Идентификация сортов, линий и мутантов гороха посевного с помощью RAPD-маркеров

Автор: Дрибноходова О.П., Кокаева З.Г., Гостимский С.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: ДНК-технологии в растениеводстве
Статья в выпуске: 5 т.40, 2005 года.
Бесплатный доступ
Проводили анализ ДНК-полиморфизма 19 форм гороха посевного (Pisum sativum L.) по 134 RAPD-маркерам. Определяли генетическое сходство, отражающее степень различий RAPD-спектров исследованных образцов. Рассчитывали генетические расстояния между линиями, сортами и мутантами гороха посевного. Оценивали эффективность RAPD-метода для паспортизации и идентификации линий и сортов растений.
Короткий адрес: https://sciup.org/142133119
IDR: 142133119 | УДК: 633.358:577.2
Identification of pea cultivars, lines and mutants by use of RAPD-markers
Analysis of DNA polymorphism of 19 pea lines by using RAPD-markers was carried out. The level of genetic diversity of different pea lines was investigated. The data set for 19 lines including 134 binary characters was used to generate UPGMA and NJ-dendrograms suggesting genetic relationships among pea cultivars, lines and mutants. RAPD-markers that can be used for identification of pea cultivars were found. The opportunity of using RAPD-markers for the identification of different pea cultivars and for genetic mapping was discussed.
Текст научной статьи Идентификация сортов, линий и мутантов гороха посевного с помощью RAPD-маркеров
В последние годы проблема идентификации видов и сортов растений становится все более актуальной в связи с ускорением селекционного процесса и появлением большого числа новых форм. Для создания нового сорта необходимы предварительная регистрация существующих генотипов, контроль за уровнем полиморфизма полученных при скрещивании партий семян и чистотой селектируемой линии. Экспресс-анализы сортопринадлежности образцов семян могут быть использованы для решения спорных вопросов в семеноводстве и для защиты прав оригинатора сорта (1, 2). Очевидно, что современная практика семеноводства нуждается в разработке быстрых и точных методов идентификации каждого сорта. Морфологические признаки не пригодны для быстрой проверки семян на сортовую принадлежность, так как требуют больших затрат времени и труда, особенно в случае необходимости разделения сортов, имеющих сходные фенотипы (3). Кроме того, при таком подходе трудно отличить генетические изменения от модификаций, вызванных влиянием окружающей среды (4).
В настоящее время для генотипирования все чаще используют ДНК-маркеры, причем наиболее часто — RAPD- и SSR-маркеры (2, 5). Эти методы обладают высокой производительностью, получаемые с их помощью результаты не зависят от условий выращивания и стадии развития растений, продолжительности и условий хранения семян. С помощью RAPD-метода можно анализировать изучаемые формы одновременно по многим локусам, случайно распределенным по всему геному, что позволяет различать даже очень близкородственные сорта (6). Хотя многие авторы отмечают недостаточную надежность RAPD-метода, оптимизация условий и отбор стабильно амплифицирующихся ДНК-маркеров позволяют добиться воспроизведения данных, полученных в разных лабораториях, и поставить вопрос о разработке дешевых, быстрых и эффективных технологий паспортизации разных линий, сортов и гибридов гороха на основе RAPD-метода. За последние годы были проведены работы по изучению генетического полиморфизма различных культур: ячменя, кукурузы, подсолнечника, картофеля, хлопчатника, пшеницы и др. (1, 3, 4, 6-9).
В задачу нашей работы входила идентификация и паспортизация сортов, линий и мутантов гороха посевного с помощью RAPD-маркеров.
технология», Москва) в следующем режиме: предварительная денатурация 2,5 мин при 94 о С; денатурация 30 с при 94 о С; отжиг 30 с при 40 о С; синтез 1,3 мин при 70,5 о С (всего 5 циклов); денатурация 20 с при 93 о С; отжиг 30 с при 37 о С; синтез 1 мин при 72 о С (всего 35 циклов); конечная элонгация 5 мин при 72 о С. Реакционная смесь объемом 25 мкл содержала 2 е.а. Taq-полимеразы («Силекс М», Москва), 2,5 мкл стандартного 10-кратного буфера для ПЦР («Силекс М», Москва), 25 пкмоль праймера, 2,5 мМ MgCl 2 , 0,5 или 0,25 мМ дезоксинуклеозидтрифосфата. На смесь наслаивали две капли минерального масла.
Продукты амплификации разделяли электрофорезом в 1,7 % агарозном геле с использованием буфера ТВЕ и окрашивали бромистым этидием. Для определения длины фрагментов использовали маркеры молекулярной массы 100 bp + 1,5 Kb DNA Ladder и 1 Kb DNA Ladder («Сибэнзим», Новосибирск). После электрофореза гели анализировали в УФ-свете и фотографировали с использованием цифровой фотокамеры.
Для проверки воспроизводимости полученных результатов эксперименты проводили в нескольких повторностях с использованием двух концентраций MgCl 2 (0,5 и 0,25 мМ) и разных партий полимеразы. В дальнейшей работе использовали только те фрагменты, о наличии или отсутствии которых в спектрах всех сортов можно было судить с достаточной степенью уверенности. Каждому полученному маркеру было дано название, состоящее из двух частей: первая часть соответствует праймеру, при амплификации с которым был обнаружен фрагмент, вторая — отражает приблизительный размер фрагмента (п.н.).
Для количественной оценки полиморфизма и определения генетического расстояния между исследованными формами данные RAPD-анализа были представлены в виде матрицы, в которой наличие или отсутствие в RAPD-спектрах фрагментов одинаковой длины соответствовало значениям 1 или 0. На основе этой матрицы были составлены две матрицы различий, генетические расстояния (ГР) в которых рассчитывали по формуле
Ч исло полиморфных маркеров
ГР =-----------—----- X 100%.
Общее число маркеров
Генетическое расстояние между сравниваемыми линиями определяли по формуле Nei с соавт. с помощью компьютерной программы Treecon (11, 12).
На основании полученной невзвешенным парно-групповым методом (Unweighted Pair-Group Method — UPGMA) матрицы мы составили генеалогическую дендрограмму, отражающую степень различий между RAPD-спектрами исследуемых форм гороха (12). Дендрограмму строили по компьютерной программе Treecon с применением 100 реплик бутстрепа (12, 13). Матрица различий, полученная по формуле Nei
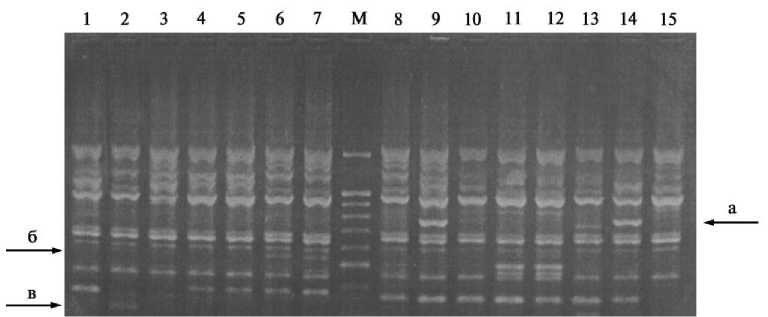
Рис. 1. RAPD-спектры линий, мутантов и сортов гороха посевного, полученные при амплификации ДНК с праймером B474: 1 — Флагман; 2 — Филби; 3 — Труженик; 4 — Виола; 5 — Виола (В); 6 — Хл-1; 7 — Хл-2; 8 — Хл-15; 9 — Хл-18; 10 — Хл-42; 11 — L-1238; 12 — L-1132; 13 — L-851; 14 — L-111; 15 — L-108; М — маркер молекулярной массы 100 bp + 1,5 Kb DNA Ladder. Стрелками отмечены полиморфные фрагменты: а, б и в — соответственно B474#730, B474#530 и B474#300.
с соавт., лучше отвечает известным данным о происхождении сортов и линий, что соответствует данным литературы (9).
Результаты. Межлинейный и межсортовой полиморфизм оценивали у семи сортов, шести линий и шести мутантов гороха посевного с использованием десяти высокоэффективных RAPD-праймеров (рис. 1). При амплификации ДНК разных образцов гороха посевного отдельные праймеры выявляли от 9 до 19 фрагментов размером 230-2900 п.н., 7-14 из которых были полиморфными; всего получено 134 RAPD-маркера, 102 из которых были полиморфными среди изученных форм гороха
(табл. 1).
Для количественной оценки полиморфизма и определения степени дивер-
0,2
0,1
76 % । -------------Немчиновский
^-----------R 64
--------------------Хл-42
-----------------Флагман
-----------------Труженик
--------------------------------------L-111
I-------------------------Филби
1-------------------------- Хл-18
Хл-115 Капитал Хл-1 Хл-2
|
1 |
|
70 %
77 %
100 % । Виола Виола (В) Демон L-108 L-851
L-1238
L-1132
генции между изученными формами данные, полученные при оценке RAPD-полиморфизма, были представлены в виде матрицы по 134 бинарным признакам, которую использовали для расчета генетических расстояний и построения дендрограммы (рис. 2). Степень различий между мутантами, полученными от одного сорта, составляла 1,514,9 %; от исходных форм — Виола и Хл-42 — они отличались соответственно на 0 и 20,9 %. Средний уровень полиморфизма между изученными формами составлял 25,6 %, что согласуется с ранее полученными данными (14).
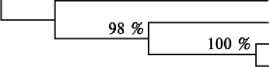
Рис. 2. Генеалогическая дендрограмма, отражающая степень различий между RAPD-спектрами линий, сортов и мутантов гороха посевного (UPGMA-метод) (12): 0,1 и 0,2 — генетическое расстояние, по Nei с соавт. (11) .
-
"^ 1. Оценка полиморфизма ДНК у различных линий, сортов и мутантов гороха посевного ( Pisum sativum L.) с помощью RAPD-маркеров
Сорт, мутант, линия
Длина ДНК-фрагментов, полученных при амплификации с праймером QR2, п.н.
1600
1550
1500
1400
1300
1250
1200
1100
1000
910
810
790
740
620
535
410
350
320
Флагман
+
+
–
+
–
+
+
–
+
+
–
+
+
+
–
+
–
–
Филби
+
–
–
–
–
+
+
–
+
+
–
–
+
+
–
–
–
–
Труженик
+
+
–
+
+
+
+
–
+
+
–
–
+
+
–
–
+
–
Виола
+
+
–
+
–
+
+
–
+
+
–
+
+
+
–
–
–
–
Виола (В)
+
+
–
+
–
+
+
–
+
+
–
+
+
+
–
–
–
–
Капитал
+
+
–
–
+
+
+
–
+
+
–
–
+
+
–
–
–
+
Немчиновский
+
–
–
–
–
+
+
–
+
+
–
–
+
+
–
–
+
–
Демон
–
–
–
+
+
+
+
–
+
+
+
–
+
+
–
–
–
+
Хл-1
+
–
+
–
+
+
+
–
+
+
–
–
+
+
–
–
–
–
Хл-2
+
–
+
–
+
+
+
–
+
+
–
–
+
+
–
–
–
–
Хл-115
+
–
–
–
–
+
+
–
+
+
–
+
+
+
–
–
+
+
Хл-18
–
–
+
–
+
+
+
–
+
+
+
–
+
+
–
+
–
+
Хл-42
+
–
–
–
–
+
+
–
+
+
–
–
+
+
–
–
+
+
L-1238
+
–
–
+
–
+
+
+
+
+
–
–
–
+
+
–
–
–
L-1132
+
–
–
+
–
+
+
+
+
+
–
–
+
+
+
+
–
+
L-851
+
–
–
+
–
+
+
+
+
–
–
–
+
+
+
+
+
–
L-111
–
+
–
–
–
+
+
–
+
+
–
+
+
+
+
–
+
+
L-108
–
+
–
+
+
+
+
+
+
+
–
–
+
+
–
–
+
–
R-64
+
–
–
–
–
+
+
–
+
+
–
–
+
+
–
–
+
+
-
2. Матрица генотипов для тестирования линий, сортов и мутантов гороха посевного ( Pisum sativum L.) с помощью RAPD-маркеров (фрагмент общей матрицы)
3. Специфические маркеры для тестирования различных образцов гороха посевного
П р и м е ч а н и е: (+) и (–) означает соответственно наличие и отсутствие маркера.
|
Сорт, мутант, линия (исходный сорт) |
Длина ДНК-фрагментов, полученных при амплификации с праймером QR2, п.н. |
||||||||||||
|
1600 |
1550 |
1500 |
1400 |
1300 |
1100 |
810 |
790 |
740 |
535 |
410 |
350 |
320 |
|
|
Флагман |
+ |
+ |
– |
+ |
– |
– |
– |
+ |
+ |
– |
+ |
– |
– |
|
Филби |
+ |
– |
– |
– |
– |
– |
– |
– |
+ |
– |
– |
– |
– |
|
Труженик |
+ |
+ |
– |
+ |
+ |
– |
– |
– |
+ |
– |
– |
+ |
– |
|
Виола |
+ |
+ |
– |
+ |
– |
– |
– |
+ |
+ |
– |
– |
– |
– |
|
Капитал |
+ |
+ |
– |
– |
+ |
– |
– |
– |
+ |
– |
– |
– |
+ |
|
Немчиновский |
+ |
– |
– |
– |
– |
– |
– |
– |
+ |
– |
– |
+ |
– |
|
Демон |
– |
– |
– |
+ |
+ |
– |
+ |
– |
+ |
– |
– |
– |
+ |
|
Хл-115 (Торсдаг) |
+ |
– |
– |
– |
– |
– |
– |
+ |
+ |
– |
– |
+ |
+ |
|
Хл-18 (Ранний зеленый) |
– |
– |
+ |
– |
+ |
– |
+ |
– |
+ |
– |
+ |
– |
+ |
|
L-1238 |
+ |
– |
– |
+ |
– |
+ |
– |
– |
– |
+ |
– |
– |
– |
|
L-111 |
– |
+ |
– |
– |
– |
– |
– |
+ |
+ |
+ |
– |
+ |
+ |
|
L-108 |
– |
+ |
– |
+ |
+ |
+ |
– |
– |
+ |
– |
– |
+ |
– |
П р и м е ч а н и е. То же, что в таблице 1.
|
Маркер |
1 Сорт, мутант, линия |
|
B474#530 |
Капитал |
|
B474#300 |
Филби |
|
QR2#1500 |
Хл-18 |
|
K10#570 |
Виола |
|
V#630 |
Виола (В) |
|
V#480 |
Флагман |
|
V#430 |
Хл-18 |
|
AD04#650 |
L-1238 |
|
Q20#650 |
Капитал |
|
Pr10#410 |
Флагман |
На основе матрицы различий всех исследованных образцов гороха посевного UPGMA-методом была построена дендрограмма, отражающая степень различий между RAPD-спект-рами исследованных форм; значимость генеалогических реконструкций определяли методом бутстрепа (13) (см. рис. 2).
Топология генеалогического дерева в участках со значениями больше 50 % может считаться на- дежно установленной. Полученная схема соответствует известной информации о происхождении изучаемых форм.
Для оценки возможности применения RAPD-метода с целью идентификации линий и сортов гороха посевного из исследованных форм было отобрано семь сортов, три маркерные линии и два мутанта. На основе RAPD-спектров изученных форм (см. табл. 1) была разработана матрица генотипов, в которую включены 95 полиморфных маркеров (табл. 2).
Для однозначной идентификации генотипов выбранных сортов, линий и мутантов гороха посевного достаточно анализа 10-15 полиморфных маркеров, то есть возможно определение сорта на основе анализа его RAPD-спектра по нескольким праймерам. В нашей работе каждый из трех RAPD-праймеров (QR2, K10, R03) позволял получать такой набор фрагментов анонимной ДНК, который точно соответствовал одному из RAPD-спектров исследованных 12 сортов, линий и мутантов гороха посевного. Следовательно, определение сортового соответствия выбранных нами образцов гороха посевного (ПЦР, электрофорез и анализ полученных данных) занимает 8 ч (один рабочий день). Среди полиморфных маркеров было выявлено 10 маркеров, амплифицирующихся только в одном генотипе (табл. 3).
Итак, полученные нами данные свидетельствуют о том, что RAPD-метод позволяет получать специфические геномные маркеры, которые можно использовать для паспортизации и идентификации видов и сортов гороха. RAPD-маркеры, специфические для определенных линий или сортов, можно применять для быстрой идентификации этих генотипов, проведения генетического анализа, изучения родственных связей между различными формами гороха посевного, а также в работе по картированию хромосом.
Л И Т Е Р А Т У Р А
-
1. С и в о л а п Ю.М., Т о п ч и е в а Е.А., Ч е б о т а р ь С.В. Идентификация и паспортизация сортов мягкой пшеницы методами RAPD- и SSR-анализа. Генетика, 2000, 36, 1: 44-51.
-
2. Х а в к и н Э.Е. Молекулярная селекция растений: ДНК-технология создания новых сортов сельскохозяйственных культур. С.-х. биол., 2003, 3: 26-41.
-
3. П о м о р ц е в А.А., Л я л и н а Е.В. Идентификация и оценка сортовой чистоты ячменя методом электрофореза запасных белков семян. М., 2003.
-
4. К о ч и е в а З.Г., О г а н и с я н А.С., Р ы с к о в А.П. RAPD-маркеры генома картофеля: клонирование и использование для определения межвидовых и межсортовых различий. Мол. биол., 1999, 33, 5: 893-895.
-
5. Г о с т и м с к и й С.А., К о к а е в а З.Г., Б о б р о в а В.К. Использование молекулярных маркеров для анализа генома растений. Генетика, 1999, 35, 11: 1538-1549.
-
6. К о ч и е в а Е.З., Р ы ж о в а Н.Н., Х р а п а л о в а И.А. и др. Использование метода RAPD-анализа в определении генетического полиморфизма и филогенетических связей у представителей рода Lico-persicon (Tourn.) Mill. Генетика, 2002, 38: 1298-1303.
-
7. К о ж у х о в а Н.Э., С и в о л а п Ю.М. Идентификация и регистрация генотипов кукурузы при помощи молекулярных маркеров. Генетика, 2004, 40, 1: 59-66.
-
8. С и в о л а п Ю.М., С о л о д е н к о Е.А., Б у р л о в В.В. RAPD-анализ молекулярно-генетического полиморфизма подсолнечника ( Heliantus annuus ). Генетика, 1998, 34, 2: 266-271.
-
9. К у д р я в ц е в А.М., М а р т ы н о в С.П., Б р о д ж и о М. и др. Оценка правомерности использования RAPD-анализа для выявления филогенетических связей между сортами яровой твердой пшеницы ( T. durum Defs.). Генетика, 2003, 39, 9: 1237-1246.
-
10. T o r r e s A.M., W e e d e n N.F., M a r t i n A. Linkage among sozyme, RFLP and RAPD markers in Vicia faba . Theor. Appl. Genet., 1993, 5: 937-945.
-
11. N e i M., L i W.-H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. of the Nat. Acad. of Sci. (USA), 1979, 76: 5269-5273.
-
12. V a n d e P e e r Y. Treecon for Windows: a software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Computer Application in the Biosciences, 1994, 10, 5: 569-570.
-
13. F e l s e n s t e i n J. Confidence limits in phylogenies an approach using bootstrap. Evolution, 1985, 39, 4: 783-791.
-
14. К о в е з а О.В., К о к а е в а З.Г., К о н о в а л о в Ф.А. и др. Использование RAPD-метода для исследования генома гороха ( Pisum sativum L.). Генетика, 2005, 41, 4: 1-8.
