Идентификация и молекулярная характеристика гранулированно-связанной синтазы крахмала, извлечённой из гречихи

Автор: Хрунгу Н.К., Деватасан Набенита, Крефт Иван, Лисен Мария
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Актуальные вопросы селекции гречихи
Статья в выпуске: 4 (25), 2010 года.
Бесплатный доступ
Обнаружены маркированные изменения в форме и размере зерен крахмала, извлеченных из разных образцов культурных сортов гречихи. Зерна отличались по форме (от круглых до полигональных) и по размеру (от 4 до 10 µm). Если зерна сорта VL-7 и IC-13145 имели четкую многогранную форму, то зерна сортов IC-133669, KBB-3, OC-2 SIVA и Дарья имели и круглую и полигональную форму. Среди образцов/ культурных сортов, исследуемых в данной работе, самыми крупными по размеру были зерна крахмала из VL-7 и KBB-3. Профили SDS-PAGE гранулировано-связанных белков, выделенных из зерен крахмала эндоспермальных тканей, обнаружили наличие зоны 59кDa, чья аминокислотная последовательность показывает >90% соответствия с белком GBSS-I, выделенным из Hardeum vulgaris, Triticum sp., Phaseolus vulgaris и Oryza sativa и 79-88% соответствия с последовательностью N-концевой аминокислоты GBSS-I у Antirrhium majus и Arabidopsis thaliana. Анализ белка GBSS-I гречихи обыкновенной (F.esculentum Moench) выявил четкое разделение на однодольные и двудольные группы. Сканирующая электронная микроскопия частично переваренных зерен крахмала обнаружила четкие модели концентрических колец с центральной сердцевиной, отражающих слоистую структуру зерен крахмала. GBSS-I был распределен в виде радиальных массивов в пределах чешуйчатой структуры зерен.
Gbss-i (гсск-гранулировано-связанная синтаза крахмала), восковой, sds-page (пагэ-электрофорез в полиакриламидном геле), clsm (кслм - конфокальная сканирующая лазерная микроскопия)
Короткий адрес: https://sciup.org/147123545
IDR: 147123545 | УДК: 633.12:
Текст научной статьи Идентификация и молекулярная характеристика гранулированно-связанной синтазы крахмала, извлечённой из гречихи
Крахмал является главным накапливаемым компонентом в семенах и других органах растения. До сих пор мы еще не конца поняли, как он синтезируется. Для того чтобы мы могли управлять процессом синтеза крахмала в зерновых культурах для получения больше крахмала или крахмалов с необходимыми свойствами, прежде всего мы 70
Marked variations were observed in the shape and size of starch grains from different accessions/cultivars of buckwheat. The grains ranged in shape from round to polygonal ranging in size from 4 µm to 10 µm .While the grains from VL-7 and IC- 13145 showed distinct polygonal shape, those from IC-133669. KBB-3, OC-2, SIVA and Daria were round to polygonal in shape. Amongst the accessions/cultivars investigated in the present study, the starch grains from VL-7 and KBB-3 were the largest in size. SDS-PAGE profiles of granule bound proteins isolated from the starch grains of endosperm tissues revealed the presence of 59 kDa band whose amino acid sequence showed >90% homology with GBSS-I protein isolated from Hordeum vulgaris, Triticum sp. , Phaseolus vulgaris and Oryza sativa and between 79-88% homology with the N-terminal amino acid sequence of GBSS-I from Antirrhium majus and Arabidopsis thaliana. Blastp analysis of GBSS-I protein of common buckwheat (F.esculentum Moench.) revealed a clear diversification into monocotyledonous and dicotyledonous groups. Scanning electron microscopy of partially digested starch grains revealed a clear pattern of concentric rings with a central core revealing the lamellar structure of starch grains. GBSS-I was organized as radial arrays within the lamellar structure of the grains.
должны понять процесс синтеза крахмала, и как он контролируется. Одной из наиболее важных задач в понимании того, как производится крахмал, является объяснение того, как два разных полимера, составляющих крахмал, амилоза и амилопектин синтезируются в одном месте и в одно время. Тем не менее, это увеличивает сложность процесса и усложняет процесс интерпретации, а также расширяется круг возможностей для биоинженерии. Программируя специфические формы энзима (таких как GBSS), можно осуществить контроль за типами производимых молекул.
Амилоза – это в большинстве случаев линейный α-1, 4-связаный глюкан, в то время как амилопектин – это разветвленный полимер α-1, 4-связанные цепочки с разветвлениями α-1, 6-соединения. Амилоза и амилопектин синтезируются деятельностью сложных энзимов, включающие глюкозные фосфорилазы ADP, синтазы крахмала (SSs), энзимы, разветвляющие крахмал (SBEs), энзимы, расщепляющие крахмал (Slattery et al., 2000; Myers et al, 2000; Smith., 2001; Nakamura., 2002; Ball and Morell., 2003). Сложные SS изозимы были обнаружены в различных видах растений и классифицированы на пять классов (GBSSI,SSI,SSII,SSIII и SSIV), основанных на их первичных последовательностях (Ball and Morell., 2003; and Smith, A.M., 1999). Оказывается, каждый изозим имеет отличительные свойства и играют особую роль в биосинтезе крахмала (Smith et al., 1997; and Kossmann et al., 2000).
Гречиха обыкновенная (Fagopyrum esculentum Moench), которая считается малоиспользуемой культурой из-за недостаточных капиталовложений в исследование ее потенциала, имеет важное экономическое значение в некоторых странах мира.
Мука из гречихи используется народами, живущими в предгорьях индийских Гималаев, так же как и в других странах мира, в том числе в Японии, Корее, Польше, Китае, СССР, Словении для приготовления лапши, макарон, хлеба и блинов.
Эта культура так же является большим источником белков, углеводов, глюкозидов, включая рутин. Тем не менее, из-за неблагоприятного соотношения амилозы / амилопектина тесто из гречневой муки имеет низкие пекарские качества. По мнению Inoue et al (2004), содержание амилозы, белка и других химических составляющих, которые имеют отношение к клейкости муки, показали богатые вариации в пределах генотипа. Так как соотношение амилозы и амилопектина влияет на текстуру и качество муки, определение локуса гена, вовлеченного в регулирование синтеза крахмала и его осаждения, окажет влияние на разработку эффективных программ племенной работы, направленных на улучшение качества крахмала. Эти локусы могут быть использованы как маркеры для определения генотипов с низким содержанием амилозы крахмала в зерновых. Хотя такие белки были изолированы, и их гены клонированы из традиционных зерновых культур, ни один из гранулировано-связанных белков в гречихе был определен, и ни один из его генов не был клонирован.
Материалы
Сертифицированные образцы гречихи (Fagopyrum esculentum Moench) были взяты из генетического банка плазмы гречихи Национального Бюро Генетических Ресурсов Растений, Ш имла, Северо-Восточной региональной станции NBPGR, лаборатории Ш илонг и Вивикананда для горного земледелия, Альмора. Европейские образцы/культурные сорта пшеницы были взяты из коллекции генетической плазмы университета Любляна, Словения, Европа.
Выделение крахмала из зерна было произведено в соответствии с описанием Nakamura et al.(1999). Содержание амилозы и амилопектина в крахмале было определенно как per Oda et al. (1992). Профиль (внешний вид) SDS PAGE гранулировано связанных белков был определен в соответствии с Nakamura et al. (1993). Имуноблотинг белков из гранул крахмала и растворимых фракций были выполнены с помощью кроличьей сыворотки anti-GBSS после SDS PAGE. SEM гранул крахмала была произведена в соответствии с David et al. 2006.
Результаты и обсуждение
Гранулы крахмала, выделенные из эндоспермальных тканей различных Европейских и Индийских образцов/культурных сортов гречихи, были проанализированы по видам, формам, размерам и другим морфологическим характеристикам с использование Сканирующей электронной микроскопии. Форма зерен была разнообразна: от круглых до полигональных.
В то время как зерна VL-7 и IC-13145 имели четкую полигональную форму, зерна IC-188669, KBB-3,OC-2. Сива и Дарья были круглыми и сферическими по форме. Зерна отличались по размеру от 4 µm до 10 µm. Зерна крахмала европейских сортов Сива и Дарья были значительно меньше по размеру в сравнении с индийскими сортами (рис. 1). Зерна крахмала, выделенные из сортов гречихи, доставленных из Индии и Словении, отличались по размеру от 4-10 мм и по форме: от круглой до полигональной.
Сканирующая электронная микроскопия зерен крахмала (рис. 1) четко показывает наличие белкового слоя, покрывающего зерна крахмала гречихи. Эта мембрана может быть включена в поддержание структурной целостности зерна. Сканирующая электронная микроскопия частично переваренных зерен крахмала показывает четкую модель концентричных колец, указывающих на чешуйчатую структуру зерен крахмала (рис. 1.). Такие слоистые структуры были предложены, чтобы представить чередование полукристаллической и аморфной зоны в пределах матрицы.






Рисунок 1 – Сканирующая электронная микроскопия зерен крахмала, выделенных из различных образцов/культурных сортов Европейской и Индийской гречихи. (I)IC-188669, (II) KKB-3, (III) VL-7, (IV) IC-13145, (V) OC-2, (VI) IC-324244 (VII) IC-363973 (VIII) SIVA, RANGUS, (IX) DARIA, (X) четко показывает зерна крахмала, укрытые протеиновой мембраной и (XI) (XII) – картина крахмала зрелого зерна, выделенного акустически и инкубированного с 3u/ml альфа-амилазой в течение 15 мин при комнатной температуре
Анализ SDC-PAGE гранулировано-связанных белков разных образцов/культурных сортов гречихи из Словении обнаружил наличие дуплекса, соответствующего молекулярной массе 59 и 56 kDa (рис. 2(А)). С другой стороны, образцы/культурные сорта из Индийских Гималаев показали наличие только одной зоны (полосы), относящейся молекулярной массе 59 kDa (рис. 2(Б)). Иммуноблоттинг выделенных SDS-PAGE белков с антителами по сравнению с гранулировано-связанными белками гречихи показал сильную перекрестную реактивность с белком 59 kDa (рис. 3(А, Б)). Это показывает, что белок 59 kDa, идентифицированный на профилях SDS-PAGE гранулировано-связанных зерен белка, может быть GBSS-I. Белок 56 kDa, присутствующий в зернах крахмала Европейских сортов, не показал никакой перекрестной реактивности с антителами по сравнению с гранулировано-связанным белком (рис. 2). SDS-PAGE профиль гранулировано-связанного белка крахмала, выделенного из зрелых листьев гречихи, показал наличие единственной зоны, соответствующей молекулярной массе белка 53 kDa (рис. 2(Б)). Тем не менее, гранулировано-связанный белок антисера, который не смог распознать белок 53 kDa (рис. 2(Б)), указывая на то, что белок 53 kDa не обладает сходством с гранулировано-связанным белком, выделенным из зерен крахмала эндоспермы гречихи. SDS PAGE профиль образцов/культурных сортов гречихи как из Индийских Гималаев, так и из Словении не показал наличия высоко молекулярных белков, которые присутствуют в других растениях, таких как горох и пшеница. Это может иметь отношение к процедуре экстрагирования, которая ведет к полной потере всех белков, имеющихся в периферийных частях зерна. Так как GBSSI локализуется в центре зерен крахмала, он задерживается в процессе экстракции.

Рисунок 2 (А) – Профиль SDS PAGE гранулированно-связанного белка, выделенный из зерен гречихи европейского происхождения.
2 [I]: -(M) = Маркер молекулярной массы белков , 1=Bf06x12, 2=Gor0619Siva Min Selo, 3=41 Min Gor05, 4=Siva Kontrola Rangus PR03, 5=53 Gor Siva Min 05, 6=Gor 0620 Siva Min Selo. 2 [II]: -(M) Маркер молекулярной массы белков , 1= Gor 1304BF0611, 2=63Min Gor Siva 05, 3=Gor0628 Siva Min Selo, 4=Gor 0413BF0618, 5= Gor 0413BF0618. 2 [III]: -(M) = Маркер молекулярной массы белков , 1=Gor 11 2004 3 CRTE, 2= Gor 2 2004 3 CRTE, 3=Gor 4 2004 3 CRTE, 4= Gor 10 20043 CRTE, 5=191 2X rumena 3 CRTE. 2 [IV]: -(M) = Маркер молекулярной массы белков , 1=Gor 93 CRTE, 2=7Roza CRTE, 3=10RDECA 3 CRTE
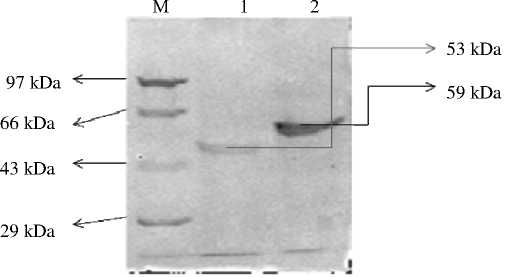
1 2

Рисунок 2 (Б) – Профиль SDS PAGE гранулированно-связанного белка, выделенный из зерен крахмала листьев и семян сорта гречихи, полученного в Индии. М= маркер , 1= связанный белок, выделенный из зерен крахмала листьев, 2= связанный белок, выделенный из зерен крахмала семян
Рисунок 3 (Б) – Иммуноблоттинг связанного белка, выделенного из листьев и семян, сорта, полученного в Индии, протестированный с помощью 56 kDa антисеры для выявления связанного белка гречихи. Перекрестнореактивные белки были обнаружены с помощью антитела белка 56 kDa и с помощью кроличьей HRP и козьей IgG сыворотки

Рисунок 3(А) – Иммуноблоттинг гранулировано-связанного белка, выделенного из зерен крахмала сорта гречихи, выращенной в Европе с помощью антисеры белка kDa для выявления связанного белка гречихи из Индии. Перекрестнореактивные белки были обнаружены с помощью антитела белка 56 kDa и с помощью кроличьей HRP и козьей IgG сыворотки
Гранулы крахмала, выделенные из созревшего эндосперма, были окрашены йодидом калия и раствором йода (0,2% KI, 0,4% I w/v) и рассмотрены под оптическим микроскопом (рис. 4). Молекулы йода, как известно, образуют комплекс с крахмалом, обладая бóльшим сродством с амилозой, по сравнению с аминопектином. Ядро гранул крахмала состояло из концентрических колец с видимым центральным ядрышком, окрашенным с помощью йодида калия и раствора йода в синий цвет, таким образом, показывая высокое содержание амилозы.
Конфокальная лазерная сканирующая микроскопия четко показала наличие гранулированной связанной синтазы крахмала (GBSS-I.) в ядре зерен крахмала (рис. 5). Белок оказался расположенным в форме радиальных массов, выходящих из ядра зерен. Изображения гранул крахмала, окрашенных йодидом калия и раствором йода, полученными конфокальной лазерной сканирующей микроскопией и оптическим микроскопом, аналогичны, это доказывает, что синтез амилозы происходит в ядре гранул крахмала, отмеченных ярким гелием, окруженным толстыми кольцами роста (рис. 4).
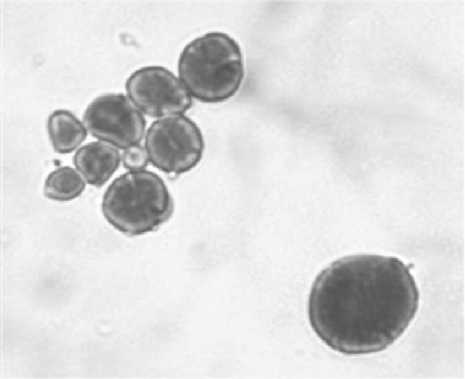
Рисунок 4 – Изображения , полученные в результате световой микроскопии гранул крахмала, полученных из зрелой эндоспермы, окрашенной раствором иодида калия и йода
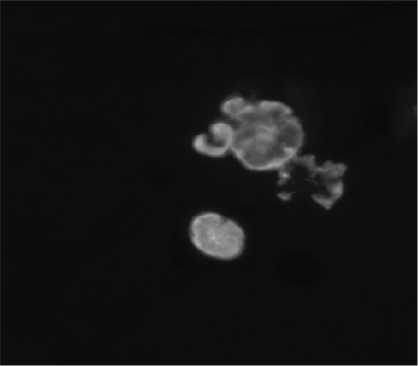
Рисунок 5 – Изображения конфокальной лазерной сканирующей микроскопии гранул крахмала.
Физиохимическая локализация GBSS-I на зернах крахмала была проведена с помощью иммунодетекции с флуоресцентным маркированным вторичным антителом (Alexa 546 – соединенная с козье и антикроличьей сыворотками IgG)
N-концевая аминокислотная последовательность для 25 радикалов 59 kDa гранулированного связанного протеина формулируется следующим образом:
GMLNVFVGAEVAPWSKTGGLGDVLA
Последовательность была помещена на хранение в SWISS-PROT банке данных белков с доступом № P 84633 . BLASTp анализ аминокислоты определил белок как GBSS-I. Отличительной чертой последовательности является наличие консервативной области, имеющей последовательность KTGGL, которая является ADP-Glc связывающим участком для фермента. N-концевая аминокислотная последовательность 25 радикалов для протеина показала гомологию с известными GBSS последовательностями другого хлебного злака (рис. 6). Используя линейную форму, что позволило получить максимальную гомологию, обыкновенная гречиха ( Fagopyrum esculentum Moench.) показала наивысшую гомологию (94%) с белком GBSS-I , выделенным из Hordeum vulgaris , Triticum sp. и Phaseolus vulgaris. Однако гомология колебалась в процентном отношении между 94% и
88% с GBSS из других хлебных злаков. Филогенетический анализ аминокислотных последовательностей белка GBSS из обыкновенной гречихи, описанной здесь, обнаруживает четкое деление на односемядольную и двусемядольную группы (рис. 7). Односемядольные растения делятся в свою очередь на две подгруппы: одна подгруппа – с исключительным доминированием риса (Oryza sativa), а другая – с доминированием пшеницы (Triticum aestivium) и ячменя (Hordeum vulgare). У риса (Oryza sativa) 4-ый и 11-ый аминокислотные радикалы ГССК были V (валин) и M (метионин) вместо L (лейцин) и V (валин), как в аминокислотных последовательностях обыкновенной гречихи, описанной здесь. В то же время, у пшеницы (Triticum aestivium) и ячменя (Hordeum vulgare) 11-ая аминокислота – M (метионин), в отличие от обыкновенной гречихи (Fagopyrum esculentum Moench.), у которой присутствует V (валин). Возможно, это одна из главных причин деления односемядольных растений на две подгруппы в результате филогенетического анализа.
GMLH V FVGAE V APWS KT GGLGD VL GMi L1FVGAE VAPWS KT GGLGD Vb GMSVIFIGAEVGPWSKTGtiLGDVL GMUGU-VGIE VAPWSKTWLGLIVL GMHLVFVtiCEVGPWC KT GGLGD VL GM VFVLAEVGPWS KT GGLGD VL
(Fagopyrum esculentum) (Pliaseolus vulgar is ) ( Ar abidopsis thaliana) (visum sativum) (Ipomoea batatas) (Ant ix rhinum majus)
GMMVVFVGAEMAPWSKTGGLGDVL GhUIVVtVI.ttEMftPWSKTWLtiLIVL GMHLVFVGAEMAPWSKT GGLGD VL
gmnlvfvgaemapwskt gglgd vl
< »1-ШЬ V FV G flEMAPW S KT GGLGD VL GMHLVFVGAEMAPWSKT GGLGD VL GMHV V b'Vf.AbMAPWTT GGLGD VL 11LVFVGAEMAPWSKT GGLGD VL HL VFVGAEMAPWSKT GGLGD VL
(Oryza sativa) ( Hordeum vu 1g ar ae ) (Triticum astivum) (setaria it al lea) Aegilops speltoides) (Aegilops tauschii) (MOIKKDTS)
Рисунок
6 – N-
концевая аминокислотная последовательность обыкновенной гречихи и расположение ее по одной линии с
N-
концевыми аминокислотными последовательностями сходных белков
,
находящихся в наличии в банке данных протеинов
--------pt WJ«J5 tfy 5*4585# f.r
_____p
$i 6624283 dtj MA586
f J f
^Lyt
«4«jj
эд aurewt*
__pet t?flM8t ли ая;?Улг ji Kt$>7$ pi Mf3*i«.l _ppi 'Ч»И ®j BMlTMt.t t-pi 4186680 to) ЙМ71360 Г ppi 71667448 ft ДЛМГ4053Л
- I e< srejriM nojUJvydMe. f
p --
$< ?tM?41i jbHJtt«JlJ
----pf 7W57«e (6
ААЯ74049
f
------p
11667*34 6bAA)T?4M0.!
-----jt r^erw jt axMwtj.r pet 16», 177» 46i BA663Z09 t
J-^L^
S167:60gt 4Ali4iZ«t 1
— — pt 7?w$s
i tt
Ai re 1
675
г
----pi
567421 mb £*146471
f
-----pi 8l»M»y jt
AAT716$l I
--------pi
448ВД854
d^
BAD12044 i
---------pt
*4в4в6$г ibj BAbtiOti I
--------pt zrefstffj «j at»r?»a# ---------pt
MS 16
1JJ
dtj Я4С4
г
2M. I
-------0
16716W5
<4j aiFjre^ 1
ppt4i7fdMS
iibj ВА0218И1
Lp
ИГ,М»?
ti, it6?jM?
1
-------pi Г «имя
(5ЛКМ7И4,1
--------pt
6624287 to) ЙМ865Г2
Г -------P^t/ffnnJmt«"l?^
ppi
16623566 mb CAC6M65.1
*-^ smiti ,toj BM623481
_J-f«
*5778156 461 BAB685751
Lft
15837971 itjfiASMliei
Triticum
sps.
Hordt* um
sps
Oi у zu
sps.
Hordeum
sps.
GBSS seq. from buckwheat ♦Pisum sativum I роста batatas Pltoseolus vulgaris Рисунок 7 – Филогенетическое древо, основанное на линейной матрице N-концевой аминокислотной последовательности 59 кДа связанного белка, выделенного из гречихи с N-концевыми аминокислотными последовательностями
GBSS-I
протеина из различных хлебных злаков
Благодарность
Авторы благодарят Департамент Науки и Технологии
,
правительство Индии
,
Нью Дели и Министерство высшего образования
,
правительство Словении за финансовую поддержку в рамках программы кооперации Индо
-
Словенской науки и технологии
.
(ФГОУ ВПО Орёл ГАУ)


Список литературы Идентификация и молекулярная характеристика гранулированно-связанной синтазы крахмала, извлечённой из гречихи
- Ball, S. G., and Morell, M. K. (2003). From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol., 54: 207-233
- David, G. S.; Russel, K. D.; Jay-lin, J.; George, E. I.(2006) Structures and functional properties of starch from seeds of three soyabean (Glycine max (L.) Merr.) varieties. Starch/Starke.,58: 509-519
- Inoue N., Kaori, F., Zhongfa, Y., Masakazu, K., Shu-hei, U., Motoyuki, H. and Ujihara, A. (2004). Effects of environmental factors on the chemical characteristics of common buckwheat in relation to flour texture (Variation in amylose and crude protein content of seeds collected at various sites in Japan). Fagopyrum 21: 65-69
- Kossmann, J., and Lloyd, J., (2000).Understanding and influencing starch biochemistry. Crit. Rev. Plant Sci., 19: 171-226
- Myers, A. M., Morell, M. K., James, M. J., and Ball, S. G., (2000). Recent progress towards understanding of the amylopectin crystal. Plant Physiol. 122: 989-997
- Nakamura, T., Virnten.P., Hayakawa, K.and Ikeda, J. (1998). Characterization of a granule bound starch synthase isoform found in the pericarp of wheat. Plant Physiol., 118: 125-132
- Nakamura, T., Yamamori, M., Hirano, H. and Hidaka, S. (1993). Identification of the three waxy protein in wheat (Triticum aestivium L.). Biochem. Genet., 248: 253-259
- Nakamura, Y., (2002). Towards a better understanding of the metabolic system for amylopectin biosynthesis in plants: rice endosperm as a model tissue. Plant Cell Physiol., 43: 718-725
- Oda, S., Kiribuchi, C., and Seko, H. (1992). A bread wheat mutant with low amylose content induced by ethyl methanesulphonate. Jap. J. Breed., 42: 151-154
- Slattery, C. J., Kavakli, I. H., and Okita, T. W., (2000). Engineering starch for increased quantity and quality.Trends Plant Sci., 5: 291-298
- Smith, A. M., (1999). Making starch. Curr. Opin. Plant Biol., 2: 223-229
- Smith, A. M., (2001).The biosynthesis of starch granules. Biomacromolecules 2: 335-341
- Smith, A. M., Denyer, K., and Martin, C., (1997). The synthesis of the starch granule. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 67-87
