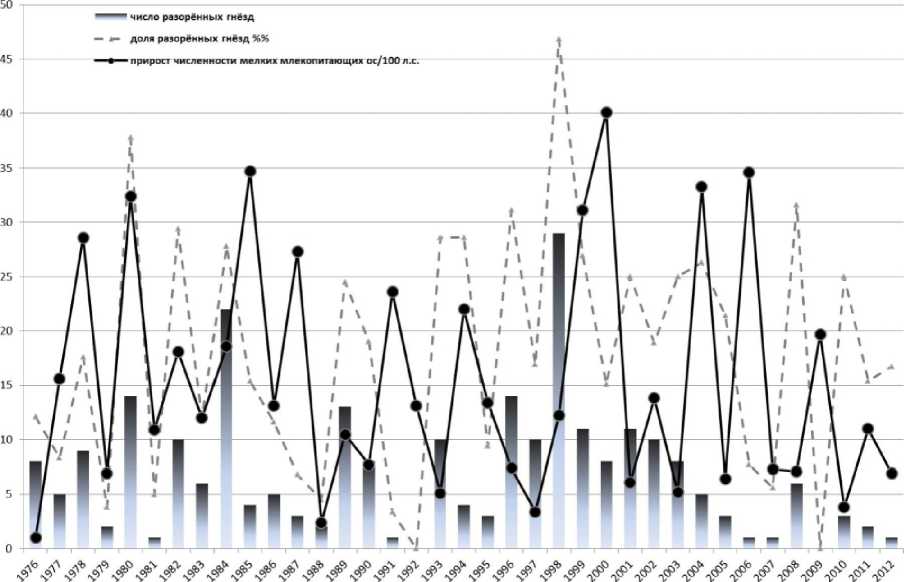
Р и с . 3 . Динамика численности мелких млекопитающих и разорение гнёзд зимородка дикими Carnivora.
Частота разорения гнёзд зачастую обусловлена плотностью населения потенциальных объектов нападения хищников. Как показали эксперименты с искусственными моделями гнёзд, степень разорения при увеличении плотности расположения гнёзд может расти (Gunnarsson, Elmberg, 2008) или оставаться постоянной (Ackerman et al., 2004). В других экспериментах положительная связь плотности расположения гнёзд и хищничества наблюдалась только в определённых биотопах или при определённом способе размещения гнёзд потенциальной жертвы (Schmidt, Whelan, 1999). На фоне значительных колебаний численности зимородка в Окском заповеднике почти ежегодно (исключая сезоны 1992 и 2009 гг.) дикие Carnivora разоряют 1-29 нор, которые составляют 3,4-48,6% жилых нор (рис. 3). Оказалось, что интенсивность разорения нор не зависит от их общего количества на контрольном участке (г=0,152 при р=0,63; t=0,063 при р=0,29; R=0,107 при р=0,26). Проверка статистической зависимости методом рангового однофакторного анализа также показала отсутствие таковой (Н=2,948, р=0,399, df=3; 1=223, р=0,483, df=4, 37). Вопреки ожиданиям, частота разорения нор зимородка не зависит от обилия ни самой жертвы, ни хищника, ни от обилия основного корма У. vulpes - мелких млекопитающих.
В сообществах Carnivora, специализирующихся на разорении птичьих гнёзд, при исчезновении или сокращении численности одного из видов хищников его роль исполняют другие виды. Например, в одном исследовании успешности размножения чибиса Vanellus vanellus L. при низкой численности лисицы Mustelidae чаще разоряли его гнёзда (Bellebaum, Bock, 2008). В сезоны низкой численности лисицы в Окском заповеднике в 1976 г. (25 особей, 3 выводка) и 1977 г. (9 особей, 2 выводка) дикие Canidae разорили соответственно 12,1 и 8,3% гнёзд зимородка (рис. 3). В 1993 г., когда по данным ЗМУ численность У. vulpes составляла 83 особи и найдено 7 выводковых нор, дикие Canidae разорили 7 из 35 (20%) жилых нор зимородка, а 3 норы (8,6%) были разорены горностаем. В сезоне 1992 г., когда в заповеднике было найдено 20 выводков лисицы, и численность её по данным ЗМУ составила 45 особей, случаи разорения гнёзд зимородка хищными млекопитающими не зарегистрированы (рис. 3). В другие сезоны, когда отмечены единичные случаи разорения Mustelidae нор зимородка (1989, 2003, 2004 гг.), численность У. vulpes была сравнительно высокой. Не установлено также увеличение частоты разорения гнёзд зимородка енотовидной собакой в годы низкой численности лисицы. Потенциальным разорителем гнёзд зимородка является барсук Meles meles L., однако, такие случаи в заповеднике не отмечены.
Как правило, выводковые норы лисицы в Окском заповеднике приурочены к повышенным участкам местности - дюнным всхолмлениям и надпойменным террасам. Так как площадь территории одной пары V. vulpes составляет в среднем 624 га (Лавровский, 2005), то гипотетически все лисицы, поселившиеся в 1-3 км от русла Пры, представляют потенциальную опасность для гнёзд зимородка. В выделенной таким образом полосе, охватывающей долину Пры и прилежащие повышенные участки местности, размещение выводков V. vulpes неравномерно. По результатам специальных исследований (Уваров, Лавровский, 2005) и данным Летописи природы Окского заповедника всю полосу можно разделить на 3 примерно равных по длине отрезка, различающихся частотой и плотностью заселения их лисицей. В верхнем по течению отрезке лисица селится постоянно и практически ежегодно многие гнёзда зимородка бывают ею разорены. В среднем отрезке V. vulpes поселяется реже и в меньшем числе, а гнёзда зимородков здесь разоряются регулярно (20 сезонов из 35). В нижнем по течению участке выводковые норы V. vulpes регистрировали редко, а разорение гнёзд A. atthis лисицей отмечено только в течение б из 35 сезонов. Из этого следует, что при увеличении численности выводков V. vulpes в долине Пры вероятность разорения гнёзд A. atthis растёт. Тем не менее, некоторые данные, например, отсутствие разорённых Carnivora нор зимородка в 1992 г. при очень высокой численности выводков V. vulpes противоречат этой тенденции.
Из изложенного выше следует, что плотность населения зимородка, численность лисицы и обилие мелких млекопитающих не оказывают заметного влияния друг на друга, а частота разорения нор зимородка не зависит от величины этих показателей. Поэтому трудно не согласиться с мнением некоторых исследователей (Schmidt, Whelan, 1999; Ackerman et ah, 2004) о том, что отсутствие зависимости хищничества от плотности населения жертвы и самого хищника отражает распределение и возможность добычи хищником альтернативных кормов.
Заключение. В результате исследований установлено, что в течение 37 сезонов размножения (1976-2012 гг.) дикие хищные млекопитающие разорили 253 (18,7%, N=1352) жилые норы обыкновенного зимородка. Наибольшее число разорённых нор (>185) приходится на долю V. vulpes; ей уничтожено более 73% разорённых дикими Carnivora и не менее 35% всех погибших по разным причинам гнёзд A. atthis.
В течение двух сезонов размножения зимородка случаи разорения его гнёзд дикими Carnivora не установлены, в другие сезоны они уничтожали 3,4-46,8% гнёзд. Плотность населения зимородка не оказывает статистически значимого (р>0,26) влияния на частоту разорения его гнёзд хищными млекопитающими. Доля разорённых гнёзд изменяется ежегодно независимо от (а) общей численности V. vulpes (данные ЗМУ), (б) численности размножающихся пар V. vulpes (количество выводковых нор) и (в) обилия основного корма V. vulpes - мелких млекопитающих.
Другие виды хищных млекопитающих, в частности, представители Mustelidae редко разоряют гнёзда зимородка, и доля гнёзд, разорённых ими, не увеличивается в годы низкой численности лисицы. В состав кормов лисицы, кроме содержимого нор зимородка и мелких млекопитающих, входит широкий спектр животных и растительных объектов. Обилие и доступность других пищевых объектов, в первую очередь, определяют частоту разорения лисицей гнёзд зимородка.
CARNIVOROUS MAMMALS AS PREDATORS
ON COMMON KINGFISHER NESTS
Yu. V. Kotyukov
Котюков Ю.В. Хищные млекопитающие ка кразорители гнезд обыкновенного зимородка / Ю.В. Котюков // Вести. ТвГУ. Сер.: Биология и экология. 2016. № 4. С. 121-129.
Список литературы Хищные млекопитающие как разорители гнёзд обыкновенного зимородка
- Бородина М.Н. 1940. Материалы по питанию лисицы в Окском заповеднике//Тр. Окского заповедника. М.: Гудок. Вып. 1. С. 150-173.
- Гуревич Я.Д., Нумеров А.Д., Алленова Л.В., Крюкова О.В., Лысенко Я.П. 1978. Некоторые данные по экологии и продуктивности зимородка//Тр. Окского заповедника. Рязань: Московский рабочий. Вып. 14. С. 210-216.
- Ковшарь А.Ф. 1981. Особенности размножения птиц в субвысокогорье (на материале Passeriformes в Тянь-Шане). Алма-Ата: Наука. 260 с.
- Кулаичев А.П. 2006. Методы и средства комплексного анализа данных. М.: ФОРУМ: ИНФРА-М. 512 с.
- Лавровский В.В. 2005. Особенности экологии лисицы в Окском заповеднике//Роль заповедников лесной зоны в сохранении и изучении биологического разнообразия европейской части России (Материалы научно-практической конференции, посвящённой 70-летию Окского государственного природного биосферного заповедника) Тр. Окского заповедника. Рязань. Вып. 24. С. 183-224.
- Паевский В.А. 1985. Демография птиц (Тр. Зоол. ин-та; Т. 125). Л.: Наука. 285 с.
- Уваров Н.В., Лавровский В.В. 2005. Размещение выводков лисицы Vulpes vulpes в Окском заповеднике//Роль заповедников лесной зоны в сохранении и изучении биологического разнообразия европейской части России (Материалы научно-практической конференции, посвящённой 70-летию Окского государственного природного биосферного заповедника) Тр. Окского заповедника. Рязань. Вып. 24. С. 165-172.
- Формозов А.Н. 1937. Программа и методика работы наблюдательных пунктов по учету мышевидных грызунов в целях прогноза их массового появления//Учен. записки МГУ. Сер. биол. М. Вып. 11. С. 37-119.
- Ackerman J.T., Blackmer A.L., Eadie J.M. 2004. Is predation on waterfowl nests density dependent? -Tests at three spatial scales//Oikos V. 107. № 1. P. 128-140.
- Bellebaum J., Bock C. 2008. Influence of ground predators and water levels on Lapwing Vanellus vanellus breeding success in two continental wetlands//J. Ornithol. V. 150 № 1. P. 221-230.
- Gunnarsson G., Elmberg J. 2008. Density-dependent nest predation -an experiment with simulated mallard nests in contrasting landscapes//Ibis V. 150. № 2. P. 259 -269.
- Martin T.E. 1993. Nest predation among vegetation layers and habitat types: revising the dogmas//Am. Nat. V. 141. № 6. P. 897-913.
- Martin T.E., Li P. 1992. Life history traits of open-vs. cavity-nesting birds//Ecology V. 73. № 2. P. 579-592.
- Newton I. 1998. Population limitation in birds. London: Academic Press. 597 p.
- Ricklefs R.E. 1969. An analysis of nesting mortality in birds//Smithson. Contrib. Zool. № 9. P. 1-48.
- Schmidt K.A., Whelan C.J. 1999. Nest predation on woodland songbirds: when is nest predation density dependent?//Oikos V. 87. № 1. P. 65-74.
