Индуцируемые низкоинтенсивным красным светом защитные фотоэффекты

Автор: Пиняскина Е.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-3 т.12, 2010 года.
Бесплатный доступ
Получены данные о защитном действии низкоинтенсивного красного света на дрожжи, инактивированные оптическим излучением экологического диапазона. Показано, что фоторепарационные эффекты ФР680 реализуются без участия репарационных систем ДНК. Впервые показана возможность фотовосстановления повреждений, образующихся по фотодинамическому механизму в генетическом аппарате клетки с участием эндогенных фотосенсибилизаторов.
Ультрафиолет, дрожжи, репарационные системы днк, фотозащита, фотореактивация
Короткий адрес: https://sciup.org/148199042
IDR: 148199042 | УДК: 577.344.2:582.282.2
Protective photoeffects induced by low-intensive red light
Data about protective action of low intensive red light on yeast, inactivated by optical radiation in ecological range are obtained. It is shown, that photoreparation effects FP680 are realized without participation of DNA reparation systems. For the first time the opportunity of photorestoration of damages, formed on photodynamic mechanism in genetic apparatus of a cel with participation of endogenic photosensitizers is shown.
Текст научной статьи Индуцируемые низкоинтенсивным красным светом защитные фотоэффекты
Мониторинговые исследования состояния озонового слоя (как части атмосферы), проводимые в разных частях Земли, констатируют о прогрессирующем разрушении озонового слоя, связанном с антропогенным воздействием на окружающую среду. Вследствие этого увеличивается интенсивность проникновения в приземные слои атмосферы наиболее опасного вида УФ излучения – средневолнового (УФ-Б 280-320 нм), что ведет к целому ряду негативных последствий для человека: преждевременному старению, учащению случаев рака кожи (меланомы), катаракты, ослаблению иммунитета. Возникновение при воздействии УФ-излучения молекулярных повреждений ДНК, не устраняемых (или устраняемых не полностью) репаративными системами клетки, а также фотодеструкция белков и биомембран обусловливают развитие довольно многочисленных биологических эффектов. Согласно «Приложению I к Венской Конвенции по охране озонового слоя» в настоящее время приоритетными признаны исследования по изучению молекулярных принципов функционирования клеточных защитных механизмов, способных обеспечить устойчивость биосистем в условиях повышенного уровня радиации, способных защитить клетки от этих видов излучения [1].
Известные процессы репарации и фотоин-дуцированной защиты клеток, уменьшающие повреждающие эффекты УФ-излучения средне-(СУФ) и длинноволнового ультрафиолета (ДУФ), сходны в том, что активным в инициации этих процессов является свет видимого и/или ДУФ-диапазонов спектра [2-4] и направлены они на устранение либо предотвращение образования пиримидиновых димеров в ДНК. Ранее нами было показано [5], что защитить клетки от средневолнового УФ-облучения (290-320 нм) можно воздействием длинноволнового света с максимумом эффективности в красной области при 680 нм. Полученный нами спектр действия обнаруживал главный
максимум в красной области при 680 нм, небольшой максимум при 430-440 нм и имел достаточно сложную структуру (рис. 1) [5].
Рис. 1. Спектр действия фотоактивации клеток, инактивированных СУФ-излучением
Установленная форма дозовых кривых (рис. 2а и 2б) отличается от известных дозовых кривых (характерных для ферментативной фотореактивации), которые при достаточно высоких дозах синего света достигают уровня насыщения. Используя указанные различия, мы предприняли попытку разделить 2 типа фотореактивации, изменяя соответствующие режимы облучения синим светом, который активен как в ферментативной фотореактивации, так и в ФР680. Эти эксперименты мы проводили только на клетках S.cerevisiae , поскольку используемый в работе штамм дрожжей C.guilliermondii не способен к ферментативной фотореактивации, т.е. УФ-инактивированные клетки не восстанавливаются при пострадиационном действии больших доз синего света.
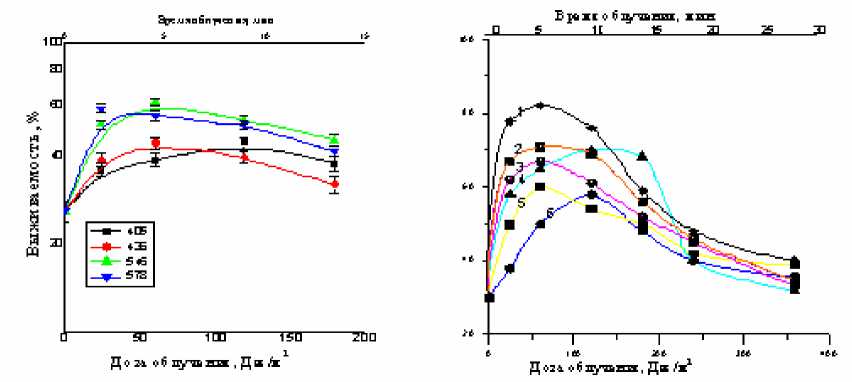
а) б)
Рис. 2: а) зависимость фотовосстановления СУФ-инактивированных клеток от дозы облучения монохроматическим светом в области 405-578 нм. Доза облучения 2,4 кДж/м2. Каждая точка отражает среднее значение из 6 экспериментов; б) зависимость фотовосстановления СУФ-инактивированных клеток от дозы (времени) облучения монохроматическим светом в области600-730 нм: 1 – 730 нм; 2 – 710 нм;
3 – 610 нм; 4 – 630 нм; 5 – 660 нм; 6 – 680 нм. Доза облучения – 2,4 кДж/м2
Опыты проводились по следующей схеме: сначала клетки облучали фиксированной дозой СУФ-света (2.4 кДж/м2), а затем подвергали их воздействию монохроматического света 405 нм разной интенсивности: ® 0,2 Вт/м2 в первые 30 мин и ~ 1,5 Вт/м2 при дальнейшем облучении этой же суспензии. Из полученных данных (рис. 3) отчетливо видно наличие 2 форм кривых фотореактивации: одной (при малых временах и дозах облучения – кривая 1), характерной для ФР 680 , и второй (кривая 2, имеющая насыщение при больших временах и дозах облучения), типичной для ферментативной фотореактивации. Важно подчеркнуть, что ФР 680 проявляется и при понижении температуры до 4 ° С во время действия фотореактивирующего света (рис. 2) (ферментативная фотореактивация в таких условиях как известно, не наблюдается [2, 4]). Этот факт может указывать на фотохимическую природу процессов, протекающих на начальной (световой) стадии ФР 680 .
Последующие эксперименты показали, что понижение температуры до 4 ° С в период между облучением клеток реактивирующим светом и их высевом на питательную среду приводит к постепенному снижению эффективности ФР 680 и к полному её исчезновению к 3 ч выдерживания. По-видимому, такой период времени необходим для завершения тех фотоиндуцированных биохимических процессов в клетке, которые в итоге приводят к реализации эффекта ФР 680 .
Рассмотренные выше данные свидетельствуют о наличии у дрожжей S.cerevisiae 2 различных фотореактивирующих систем, направленных на увеличение выживаемости СУФ-инактивиро-ванных клеток – ФР680 и ферментативной фотореактивации. Используя ранее найденые оптимальные режимы облучения, мы выявили эффект фотовосстановления ДУФ-инактивированных клеток. Установлено, что повышение уровня выживаемости таких клеток наблюдается при воздействии света всех использованных ранее длин волн в диапазоне 400-730 нм, причем максимальный эффект фотореактивации проявлялся при облучении клеток красным светом 680 нм.
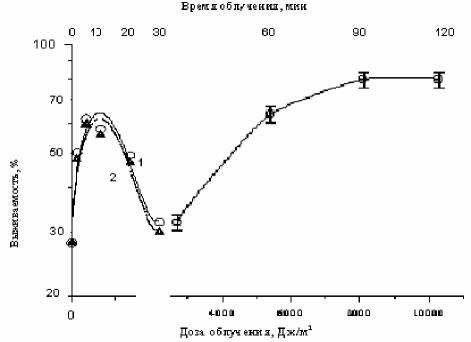
Рис. 3. Зависимость фотовосстановления СУФ-инактивированных клеток от дозы (времени) облучения монохроматическим светом 405 нм при 220С (кривая 1) и 40С (кривая 2). Доза СУФ-излучения – 2,4 кДж/м2 (≈ 30% выжываемости)
Кроме того, на эффективность фотореактивации (как и при ФР680 в случае действия СУФ-излучения) понижение температуры до 4°С во время облучения монохроматическим светом не влияет. Поскольку, СУФ- и ДУФ-индуцированные повреждения могут ликвидироваться системами репарации ДНК [6], из которых основными являются «темновые», (т.е. не нуждающиеся в свете) – эксцизионная и пострепликативная. Они менее специфичны, чем фотореактивация, и наряду с димерами способны устранять и другие повреждения структуры ДНК, однако механизм их работы сложнее и включает несколько последовательных стадий, контролируемых разными ферментами.
Нам представлялось целесообразным проверить, не связано ли действие обнаруженной нами защитной системы с фотоиндуцированной активацией этих репарационных систем. Для решения поставленного вопроса мы исследовали способность к фотореактивации (680) мутантных штаммов дрожжей ( S.cerevisiae ), дефицитных по эксцизионной ( rad 3-2 ) и пострепликативной ( rad 50-1 ) репарации ДНК. Предварительно было показано, что такие клетки более чувствительны к СУФ ( rad 3-2 ) и ДУФ ( rad 50-1 ) по сравнению с диким штаммом. Опыты по фотореактивации проводились по следующей схеме: клетки дикого штамма и мутантов rad 3-2 или rad 50-1 облучали фиксированной дозой СУФ (2,4 кДж/м2) или ДУФ (18 кДж/м2), после чего их подвергали воздействию монохроматического света 680 нм.
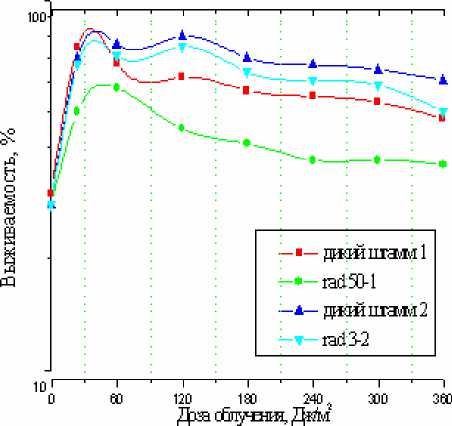
Рис. 4. Зависимость фотовосстановления СУФ- и ДУФ-инактивированных дрожжевых клеток от дозы (времени) облучения монохроматическим светом 680 нм
Как следует из полученных данных (рис. 4), фотореактивация мутантных штаммов наблюдается, причем ее эффективность примерно такая же, как и у дикого штамма. Эти данные могут указывать на то, что устранение СУФ-, ДУФ-индуци-рованных повреждений ДНК в процессе фотореактивации (680) осуществляется без участия эксцизионной и пострепликативной репарации. Очевидно, действие фотоиндуцированной защитной системы включает какой-то другой механизм ликвидации таких повреждений.
Выводы: обнаружение эффекта фотовосстановления при инактивирующем воздействии ДУФ-света является первым указанием на возможность фоторепарации повреждений, образующихся по фотодинамическому механизму в генетическом аппарате клетки с участием эндогенных фотосенсибилизаторов [7]. Проанализировав экспериментальные данные, мы пришли к выводу, что обнаруженная нами защитная система действует по общему первичному механизму, запускаемому одним клеточным фоторецептором. Данные о её высокой квантовой чувствительности, а также триггерном характере действия указывают на существование весьма эффективных механизмов биохимического усиления, которые инициируются первичным фоторецептором и способны обеспечить устойчивость клеток при инактивирующем действии оптического излучения различных длин волн экологического диапазона.
Список литературы Индуцируемые низкоинтенсивным красным светом защитные фотоэффекты
- Приложение I к Венской Конвенции об охране озонового слоя (Вена, 22 марта 1985 г.).
- Смит, К. Молекулярная фотобиология/К. Смит, Ф. Хэнеуолт. -М.: Мир. 1972. -272 с.
- Владимиров, Ю.А. Физико-химические основы фотобиологических процессов/Ю.А. Владимиров, А.Я. Потапенко. -М.: Высш. шк., 1989. -252 с.
- Конев, С.В. Фотобиология животной клетки/С.В. Конев, И.Д. Волотовский. -Л.: Наука, Ленингр.отд., 1979. -280 с.
- Фрайкин, Г.Я. Новая фотоиндуцибельная защитная система в клетках Candida guilliermondii при летальном действии средневолнового ультрафиолетового излучения/Г.Я. Фрайкин, Е.В. Пиняскина, М.Г. Страховская, А.Б. Рубин//ДАН. -1995. -Т. 343, № 2. -С. 265-267.
- Jagger, J. Near-UV radiation effects on microorganisms//Photochem. Photobiol. -1981. -V. 34. -P. 761-768.
- Пиняскина, Е.В. Действие красного света на дрожжи, инактивированные UVA-излучением//Вестник Российской Военной Академии. -СПб.: ВМедА, 2008. -Ч. 2. -С. 342.
