Хронобиологические аспекты оптимизации пестицидной нагрузки в насаждениях ивы корзиночной (Salix viminalis L.) интенсивного типа
 интенсивного типа")
Автор: Афонин А.А.
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология микроорганизмов, растений
Статья в выпуске: 2, 2019 года.
Бесплатный доступ
Дана характеристика ивы корзиночной с утилитарной точки зрения. Обоснована необходимость применения пестицидов с пролонгированным действием в насаждениях интенсивного типа. Для снижения пестицидной нагрузки предложено планировать агролесотехнические мероприятия с учетом сезонной динамики нарастания однолетних побегов. Цель - выявление закономерностей сезонной динамики суточного прироста однолетних побегов ивы корзиночной. Материал: модельная инбредная популяция ивы корзиночной. Методы: селекционные, структурно-морфологические, методы гармонического анализа. Установлено, что сезонная динамика суточного прироста побегов Δ L(t) носит циклический характер. Выявлена трехуровневая структура сезонных биоритмов суточного прироста побегов. Вклад низших гармоник с периодом колебаний 32…96 суток в общую цикличность Δ L(t) составляет 65%. Вклад средних гармоник с периодом колебаний 16…24 суток в общую цикличность Δ L(t) составляет 23%. Вклад высших гармоник с периодом колебаний менее 14 суток в общую цикличность Δ L(t) составляет 6%. Для оптимизации пестицидной нагрузки в насаждениях ивы корзиночной рекомендуется учитывать биоритмы нарастания побегов с периодичностью колебаний порядка 16…24 суток.
Ива корзиночная, хронобиология, биоритмы, суточный прирост, сезонная динамика, ангармонический, квазипериодический, квазициклический
Короткий адрес: https://sciup.org/14117149
IDR: 14117149 | УДК: 582.623.2 | DOI: 10.36906/2311-4444/19-2/06
Chronobiological aspects of pesticide load optimization in Salix viminalis L. plantings of intensive type
The article considers common osier from the utilitarian perspective and addresses the necessity to applyprolonged-action pesticides in the plantings of intensive type. To reduce the pesticide load, it is proposed to schedule agroforestry activities taking into account the seasonal dynamics of annual shoot growth. The study aims to identify patterns of seasonal dynamics of daily increment of annual shoots of common osier, using such methods as selection, structural-morphological methods, and harmonic analysis. The object of the study is a model inbred population of common osier. It was found that the seasonal dynamics of daily shoot growth ΔL(t) is cyclic. A three-level structure of seasonal biorhythms of daily shoot growth wasdiscovered. The contribution of lower harmonics with the oscillation period of 32 to 96 days to the total cyclicity ΔL(t) was 65%. The contribution of the medium harmonics with the oscillation period of 16 to 24daysto the total cyclicity ΔL(t) was 23%. The contribution of higher harmonics with the oscillation period of less than 14 days to the total cyclicity ΔL(t) was 6%. To optimize the pesticide load in the plantings of common osier, it is recommended to take into account the biorhythms of shoot growth with a frequency of fluctuations from 16 to 24 days.
Текст научной статьи Хронобиологические аспекты оптимизации пестицидной нагрузки в насаждениях ивы корзиночной (Salix viminalis L.) интенсивного типа
Ива корзиночная, или прутовидная - Salix viminalis L. 1753 - общеизвестный представитель кустарниковых ив секции Vimen Dum. 1825 (Viminella Ser. 1824) (Скворцов 1968; Анциферов 1984). Прут ивы корзиночной традиционно используется для плетения (Керн 1915). В каталоге “Checklist for Cultivars of Salix...” (Kuzovkina 2015) описано 28 культиваров, из них для выращивания на прут рекомендуются десять клонов. Ива корзиночная перспективна для создания энергетических плантаций (Фучи-ло и др. 2009; Афонин, Булавинцева 2011b; Karp et al. 2011; Nemeth et al. 2013). Ива корзиночная может произрастать в самых неблагоприятных условиях (Miko et al. 2014), поэтому она широко используется как биоремедиатор почв (Zhao, Yang 2017), загрязненных тяжелыми металлами (Hammer et al. 2003; Mleczek et al. 2010; Jama, Nowak 2012). В коре ивы корзиночной содержится целый комплекс биологи чески активных компонентов (Созинов, Кузьмичева 2016), например, полифенолы - антиоксиданты (Ilnicka et al. 2014). Ива корзиночная характеризуется высоким уровнем географической и внутрипопуляционной изменчивости (Сукачев 1934; Скворцов 1968; Trybush et al. 2012; Berlin et al. 2014). Высокий уровень внутривидового разнообразия и способность к образованию межвидовых гибридов способствуют успешной селекции рассматриваемого вида (Царев и др. 2003; Ronnberg-Wastljung et al. 2003; Fuchylo et al. 2016). При проектировании устойчивых высокопродуктивных насаждений следует учитывать взаимодействия «генотип-среда» (Ronnberg-Wastljung 2001; Fabio et al. 2017; Фучило и др. 2018).
Для повышения устойчивости и продуктивности ивовых насаждений необходимо обеспечить защиту растений от вредителей. Листья ив поражаются фитофагами, например,
Phratora (Phyllodecta) vulgatissima (Sage, Tucker 1998). Качество прута снижается при поражении побеговыми галлицами рода Dasineura (=Rhabdophaga) (Ollerstam, Larson 2003). Реальным способом борьбы с патогенами и вредителями в агробиоценозах является использование химических средств защиты растений (Ганиев, Недорезков 2006). Современный ассортимент пестицидов достаточно обширен (Государственный каталог... 2017). Надежную защиту от стеблевых вредителей создают современные инсектициды с глубинным, трансламинарным и системным эффектом, а также препаративные формы традиционных пестицидов с пролонгированным действием (Литвишко 2018).
Агролесотехнические мероприятия наиболее эффективны, если они приурочиваются к переломным этапам развития (Кренке 1940). Режимы обработки ивовых плантаций фунгицидами и инсектоакарицидами должны разрабатываться с учетом биоритмов нарастания годичных побегов, что позволит снизить пестицидную нагрузку на агробиоценозы (Афонин, Булавинцева 2011а). Однако сезонной динамике морфо-физиологических признаков ивы корзиночной посвящены лишь немногие исследования (Иванова, Костюченко 2011; Кузьмичева 2015; Созинов, Кузьмичева 2016).
Цель настоящего исследования - выявление закономерностей сезонной динамики суточного прироста однолетних побегов ивы кор-зиночпой для разработки хронобиологических основ оптимизации пестицидной нагрузки в насаждениях интенсивного типа.
Материал и методы
Объект исследования - модельная популяция ивы корзиночной, созданная методом регулярного инбридинга в культуре (салицетум Брянского государственного университета; серые лесные почвы на лёссовидном суглинке с меловыми подстилающими породами; ТЛУ — D3). ’
В качестве первичного генетического материала (Fo) использовались сеянцы, выращенные из семян, собранных весной 2001 г. с маточной особи 5. viminalis. Место сбора - центральная пойма р. Десны в черте г. Брянска (координаты 53.220569, 34.371874), тип растительности Salicetum triandro-viminalis (Тх. 1931) Lohm. 1952. В 2004 г. две наиболее продуктивные особи Fo были скрещены между собой. Наивысшую продуктивность в их потомстве — инбредной семье Fi - показали две пестичные особи. В последующие годы путем их беккрос-сирования с отцовским клоном Fo было полу чено пять инбредных семей F уЬ, из которых были сохранены две: vi 2-х (2007 г.) и vi 4-х (2009 г.). Наиболее продуктивные сибсы инбредных семей были использованы для дальнейшего изучения. Пять из них —vz'21, vz 22, vi 41, vi 46, vi 42 — составили модельную популяцию.
Материал для данного исследования собирался в течение вегетационного периода 2014 г. Начиная с 11 мая (начало активной вегетации) каждые 5... 6 суток с точностью ± 1 см производили замеры длины L наиболее сильных побегов. Дата последнего измерения -10 августа (завершение активного роста побегов). Для последующего анализа использовались три особи (vz 22, vi 46,vz 47), у которых были получены данные по трем наиболее сильным побегам, завершившим свое развитие без видимых повреждений. Все побеги были пронумерованы в порядке уменьшения их длины в соответствии с номерами особей: vz'221, vz 222, vz 223, vi 461, vz 462, vi 463, vi 471, vi 472, vi 473. Каждому очередному наблюдению i был присвоен порядковый номер k (1, 2, З..Д,). Все даты, в которые проводились наблюдения (/), были пронумерованы по порядку (0, 4, 8...Ц).
Вычисление рядов сезонной динамики суточного прироста побегов производили по алгоритму, описанному нами ранее (Афонин 2019). ' " '
На основании фактических исходных данных (actual data) были рассчитаны эмпирические ряды NL(t)acX;
Д^ЮасТ = где AL(7)act - текущий суточный прирост побега в интервале между предыдущим и последующим наблюдениями (см/сут); Lk+X и Lk-\ — длина побега (см) в предыдущем и последующем наблюдении; tk+\ и tk_x - предыдущая и последующая пронумерованные даты наблюдений.
Для выявления сезонного тренда NL(t) использовалась линейная аппроксимация эмпирических рядов:
Д1щп = Д10 + ^t, где AL iin - ожидаемый суточный прирост побегов в расчетный день наблюдений t; \L 0 - ожидаемый суточный прирост побегов при t = 0; 9 — коэффициент, определяющий тенденцию сезонной динамики AL; для каждого уравнения вычислялся коэффициент детерминации R2, показывающий, насколько адекватно эмпирический ряд аппроксимируется вычисленным рядом.
Для каждого побега были вычислены эмпирические ряды динамики отклонений факти- ческих значений суточного прироста побегов от расчетного сезонного тренда: iU'COact = Д^СОасГ - ALtolin, где dLact — фактическое отклонение фактического значения суточного прироста побегов ^L act от значения ААь„, вычисленного по уравнению линейной регрессии для каждого t.
На основании эмпирических рядов динамики dL(t)act были вычислены соответствующие средние ряды для каждой особи, а для всех побегов - средний агрегированный ряд. Для выявления закономерностей динамики суточного прироста побегов использовался разработанный нами алгоритм гармонического анализа (Афонин, Зайцев 2016). Эмпирические ряды динамики dL(t) act каждого побега были аппроксимированы суммами гармоник h с порядковым номером п;
сдс = У- (Л" ^"^^ + ^ та=1
где dL(t)ca\c — ожидаемое отклонение фактического значения суточного прироста побегов АЛ act от значения АЛИп, вычисленного по уравнению линейной регрессии; Ап — амплитуда колебания; п — порядковый номер гармоники; л -число «пи»; t — пронумерованные даты наблюдений; Т- главный период колебаний, фп — начальная фаза; период колебания каждой гармоники равен т = Т/п.
Достоверность аппроксимации эмпирических рядов каждой гармоникой и суммой гармоник определяли, сравнивая расчетные (calc) и критические (crit) значения F-критерия Фишера на высшем уровне значимости (Р < 0,001). Адекватность аппроксимации эмпирических рядов суммами гармоник оценивали с помощью коэффициента детерминации R". На основании частных рядов динамики был построен агрегированный (средний) ряд сезонной динамики dL(t). Для всех побегов были построены амплитудно-частотные (АЧ) спектрограммы расчетной динамики dL(t)ca\c. Далее для каждой особи были вычислены средние значения амплитуд А и построены соответствующие спектрограммы. На основании визуального анализа АЧ-спектрограмм последовательность гармоник агрегированного (среднего) амплитудно-частотного спектра была разделена на три группы: низшие I, средние m и высшие И. Для каждой группы гармоник вычисляли размах варьирования dL. Для оценки вклада гар моник, различающихся по частоте и амплитуде, в общую сезонную динамику суточного прироста побегов использовали средние квадраты отклонений dL(t) от нуля:
^=Л2+Д2 +s2+£ где D" — средние квадраты отклонений на уровнях а, I, m и И; е - остаточный член ряда.
Результаты и обсуждение
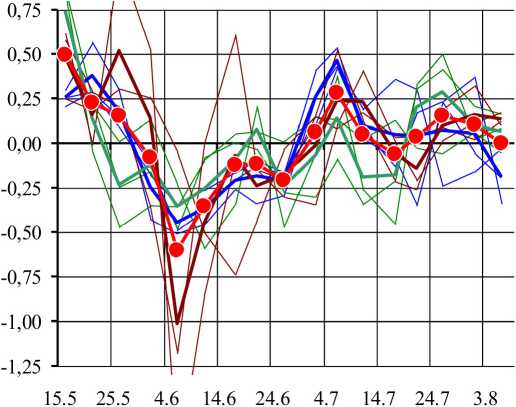
Среднесуточный прирост изученных побегов - vz'221, vi 222, vz 223, vz'461, vz 462, vz 463, vz'471, vz 472, vz 473 - составил 0,66...2,10 см/сут при длине побегов в конце сезона 80...217 см. В большинстве случаев (кроме vi 221 и vi 223) сезонные тренды изменчивости суточного прироста побегов удовлетворительно (R2 = 0,45... 0,80) аппроксимируются уравнениями линейной регрессии. Выявлена тенденция к снижению суточного прироста побегов на протяжении сезона: от АЛНп = 1,5...3,2 см/сут в середине мая до АЛ|in = 0,0... 1,0 см/сут в начале августа. На основании вычисленных рядов линейной регрессии были вычислены эмпирические ряды динамики dL(t)act для каждого побега, средние ряды для каждой особи и агрегированный ряд для всех девяти побегов. Полученные эмпирические ряды показаны в виде соответствующих графиков (рис. 1).
Все ряды динамики dL(t) демонстрируют циклический характер сезонной динамики суточного прироста побегов, который может быть сведен к агрегированному среднему ряду dL(t)a. Этот ряд представляет собой периодическую ангармоническую осциллограмму, на которой выделяются минимумы и максимумы, разделенные периодами 19...21 сут. Размах варьирования dL в агрегированном ряду составляет 1,10 см/сут (от -0,61 до 0,49), при этом средний квадрат отклонений dL(t) от нуля D "act = 0,060. Эмпирические ряды динамики dL(t)ac{ каждого побега были аппроксимированы суммами гармоник h\...h% при основном периоде колебаний Т= 96 сут. В итоге были получены расчетные ряды динамики: dL(t) са]с. Достоверность такой аппроксимации оказалась исключительно высокой: Fcalc = 38,31...213,18 (Fcrit=3,92 при Р< 0,001), R2 = 0,97... 1,00. Таким образом, на генетически выровненном материале, культивируемом в однородных условиях, установлено, что сезонные биоритмы суточного линейного прироста побегов ивы корзиночной с высокой точностью аппроксимируются уравнениями гармонических колебаний. Сравнительный анализ амплитудночастотных спектров позволил оценить вклад гармоник, различающихся по частоте и амплитуде, в общую сезонную динамику суточного прироста побегов (рис. 2).
----221
---222
---223
-461
-462
-463
----471
---472
---473
---22
---46
---47
-•- aggr
Рис. 1. Эмпирические ряды динамики отклонений фактических значений суточного прироста побегов от сезонного тренда
По оси абсцисс -расчетные даты, по оси ординат - отклонения (см/сут). В легенде указаны номера побегов и особей. Приведен агрегированный (aggr) ряд динамики.
-•— dL act
---dLl
--dL m
--dLh
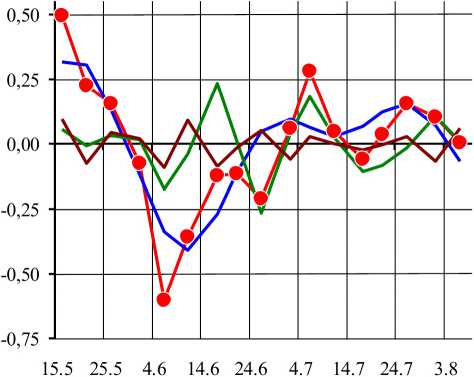
Рис. 2. Вклад гармоник, различающихся по частоте и амплитуде, в общую сезонную динамику суточного прироста побегов
По оси абсцисс — расчетные даты, по оси ординат - отклонения (см/сут). В легенде указаны уровни гармоник. Приведен фактический (act) агрегированный ряд динамики.
В агрегированном АЧС наибольшей средней амплитудой (Л1,5 = 0,236) характеризуется полуторная гармоника h\5 (ti,5 = 64 сут). Для большинства побегов Hi-5 = 0,100...0,400, и только для побега vi 462 Hi_5 = 0,025. Полуторной гармоникой определяется общая цикличность сезонной динамики суточного прироста побегов: глобальный минимум суточного прироста побегов в конце первой половины июня и глобальный максимум в середине июля. Полуторная гармоника окружена первой (т15 = 96 сут) и второй (г1-5 = 48 сут) гармониками со средними амплитудами Ах = 0,083 (от 0,000 до 0,300) и А3 = 0,106 (от 0,025 до 0,200). На большинстве побегов первая и вторая гармоники оказывают незначительное модули рующее влияние на общую цикличность сезонной динамики суточного прироста побегов (за исключением побега vz 462). На втором месте после полуторной гармоники в агрегированном АЧС идет третья гармоника Аз (тз = 32 сут) со средней амплитудой А3 = 0,194 (от 0,025 до 0,450). Эта гармоника оказывает значительное влияние на общую цикличность сезонной динамики суточного прироста побегов, разделяя глобальный июльский максимум на два. Размах варьирования dL в агрегированном ряду низших гармоник А/ составляет 0,72 см/сут (от -0,41 до 0,32). Средний квадрат отклонений dL(t) от нуля в сумме низших гармоник 0^1= 0,039, что составляет 65% от D\ct. Таким образом, длиннопериодические низшие гармо- ники (прежде всего, высокоамплитудные Ai,5 и Аз) определяют общую цикличность сезонной динамики суточного прироста побегов: глобальный минимум в середине июня и два примерно равных максимума в начале и конце ию ля, разделенные относительным минимумом в середине июля.
Среди гармоник с длиной периода менее 30 дней на спектрограмме выделяются три средние гармоники с относительно высокой амплитудой: четвертая
Л4 = 0,075...0,400), пятая
Л4 = 0,050...0,300) и шестая
(г4 = 24 сут;
(т5 ~ 19 сут;
(г6= 16 сут;
Л = 0,000...0,300). Сумма средних гармоник h^...h6 образует квазипериодический ряд с двумя максимумами: 17 июня и 6 июля. Максимум 17 июня проявляется на графике агрегированного ряда dL(t)a в виде выпуклого излома, а максимум 6 июля совпадает с раннеиюльским максимумом. Даты 17 июня и 6 июля разделены интервалом в 19 суток, что соответствует периоду колебаний в агрегированном ряду и периоду колебаний в пятой гармонике. Кроме основных максимумов в сумме средних гармоник А4...Аб выявляются и дополнительные максимумы: 21...31 мая и 26...31 июля. Даты майских максимумов привязаны к определенным особям, а даты позднеиюльских максимумов -к разным побегам. На общую сезонную динамику суточного прироста побегов дополнительные максимумы не оказывают заметного влияния. Размах варьирования dL в агрегированном ряду средних гармоник hm составляет 0,50 см/сут (от -0,27 до 0,23). Средний квадрат отклонений dL(t) от нуля в сумме средних гармоник D"m = 0,014, что составляет 23% от D~act. В целом средние гармоники с периодом колебания 16...24 сут оказывают достаточно сильное модулирующее воздействие на общую цикличность сезонной динамики суточного прироста побегов, определяя периодичность колебаний в середине сезона.
Высшие гармоники А7 и А8 с периодом колебаний 12... 14 сут характеризуются низкими амплитудами (в среднем А = 0,080). Сумма высших гармоник А7...А8 образует квазицикли-ческую последовательность: в первой половине сезона выявляется три слабо выраженных максимума с интервалом в 15 суток, а во второй половине сезона пики практически отсутствуют. Размах варьирования dL в агрегированном ряду высших гармоник hh составляет 0,19 см/сут (от -0,09 до 0,09). Средний квадрат отклонений dL(t) от нуля в сумме высших гармоник Ь1 h = 0,003, что составляет 6% от D~act. Таким образом, высшие низкоамплитудные рующее воздействие на сезонную динамику суточного прироста побегов.
Средний квадрат отклонений dL(t) от нуля, обусловленный остаточной изменчивостью е, равен 0,004, что составляет 7% от D"act. Таким образом, влияние неучтенных факторов на сезонную динамику суточного прироста побегов оказывается незначительным и сопоставимым по значимости с влиянием высших низкоамплитудных гармоник.
Выводы
Анализ биоритмов нарастания однолетних побегов в модельной инбредной популяции ивы корзиночной позволил установить основные хронобиологические закономерности динамики суточного прироста побегов.
-
1. Сезонная динамика суточного прироста побегов носит циклический характер и представляет собой ангармонический агрегированный средний ряд с периодом колебаний 19... 21 сут.
-
2. Эмпирические ряды динамики суточного прироста побегов с высокой точностью аппроксимируются суммами гармоник с периодом колебаний от 12 до 96 сут.
-
3. Низшие гармоники с периодом колебания от 32 до 96 сут определяют общую цикличность сезонной динамики суточного прироста побегов. Их вклад в общую цикличность составляет 65%.
-
4. Средние гармоники с периодом колебания от 16 до 24 сут оказывают сильное модулирующее воздействие на общую цикличность сезонной динамики суточного прироста побегов в середине сезона. Сумма средних гармоник образует квазипериодический ряд с периодом колебаний 19 сут. Их вклад в общую цикличность суточного прироста побегов составляет 23%. ' ‘
-
6. Высшие низкоамплитудные гармоники с периодом колебания менее 14 сут образуют квазициклическую компоненту агрегированного ряда колебаний. Их вклад в общую цикличность суточного прироста побегов составляет 6%. ’ ' '
-
7. Для оптимизации пестицидной нагрузки при использовании химических средств защиты плантаций ивы корзиночной от вредителей и патогенов рекомендуется учитывать биоритмы нарастания побегов с периодичностью колебаний порядка 16...24 сут.
гармоники оказывают незначительное модули
Список литературы Хронобиологические аспекты оптимизации пестицидной нагрузки в насаждениях ивы корзиночной (Salix viminalis L.) интенсивного типа
- Анциферов Г. И. 1984. Ива. М.: Лесная промышленность.
- Афонин А. А. 2019. Структурный анализ ритмов развития однолетних побегов ивы трехтычинковой // Бюллетень науки и практики 5(1), 22-32.
- Афонин А. А., Булавинцева Л. И. 2011a. Генетическая безопасность агробиоценозов // Биология в школе 5, 3-10.
- Афонин А. А., Булавинцева Л. И. 2011b. Эколого-генетические проблемы "зеленой" энергетики // Биология в школе 6, 16-24.
- Афонин А. А., Зайцев С. А. 2016. Цикличность среднесуточного радиального прироста несущих побегов ивы белой (Salix alba L.) в условиях Брянского лесного массива // Известия высших учебных заведений. Лесной журнал 3(351), 66-76.
- Ганиев М. М., Недорезков В. Д. 2006. Химические средства защиты растений. М.: КолосС.
- Государственный каталог пестицидов и агрохимикатов, разрешенных к применению на территории Российской Федерации. 2017. М.: Минсельхоз РФ.
- Иванова Н. А., Костюченко Р. Н. 2011. Эколого-физиологические механизмы адаптации некоторых видов ив в различных условиях обитания на территории Среднего Приобья: Монография. Нижневартовск: Изд-во Нижневарт. гуманит. ун-та.
- Керн Э. Э. Ива, ее значение, разведение и употребление. 1915. Петроград: Тип. Мин-ва Путей Сообщения (Тов-ва И.Н. Кушнерев и К°).
- Кренке Н. П. 1940. Теория циклического старения и омоложения растений и практическое её применение. М.: Огиз-Сельхозгиз.
- Кузьмичева Н. А. 2015. Взаимосвязь морфолого-химических параметров листьев ивы прутовидной с их положением на побеге // Вестник фармации 3(69), 40-46.
- Литвишко В. С. 2018. Экологизация средств защиты растений // Естественные и технические науки 2 (116), 53-54.
- Скворцов А. К.1968. Ивы СССР: систематический и географический обзор. М.: Наука.
- Созинов О. В., Кузьмичева Н. Л. 2016. Сезонная и разногодичная изменчивость содержания биологически активных веществ в коре Salix viminalis (Salicaceae) в Беларуси // Растительные ресурсы 52(4), 610-619.
- Сукачев В. Н. 1934. Из работ по селекции ивы // Селекция и интродукция быстрорастущих древесных пород. Л.: Гослестехиздат, 51-85.
- Фучило Я. Д., Сбитна М. В., Фучило О. Я., Лiтвiн В. М. 2009. Створення та вирощування енергетичних плантацiй верб i тополь: Науково-методичнi рекомендацiї. К.: Логос.
- Фучило Я. Д., Сбытна М. В., Зелинский Б. В. 2018. Рост и продуктивность некоторых сортов энергетической ивы в зависимости от степени увлажненности почвы // Plant Varieties Studying and Protection 14(3), 323-327.
- Царев А. П., Погиба С. П., Тренин В. В. 2003. Селекция и репродукция лесных древесных пород. М.: Логос.
- Berlin S., Trybush S. O., Fogelqvist J. et al. 2014. Genetic diversity, population structure and phenotypic variation in European Salix viminalis L. (Salicaceae) // Tree Genetics & Genomes 10, 1595-1610.
- Fabio E. S., Kemanian A. R., Montes F. et al. 2017. A mixed model approach for evaluating yield improvements in interspecific hybrids of shrub willow, a dedicated bioenergy crop // Industrial Crops and Products 96, 57-70.
- Fuchylo Ya. D., Afonin A. A., Sbytna M. V. 2016. Selection bases of Developing new varieties of willow family (Salicaceae Mirb.) to createenergy plantations // Plant Varieties Studying and Protection 4(33), 18-25.
- Hammer D., Kayser A., Keller C. 2003. Phytoextraction of Cd and Zn with Salix viminalis in field trials // Soil Use & Management 19(3), 187-192.
- Ilnicka A., Roszek K., Olejniczak A. et al. 2014. Biologically active constituents from Salix viminalis bio-oil and their protective activity against hydrogen peroxide-induced oxidative stress in chinese hamster ovary cells // Applied Biochemistry and Biotechnology 174(6), 2153-2161.
- Jama A., Nowak W. 2012. Willow (Salix viminalis L.) in purifying sewage sludge treated soils // Polish Journal of Agronomy 9, 3-6.
- Karp A., Hanley S. J., Trybush S. O. et al. 2011. Genetic improvement of willow for bioenergy and biofuels // Journal of integrative plant biology 53(2), 151-165.
- Kuzovkina Y. A. 2015. Checklist for Cultivars of Salix L. (Willow) // FAO International Poplar Commission 11.
- Mikó P., Kovács G. P., Alexa L. et al. 2014. Biomass production of energy willow under unfavourable field conditions // Applied Ecology and Environmental Research 12(1), 1-12.
- Mleczek M., Rutkowski P., Rissmann I. et al. 2010. Biomass productivity and phytoremediation potential of Salix alba and Salix viminalis // Biomass Bioenergy 34(9), 1410-1418.
- Németh A. V., Dudits D., Molnár-Láng M., Linc G. 2013. Molecular cytogenetic characterization of Salix viminalis L. using repetitive DNA sequences // Journal of Applied Genetics 54, 265-269.
- Ollerstam O., Larson S. 2003. Salicylic acid mediates resistance in the willow Salix viminalis against the gall midge Dasineura marginemtorquens // Journal of Chemical Ecology 29, 163-174.
- Rönnberg-Wästljung A. C. 2001. Genetic structure of growth and phenological traits in Salix viminalis // Canadian Journal of Forest Research 31(2), 276-282.
- Rönnberg-Wästljung A. C., Tsarouhas V., Semerikov V., Lagercrantz U. 2003. A genetic linkage map of a tetraploid Salix viminalis'S. dasyclados hybrid based on AFLP markers // Forest Genetics 10(3), 185-194.
- Sage R. B., Tucker K. 1998. The distribution of Phratora vulgatissima (Coleoptera: Chrysomelidae) on cultivated willows in Britain and Ireland // Forest Pathology 28, 289-296.
- Trybush S. O., Jahodova S., Cizkova L. et al. 2012. High Levels of genetic diversity in Salix viminalis of the Czech Republic as revealed by microsatellite markers // Bioenergy Research 5 (4), 969-977.
- Zhao F., Yang W. 2017. Review on application of willows (Salix spp.) in remediation of contaminated environment // Acta Agriculturae Zhejiangensis 29(2), 300-306.
