Индикация представителей рода Trapa L. Приморского края по морфометрическим признакам плодов

Автор: Берестенко Е.Н., Кислов Д.Е.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Растениеводство
Статья в выпуске: 11, 2013 года.
Бесплатный доступ
На основе экспертных представлений и результатов статистического анализа изменчивости морфометрических признаков плодов рода Trapa L., произрастающих на территории Приморского края, построен ключ для их видовой дифференциации. Приводятся результаты машинной классификации плодов по общим морфометрическим показателям, дается оценка точности определения видов.
Семейство trapaceae, род trapa l, рогульник, систематика, приморский край, классификационный ключ
Короткий адрес: https://sciup.org/14082835
IDR: 14082835 | УДК: 582.885
Classification of genus Trapa L. species of the Primorskiy Krai by fruit morphometric features
On the basis of the expert representations and the statistical analysis results of the fruit morphometric feature variability of sort Trapa L. growing in the territory of Primorsky Krai, the key for their specific differentiation is developed. The fruit machine classification results on the general morphometric indices are resulted, the assessment of the sort definition accuracy is given.
Текст научной статьи Индикация представителей рода Trapa L. Приморского края по морфометрическим признакам плодов
Материалом для исследований послужили плоды перечисленных видов, собранные в ходе полевых работ на территориях Дальнереченского, Кировского, Пожарского, Спасского, Уссурийского, Ханкайского и
Хасанского районов Приморского края, а также в условиях культуры в Ботаническом саду-институте ДВО РАН [9, 12]. Для каждого плода осуществлялись измерения его морфометрических показателей. В зависимости от принадлежности вида к той или иной секции количество параметров плодов не оставалось постоянным. Перечень всех морфометрических признаков представлен в табл. 1 и на рис. 1.
Оценка дискриминационного потенциала морфометрических показателей плодов проводилась на основе традиционной и непараметрической схем дисперсионного анализа. При соответствии данных нормальному распределению (в качестве критерия согласия для проверки соответствия данных нормальному распределению использовался тест Шапиро-Уилка) и равенстве дисперсий использовалась традиционная схема однофакторного дисперсионного анализа Фишера, в противном случае, использовался непараметрический тест Крускала-Уоллиса [6].
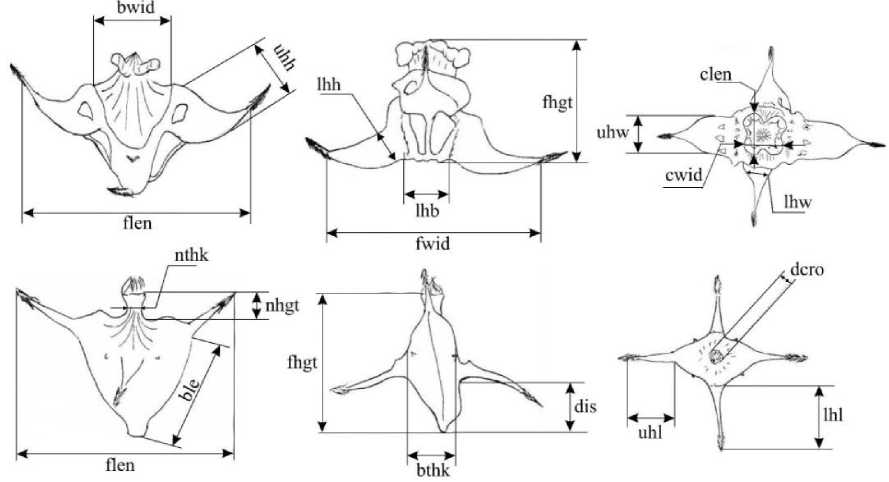
Рис. 1. Схема измерений морфометрических параметров плодов водяного ореха (вверху T. manshurica (секция Trapa), внизу T. nedoluzhkoi (секция Prototrapa))
Таблица 1
Условные обозначения морфометрических параметров плодов
|
Сокращение |
Описание признака |
Сокращение |
Описание признака |
|
flen |
Длина по верхним рогам |
uhh1 |
Высота верхних рогов |
|
fwid |
Ширина по нижним рогам |
uhh2 |
|
|
fhgt |
Высота плода |
uhw1 |
Ширина верхних рогов |
|
bthk |
Толщина тела плода |
uhw2 |
|
|
bwid |
Ширина тела плода |
lhl1 |
Длина нижних рогов |
|
ble1 |
Длина тела плода |
lhl2 |
|
|
ble2 |
lhh1 |
Высота нижних рогов |
|
|
clen |
Длина коронки |
lhh2 |
|
|
cwid |
Ширина коронки |
lhw1 |
Ширина нижних рогов |
|
dcro |
Диаметр коронки |
lhw2 |
|
|
nhgt |
Высота шейки |
lhb |
Расстояние между нижними рогами |
|
nthk |
Толщина шейки |
dis1 |
Расстояние от основания плода до основания нижних рогов |
|
uhl1 uhl2 |
Длина верхних рогов |
dis2 |
Примечание. Цифры 1 и 2 после сокращенного названия параметра означают минимальную и максимальную характеристику из двух симметричных.
Классификация представителей водяных орехов осуществлялась с использованием аппарата линейных классифицирующих функций, а визуальное представление объектов в факторном пространстве с привлечением линейного дискриминантного анализа [1]. Следует отметить, что предпосылки, определяющие оптимальность линейной классификации (нормальность, равенство корреляционных матриц) не выполнялись в рассматриваемых случаях. Последнее, однако, не являлось ограничением для использования схемы линейной классификации, учитывая, что верификация ее результатов может проводиться на базе имеющихся данных путем их предварительного разделения на тестовые и обучающие выборки.
Для проведения расчетов использовалась интегрированная посредством языка программирования Python (с использованием пакета Rpy2 ) вычислительная среда, включающая возможности статистического пакета R и библиотеки научных вычислений SciPy .
Результаты исследований и их обсуждение . Выполнение процедур статистического анализа осуществлялось на группах видов, предварительно разделенных в соответствии с принадлежностью их к той или иной секции, а также исходя из экспертных представлений о сходстве видов.
Состав сформированных групп имел следующий вид:
G1 – виды секции Prototrapa ( T. incisa , T. nedoluzhkoi , T. maximowiczii );
G3 – виды секции Trapa с четырехрогими плодами ( T. kozhevnikoviorum , T. manshurica (Ильинские озера), T. manshurica (оз. Малое Мраморное)).
В качестве примера рассмотрим процедуру определения наиболее информативных показателей в плане дискриминации видов для группы G2.
В результате такого анализа были определены наиболее информативные в плане дискриминации видов признаки. В частности, было установлено, что подходящими признаками для дифференциации видов являются высота верхних рогов (uhh1, 2), длина коронки (сlen), ширина коронки (cwid). Вместе с тем неприемлемым для разделения видов этой группы является такой признак, как расстояние от основания плода до основания нижних рогов (dis1, 2).
Результаты дисперсионного анализа для трех групп приведены в табл. 2. Первые три признака характеризуются наименьшим значением p-value и, следовательно, можно предположить, что они вносят больший вклад в разделение видов. Последним трем признакам свойственно наибольшее значение p-value, они характеризуются наименьшим вкладом в дискриминацию видов.
Дисперсионный анализ для групп G1 – G3
Таблица 2
|
G1 |
G2 |
G3 |
|||
|
Признак |
p-value |
Признак |
p-value |
Признак |
p-value |
|
flen |
4,1·10-74 |
uhh1 (uhh2) |
2,14·10-60 |
uhw2 (uhw1) |
8,57·10-40 |
|
ble2 |
3,2·10-73 |
clen |
1,04·10-59 |
uhh1 (uhh2) |
1,56·10-29 |
|
fhgt |
1.8·10-72 |
cwid |
2,56·10-56 |
bthk |
6,62·10-25 |
|
nthk |
9,5·10-23 |
lhb |
0,03 |
lhl1 |
0,17 |
|
lhh1 |
2,2·10-22 |
dis2 |
0,58 |
lhl2 |
0,08 |
|
lhl1 (lhl2) |
4,9·10-15 |
dis1 |
0,81 |
flen |
0,44 |
Следует отметить, что традиционные параметрические (дисперсионный анализ Фишера) и непараметрические (тест Крускала-Уоллиса) схемы множественного сравнения средних не позволяют в полной мере адекватно оценить дискриминационный потенциал морфометрических показателей. Это связано с тем, что по некоторым морфометрическим признакам в анализируемой группе видов могут одновременно состоять как сильно различимые по этому признаку виды, так и неразличимые. В связи с этим дополнительный анализ парных различий средних значений для видов, составляющих группу, является более предпочтительным.
Более конкретно судить о принципиальной возможности дифференциации видов по морфологическим параметрам плодов можно исходя из результатов метода «скользящего» (иностранный термин – leave-one-out cross validation) экзамена [1], который в данной работе применяется для оценки точности классификации, выполняемой на базе традиционной схемы линейного дискриминантного анализа в вычислительной среде R (функция lda библиотеки MASS).
Распределение исходных данных в проекциях на главные дискриминантные оси (рис. 2) визуально позволяет судить об успешности разделения видов водяных орехов по их общим (табл. 3) морфометрическим показателям.
Общие морфометрические признаки для групп G1–G3
Таблица 3
|
G1 |
G2 |
G3 |
|
ble1, ble2, bthk, bwid, dis1, dis2, fhgt, flen, fwid, lhh1, lhh2, lhl1, lhl2, lhw1, lhw2, nhgt, nthk, uhh1, uhh2, uhl1, uhw1, uhw2, uhl2 |
bthk, bwid, clen, cwid, dis1, dis2, fhgt, flen, fwid, lhb, lhh1, lhh2, lhl1, lhl2, lhw1, lhw2, nhgt, uhh1, uhh2, uhw1, uhw2 |
bthk, bwid, clen, cwid, dis1, dis2, fhgt, flen, fwid, lhb, lhh1, lhh2, lhl1, lhl2, lhw1, uhh1, uhh2, uhw1, uhw2, lhw2 |
Для групп G1–G3 были получены следующие оценки ошибок классификации:
G1: T. incisa – ошибка классификации 1,3 %, T. nedoluzhkoi – 0 %, T. maximowiczii – 2,5 %;
G2: каждый из видов идентифицируется абсолютно точно (ошибка 0 %);
G3: T. manshurica (Ильинские озера) – 34 %, T. manshurica (оз. Малое Мраморное) – 49 %, T. kozhevnikoviorum – 0 %.
Из рисунка 2 видно, что наилучшим образом по имеющимся признакам классифицируются представители группы G2. При этом наиболее значимыми для дискриминации признаками являются высота верхних рогов (uhh2, uhh1) и длина коронки (clen). Этим же признакам по результатам дисперсионного анализа свойственно наименьшее значение p-value.
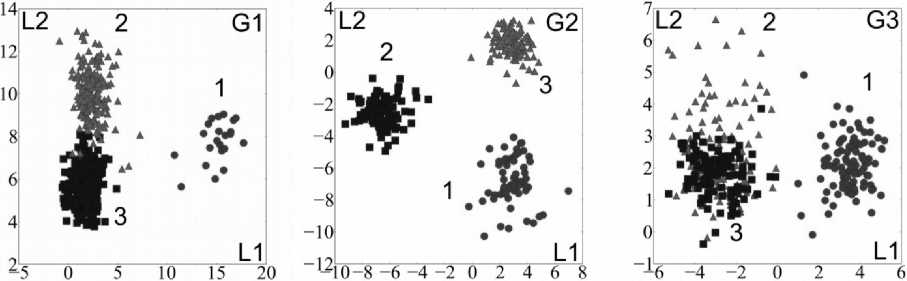
Рис. 2. Характеристики представителей групп G1-G3 в главных дискриминантных осях:
G1: 1 –T. Nedoluzhkoi; 2 – T. Maximowiczii; 3 – T. incisa; G2: 1 – T. Japonica; 2 – T. Khankensis;
Группа G3 также довольно четко характеризуется восстановлением видовой структуры: T. kozhevnikoviorum определяется как отдельный вид, а вот T. manshurica из разных мест произрастания практически неотличимы. Этот очевидный вывод о невозможности разделения представителей одного вида с различных мест обитания имеет важное значение в контексте проводимых исследований. При визуальном сравнении T. manshurica (Ильинские озера) и T. manshurica (оз. Малое Мраморное), учитывая качественные и морфологические особенности плодов, можно заключить, что представители данных видов вполне различны. Однако использование общих для представителей группы G3 морфометрических признаков (табл. 3) указывает на невозможность их определенной дифференциации, что, в частности, является свидетельством успешности выбора рассматриваемого набора морфометрических показателей. Тем не менее возможно ис- пользование более тонких методов, например, методов геометрической морфометрии, которые позволят провести дифференциацию представителей T. manshurica с различных мест обитания.
Если говорить о значимых для классификации признаках, то общими по результатам дискриминантного анализа и парных сравнений средних таковыми можно читать толщину тела плода (bthk) и ширину верхних рогов (uhw1 и uhw2). Но эти виды хорошо отличаются по такому морфологическому признаку, как нали-чие/отсутствие шейки. В связи с этим количественные признаки, определяемые статистическими расчетами как значимые для классификации, должны использоваться наряду с качественными.
Группа G1 характеризуется четким отличием T. nedoluzhkoi от двух других видов секции Prototrapa , которые в свою очередь представляют больший интерес для классификации. Поскольку плоды видов T. inсisa и T. maximowiczii при условии приблизительного равенства их метрических параметров практически неразличимы, вопрос о поиске наиболее информативных признаков очень актуален. Таковым признаком, однако, является не количественный, а качественный признак – наличие/отсутствие коронки. У T. inсisa коронка отсутствует, у T. maximowiczii она развита в той или иной степени. Но достоверно эти виды отличаются по особенностям вегетативных и генеративных органов – по форме листовых пластинок и окраске венчика.
Обобщением изложенного является приводимый ниже ключ для определения видов Trapa , произрастающих на территории Приморского края. В основу определения видов положены морфометрические и структурные признаки плодов, лишенных экзокарпия. Признаки вегетативных органов, в частности, листьев, а также цветков, приводятся лишь в тех случаях, когда их особенности однозначно указывают на тот или иной вид. В нашем случае это относится к двум видам T. incisa и T. maximowiczii . Пожалуй, это единственные из изученных виды флоры юга Дальнего Востока, различия между которыми наиболее достоверно проявляются в особенностях морфологии листьев и цветков, нежели плодов.
Ключ к определению видов Trapa
-
1. Плоды сравнительно мелкие, 12–31 мм длиной по верхним рогам и 5–18 мм высотой. Поверхность тела плода, лишенного экзокарпия, гладкая, скульптура не развита. Плоды 4-рогие, рога отчетливо отделяются от тела плода ……………………………………………………………………………………………………………..2
-
- Плоды среднего и крупного размера, 30–72 мм длиной и 11–32 мм высотой. Поверхность тела плода, лишенного экзокарпия, имеет обычно хорошо развитую скульптуру из бугров, вмятин и гребней. Плоды 2 или 4-рогие ...……….……………………………………………………………………………………………..………..……4
-
2. Нижние рога отходят практически от основания плода: расстояние от основания плода до основания нижних рогов 1–5 мм. Длина плода по верхним рогам больше ширины плода по нижним рогам.................3
-
-Нижние рога отходят от середины тела плода: расстояние от основания плода до основания нижних рогов 5–9 мм. Длина плода по верхним рогам меньше или приблизительно равна ширине плода по нижним рогам........................................................................................................................................…….……T. nedoluzhkoi
-
3. Шейка не расширяется в коронку. Листовые пластинки ромбические. Цветки розовые, мелкие (до 10 мм высотой). Плоды 12–22 мм длиной и 5–11,5 мм высотой .................................................................T. incisa
-
- Шейка с едва заметной или хорошо различимой коронкой. Листовые пластинки с усеченным основанием. Цветки белые. Плоды 17–31 мм длиной и 8–17 мм высотой.................................................T. maximowiczii
-
4. Плоды двурогие. Нижние рога отсутствуют либо представлены короткими тупыми выростами, лишенными шипиков.......................................................................................................................................................5
-
- Плоды четырехрогие. Нижние рога хорошо развиты, как правило, несколько меньше верхних, имеют шипики ……………………………….………………………………….………………………………………….…………….7
-
5. Плоды разнообразны по форме и величине. Шейка имеется высотой 1–5 мм. Коронка округлой, вытянутой или лопастной формы, 3–21 мм длиной и 2,5–20 мм шириной.............................................................6
-
- Рога утолщенные, высотой 3–20 мм. Шейка 2–12,5 мм толщиной. Коронка 4,5–21 мм длиной и 4–20 мм шириной, округлой, квадратной, лопастной формы. Скульптура плода, как правило, хорошо развита. Выросты на месте нижних рогов могут достигать в длину 10 мм. Плоды 33–72 мм длиной и 12–32 мм высотой…………………………………………………………………………………………………………...……….. T. Japonica
-
7. Шейка имеется. Коронка хорошо развита, лопастная, длина и ширина ее 5–14 мм. Верхние рога часто загнуты кверху …………………..….…..........................................................................................T. manshurica
-
-Шейка отсутствует. Коронка прямоугольная, лежит на теле плода, длина и ширина 5–11 мм. Верхние рога покатые, горизонтально направленные ….…………………………………………...……….T. kozhevnikoviorum
Интересно отметить результаты исследований, направленные на анализ точности предлагаемого ключа. Поскольку данные обучающих выборок, используемых в случае линейной классификации, содержали исключительно количественные морфометрические показатели плодов, а переходы по иерархической структуре ключа предполагают знание качественных признаков, было решено провести анализ в отношении «упрощенного» ключа, полученного из исходного исключением всех качественных условий (были исключены такие важные диагностические признаки, как наличие/отсутствие шейки, количество рогов и др.; ключевое выражение «Нижние рога отсутствуют …» было формализовано условием lhl2 < 5 мм). В результате тестирования работы «упрощенного» ключа было установлено, что вид T. kozhevnikoviorum определяется абсолютно точно, вид T. incisa – с ошибкой 8,7 %, T. manshurica – с ошибкой 18 %, T. maximowiczii – с ошибкой 23 %, T. nedoluzhkoi – с ошибкой 40 %, остальные – с ошибками более 50 %. Таким образом, даже в случае исключения всех качественных признаков при осуществлении переходов по классифицирующему дереву остается возможным успешное определение вида Trapa , по крайней мере, T. kozhevnikoviorum или T. incisa .
Заключение. При определении видов Trapa следует уделять первоочередное внимание не просто метрическим значениям тех или иных признаков, числовые диапазоны которых зачастую перекрываются, а информативным признакам. Наличие таких признаков позволяет избежать многочисленных измерений, тем самым оптимизируя и упрощая работу по определению видов. И здесь интересным является тот факт, что та или иная степень информативности свойственна таким структурным элементам плодов, как шейка и коронка. Именно на основании наличия/отсутствия шейки осуществляется разделение видов в секции Trapa , а по наличию/отсутствию коронки можно различить виды T. incisa и T. maximowiczii в секции Prototrapa .
В методологическом плане важно отметить то, что применение алгоритмов линейной классификации позволяет весьма точно идентифицировать видовую принадлежность водяных орехов. Таким образом, применение методов распознавания образов может быть эффективным при решении проблем систематики видов, характеризующихся выраженным полиморфизмом.
