Информационная модель моносистемной организации высокогорных лугов (на примере Приэльбрусья)
")
Автор: Коломыц Э.Г., Цепкова Н.Л.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 1-1 т.12, 2010 года.
Бесплатный доступ
На основе информационно-статистических параметров геокомпонентных взаимосвязей установлены причинные механизмы пространственной организации высокогорных луговых экосистем, с количественной оценкой природных и антропогенных факторов, формирующих структуру травянистых сообществ и их продуктивность. Вскрыта системообразующая роль климатических и геоморфологических факторов, выявлены структурно-функциональные параметры пастбищной дигрессии высокогорных лугов.
Высокогорные экосистемы, луговые сообщества, пастбищная дигрессия, моносистемная организация, информационное моделирование
Короткий адрес: https://sciup.org/148198916
IDR: 148198916 | УДК: 581.52+581.524.4:574.4
Information model of the monosystem organization of alpine meadows (by the example of the by-Elbrus area)
The cause mechanisms of space organization of alpine meadow ecosystems have been determined on the base of information-statistical geocomponent interrelations. The quantitative valuation of nature and anthropogenic factors forming the grass community structure and meadow productivity have been carried out. The system-forming role of climatic and geomorphic factors have been revealed, the structure-functional parameters of pasture digression have been brought to light.
Текст научной статьи Информационная модель моносистемной организации высокогорных лугов (на примере Приэльбрусья)
результаты крупномасштабной ландшафтноэкологической съемки, проведенной авторами по методике, изложенной в [11, 12]. Всего было описано 72 пробных площади по наиболее типичным ландшафтным профилям. Съемка проводилась в 1983 г. (12-25 июля), т.е. в конце того периода современной истории России, когда в горных районах Большого Кавказа еще продолжалось интенсивное пастбищное использование земель.
Высокогорное пастбище «Жиласу», в пределах которого был заложен экспериментальный полигон, охватывает три высотных биоклима-тических пояса: субальпийский (абс. высоты 2150-2430 м), альпийский (2430-2820 м) и субнивальный (2650-2900 м). Пастбище используется под выпас мелкого и крупного рогатого скота. К середине сентября травостой выедался полностью. В июне-июле и сентябре выпас шел в субальпийском поясе, а в августе — в альпийском и субнивальном поясах.
ОБЩАЯ ХАРАКТЕРИСТИКА ЛУГОВЫХ ЭКОСИСТЕМ И СТЕПЕНИ ИХ ПАСБИЩНОЙ ДИГРЕССИИ
Территория экспериментального полигона представляет собой ступенчатое, полого наклонное эффузивное плато среднеплейстоценового возраста [8, 23]. Плато расчленено продольными эрозионно-тектоническими долинами, с невыработанными продольными профилями и скалистыми бортами, а также древними ледниковыми карами. Почвообразующим породами служат водно-ледниковые и пролювиальные отложения — гравийно-песчаные, щебнисто-суглинистые и галечные, с крупными валунами. На водоразделах и склонах тонкий плащ пролювия часто пронизан выходами коренных пород — вулканических туфо-брекчий.
Все разнообразие форм микрорельефа нами объединено в шесть типов, начиная от рассеивающих форм и кончая собирающими (см. рис. 4б). Наряду с этим, согласно ландшафтно- геохимической классификации [6], были выделены локальные типы местоположений (МП). Они образуют парагенетические ряды ландшафтных сопряжений (катен), включающие следующие звенья (в направлении от водораздела к местному базису эрозии): элювиальное (Э), трансэлювиальное (ТЭ), транзитное (Т), трансаккумулятивное (ТА), аккумулятивное (А) и супераквальное (Saq). Формы микрорельефа и типы местоположений объединяются одним общим понятием «геотоп», который характеризует геоморфологическую составляющую элементарного природного комплекса — ландшафтной фации, или биогеоценоза [13, 19]. Элювиальные геотопы мы относим к категории плакоров, по определению [4], и рассмат риваем произрастающие на них ненарушенные (коренные) или слабо нарушенные растительные сообщества как зональный представитель данного высотного пояса, в наибольшей степени отображающий фоновые климатические условия на данном интервале абсолютных высот.
В регионе распространены горно-луговые, сильно кислые, ненасыщенные основаниями и бедные азотом почвы [8]. Развиваясь на рыхлых водноледниковых и пролювиальных отложениях и на элювии эффузивных пород, они содержат значительное количество органических соединений (содержание гумуса до — 1520%), поэтому обладают высоким продукционным потенциалом, весьма привлекательным для выпаса.
Таблица 1. Ценотические группы высокогорных лугов, степень их пастбищной дигрессии, а также таксономические (средневзвешенные) нормы некоторых структурных и функциональных параметров
|
Ценотические группы, их дигрессионные стадии |
Структурно-функциональные параметры |
||||||
|
Проективное покрытие травостоя, % |
Средняя высота травостоя, см |
Проективное покрытие лишайников, % |
Мера разнообразия, бит |
Зеленая масса, ц/га |
|||
|
флористического |
ценоти-ческого |
сырая |
воздушносухая |
||||
|
Пустошные луга — ПЛ (ко-брезиево-лишайниковые, лишайни-ково-манжетково-кобрезиевые), Д о |
59.5 |
8.5 |
8.0 |
4.44 |
4.13 |
26.79 |
12.41 |
|
Альпийские ковры — АК (кобрезиево-манжетково-сиббальдиевые), Д 1—2 |
73.0 |
2.0 |
0 |
4.12 |
3.56 |
22.66 |
11.12 |
|
Плотнодернинно-манжетко-вые луга — ПМЛ (овсянице-осоково-манжетковые), Д 2 |
71.7 |
4.7 |
2.8 |
3.87 |
3.53 |
53.21 |
20.41 |
|
Манжетково-плотнодернин-инные луга — МПЛ (ман-жетково-осоково-злаковые, ра-знотравно-злаковые), Д 1 |
83.3 |
8.8 |
8.4 |
4.38 |
3.40 |
61.74 |
20.65 |
|
Бурьянистые сообщества — БС (чемерице-крапивные, сиббальдиевые), Д з |
70.9 |
35.3 |
39.9 |
3.49 |
2.92 |
88.41 |
21.89 |
|
Осоковые луга — ОС, Д о |
85.0 |
25.0 |
27.5 |
3.00 |
4.30 |
127.00 |
27.50 |
|
Пестроовсяницевые луга — ПОЛ (разнотравно-пестро-овсяницевые, злаковокрес-товниковые), Д о |
92.0 |
29.0 |
32.3 |
4.88 |
5.38 |
101.50 |
33.14 |
Таблица 2. Высотно-поясное распределение таксономических норм некоторых параметров
Высокогорно-луговых экосистем
|
Высотные пояса |
Абс. высоты, м |
Экосистемные параметры |
|||||||
|
Защебнен-ность поверхности почвы, % |
Температура почвы на глубине (оС) |
Влажность почвы на глубине 20-30 см, % |
Проективное покрытие лишайников, % |
Флористическое разнообразие, бит |
Зеленая масса, ц/га |
||||
|
сырая |
воз душно сухая |
||||||||
|
20 см |
40 см |
||||||||
|
Субальпийский |
2150—2290 |
16 |
10.9 |
10.4 |
36 |
4 |
3.50 |
84.85 |
26.77 |
|
2290—2330 |
26 |
15.8 |
15.4 |
30 |
16 |
4.02 |
74.82 |
25.00 |
|
|
2330—2400 |
21 |
15.4 |
14.3 |
33 |
22 |
4.24 |
54.71 |
27.04 |
|
|
2400—2500 |
44 |
13.9 |
11.9 |
32 |
31 |
4.64 |
59.55 |
18.97 |
|
|
Альпийский |
2500—2700 |
36 |
11.5 |
9.3 |
36 |
34 |
4.26 |
36.52 |
15.25 |
|
Субнивальный |
2700—2840 |
20 |
11.1 |
9.0 |
18 |
34 |
4.42 |
21.18 |
10.26 |
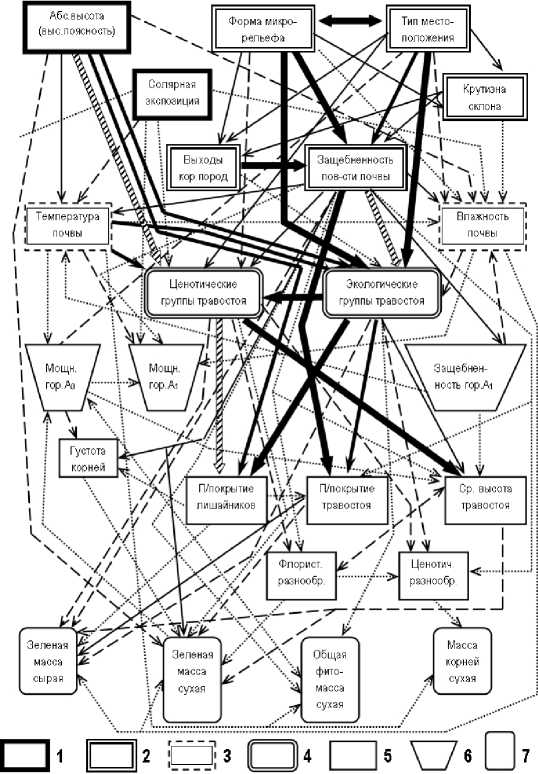
8 -—*9 --*10 --**11 ™^хг ^^13
Рис. 1. Общая информационно-статистическая модель пространственной организации высокогорнолуговых экосистем северного Приэльбрусья.
Блоки абиотических компонентов: 1 - независимых климатических; 2 - независимых и частично зависимых геоморфологических; 3 - гидротермических. Фитоценотические блоки: 4 - комплексный блок травянистых сообществ; 5 - структурный парциальный; 6 - функциональный парциальный. 7 - почвенный морфологический блок. Нормированные коэффициенты сопряженности признаков: 8 - 0.071-0.090; 9 - 0.091-0.110; 10 - 0.1110.140; 11 - 0.141-0.170; 12 - 0.171-0.200; 13 > 0.201.
Коренными типами травянистой растительности в исследуемом районе являются овсяни-цевые, кобрезиевые и осоковые луга, доминантами которых выступают овсяница пестрая и овсяница овечья , кобрезия волосовидная, изредка осока низкая, осока печальная [25], что особенно характерно для субальпийского пояса. В альпийском поясе эти фитоценозы сменяются альпийскими дерновыми лугами, альпийскими коврами и пустошными лугами. В субнивальном же поясе преобладают пустошные луга.
Описанные нами растительные ассоциации объединены (по степени участия различных видов) в семь цепотических групп (табл. 1). Кроме того, на основе известных отношений различных травянистых видов к условиями эдафического увлажнения, а также к температуре почвы [18] все растительные сообщества были разделены на пять экологических групп: 1) психрофиты (П); 2) ксерофиты (К); 3) ксе- ромезофиты (КМ); 4) мезофиты (М); 5) мезогигрофиты (МГ). Флористическое и ценотиче-ское разнообразия травостоя рассчитывались по известным формулам [26] на основе геоботанических описаний площадок. Первый параметр отражает общее число видов в данном сообществе, а второй - учитывает также обилие каждого вида.
Высокогорнолуговые экосистемы, находясь под тысячелетним воздействием человека, сформированы не только природными, но и антропогенными процессами, в данном случае пастбищным воздействием. В связи с этим при анализе необходимо было учитывать общую степень пастбищной дигрессии и выявлять ее основные геокомпонентные признаки. На основе соотношений в травостое ценных кормовых и плохо поедаемых видов, а также присутствия сорного разнотравья нами было выделено три стадии пастбищной дигрессии лугов (см. табл. 1).
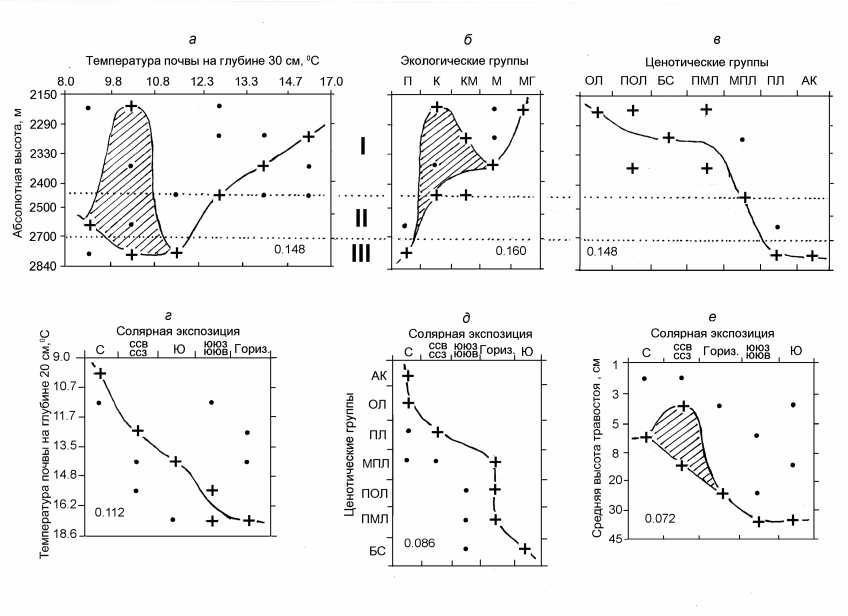
Рис. 2. Бинарная ординация температуры почвы, а также экологических и ценотических характеристик травостоя по факторам абсолютной высоты местности и солярной экспозиции склона (Обозначения экологических и ценотических групп см. в тексте и в табл. 1.)
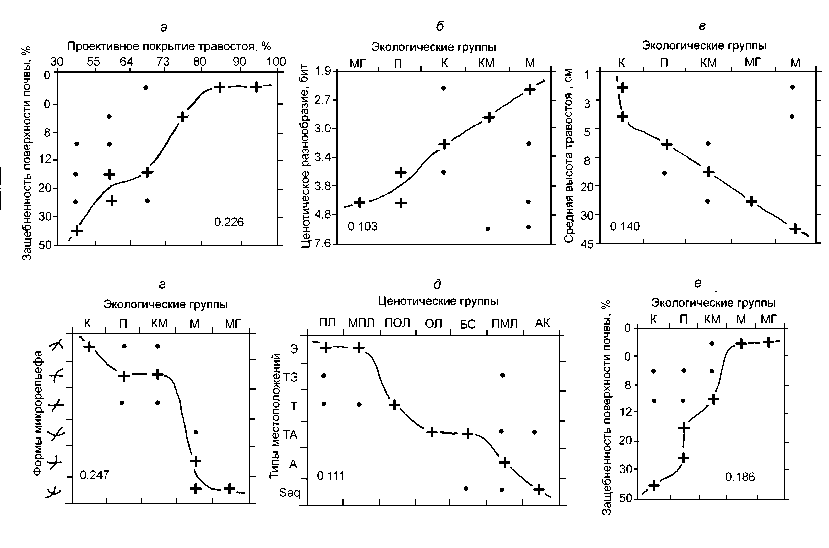
Рис. 3. Бинарная ординация экологических и ценотических характеристик травостоя между собой, а также по локальным геоморфологическим факторам.
Формы микрорельефа: 1 — гребни с ровным или выпуклыми продольным профилем; 2 — бровки террас и уступов с ровным поперечным профилем; 3 — равные участки склонов; 4 — ложбины с ровным тальвегом; 5 — вогнутые подножья террас и уступов с ровным поперечным профилем; 6 — кресловидные западины и ложбины (Обозначения типов местоположений см. в тексте.)
К ненарушенным (стадия Д0) отнесены луга, в которых доминантами и содоминантами являются виды, характерные для данного вы сотного пояса, а сорные виды практически отсутствуют. Состояние сообщества, где преобладают достаточно ценные и в то же время ус- тойчивые к выпасу растения, а в качестве субдоминантов выступает плохо поедаемое разнотравье, мы считаем устойчивым рабочим состоянием пастбищной экосистемы, находящемся в начальной дигрессионной стадии (Д1).
При чрезмерных нагрузках малоценные в кормовом отношении и в то же время устойчивые к вытаптыванию виды разнотравья увеличивают обилие. Состояние сообществ, где доминирование переходит к манжеткам, при сохранении в травостое плотнодернинных растений, мы считаем средней стадией дигрессии (Д2). Наконец, фитоценозы, состоящие из малоценных в кормовом отношении или вообще непоедаемых видов (чемерицы, сиббальдии), а также сообщества рудерального, бурьянистого разнотравья относятся к третьей дигрессионной стадии (Д з ).
МЕТОДЫ ИНФОРМАЦИОННОГО МОДЕЛИРОВАНИЯ
Информационный анализ геокомпонентных связей служит одним из инструментов моно-системного моделирования природных комплексов, согласно [15]. С помощью энтропийных мер [9] проводится количественная оценка пространственной сопряженности между раз личными природными компонентами, каждый из которых может быть представлен несколькими признаками. Нами были использованы два основных параметра информационностатистических связей [1, 16, 17]: нормированный коэффициент сопряженности К(А;В) явления А с фактором В (в каждой паре признаков) и частный коэффициент связи Ci,. По первому параметру построена общая информационная модель высокогорно-луговых экосистем на множестве выбранных признаков (рис. 1). Упорядоченная совокупность значений К(А;В) позволяет определить, что от чего зависит и насколько, и тем самым построить генеральную схему ландшафтно-экологических связей. По второму же параметру устанавливалась система экологических ниш каждого значения (градации) ai явления А в пространстве значений (b,) фактора В. Экологическая ниша данного геокомпонента или всего природного комплекса есть область его распространения в одном из пространств того или иного экологического фактора. Проведенный нами прямой градиентный анализ экологических ниш соответствует известному в геоботанике и ландшафтной экологии методу ординационного анализа [18, 21].
Таблица 3. Распределение таксономических норм некоторых параметров высокогорно-луговых экосистем по уровню эдафического увлажнения (экологическим группам травостоя)
Экосистемные параметры
|
Экологические группы травостоя |
Температура почвы на глубине |
Влажность почвы в слое, (%) |
Проективное покрытие травостоя, % |
Мера разнообразия, бит |
Зеленая масса, ц/га |
||||
|
сырая |
воздушносухая |
||||||||
|
флорис-тичес-кого |
ценоти-ческого |
||||||||
|
30 см |
40 см |
30-40 см |
40-50 см |
||||||
|
Психрофиты |
10.3 |
8.9 |
27 |
30 |
59 |
4.45 |
3.90 |
26.79 |
12.15 |
|
Ксерофиты |
14,1 |
13.2 |
19 |
15 |
52 |
4.20 |
3.07 |
32.93 |
10.43 |
|
Ксеромезофиты |
13,1 |
13.5 |
30 |
26 |
78 |
4.18 |
3.77 |
52.17 |
17.53 |
|
Мезофиты |
13,1 |
13.3 |
37 |
36 |
82 |
4.01 |
3.77 |
73.79 |
25.46 |
|
Мезогигрофиты |
8,9 |
7.6 |
55 |
56 |
85 |
3.00 |
4.30 |
127.00 |
27.50 |
Таблица. 4 Распределение таксономических норм некоторых экосистемных параметров по типам местоположений
|
Тип местоположения |
Экосистемные параметры |
||||||||
|
Выходы коренных пород, % |
Защебнен-ность поверхности почвы, % |
Температура почвы на глубине 30 см, ° C |
Влажность почвы в слое 30-40 см, % |
Проективное покрытие травостоя, % |
Мера разнообразия, бит |
Мощность Почвенно-гор. А о , см |
Зеленая сырая масса, ц/га |
||
|
флорис-тичес- кого |
ценоти-ческого |
||||||||
|
Элювиальный |
15 |
15 |
10.9 |
31 |
73 |
4.26 |
3.11 |
7.5 |
37.95 |
|
Трансэлювиальный |
33 |
26 |
12.7 |
21 |
52 |
4.25 |
3.61 |
4.4 |
32.11 |
|
Транзитный |
11 |
12 |
11.7 |
28 |
71 |
4.46 |
4.14 |
6.7 |
48.89 |
|
Трансаккумулятивный |
14 |
2 |
12.7 |
44 |
80 |
3.71 |
4.07 |
6.2 |
81.49 |
|
Аккумулятивный |
5 |
0 |
13.6 |
37 |
80 |
3.83 |
3.67 |
3.5 |
46.56 |
|
Супераква-льный |
13 |
2 |
13.5 |
37 |
83 |
3.81 |
2.94 |
3.4 |
79.76 |
В графически преобразованных матрицах значимых (>1) величин C i, по горизонтали идут градации явления, а по вертикали — градации фактора. Каждый вектор-столбец описывает экологическую нишу определенной градации явления в пространстве значений данного фактора. Градации фактора с наибольшими значениями частного коэффициента связи образуют некоторую область доминирования явления — экологический доминант (обозначен символом " + "), остальные же градации относятся к "размытой" части ниши (со знаком " • ", см. рис. 2 и др.).
Основную картину бинарной ординации явления по фактору дает кривая, проведенная через экологические доминанты явления. В соответствии с правилами информационного анализа [16] при наличии в столбце двух яче-ек-доминантов, разделенных одной незначимой ячейкой, кривая проводится по данной ячейке. Позиции «размытых» частей экологической ниши указывают на наличие возможных отклонений от доминанта, что позволяет отчетливо представить развертку объема ниши данного явления (как числа реализованных позиций вектора-столбца) в пространстве изменений состояний фактора. Чем шире ниша, тем менее чувствительно данное состояние явления к изменению фактора и в этом смысле оно более устойчиво. И наоборот, более узкая ниша (в предельном случае состоящая из одного доминанта, без «размытых» частей) указывает на повышенную детерминацию данного состояния явления тем или иным фактором. При наличии в векторе-столбце экологической ниши двух отстоящих друг от друга доминантов между ними образуется «зона» толерантности — неустойчивого равновесия (на графиках бинарной ординации такие «зоны» показаны штриховкой).
ГЕНЕРАЛЬНАЯ СХЕМА МОНОСИСТЕМНОЙ ОРГАНИЗАЦИИ ЛУГОВЫХ ПРИРОДНЫХ КОМПЛЕКСОВ
При построении общей информационной модели (см. рис. 1) направление связей определялось на основе существующих представлений об иерархии системообразующей роли различных природных компонентов и об их характерных временах [1, 2]. С одной стороны, чем больше входных стрелок к данному признаку и чем они крупнее, а с другой — чем меньше выходных стрелок и чем они мельче, тем сильнее выражены свойства этого признака как индикатора состояния экосистемы. Преобладание же выходных стрелок, особенно крупных, указывает на то, что данный признак играет роль преимущественно эдификатора , т.е. средообразователя.
На орграфе признаки расположены сверху вниз в пять рядов (пять экологических рангов)
— в порядке уменьшения их роли как ландшафтообразующих эдификаторов и роста их индикационных свойств, т.е. в направлении от независимых переменных к все более зависимым. Таким образом, эти схемы отражают упорядоченную иерархию взаимовлияния природных компонентов. Первый ряд образует плеяда первичных входных абиотических факторов — как климатических, так и геоморфологических. Во втором ряду помещены ценотические и экологические группы травостоя, отображающие, соответственно, структурный и режимный аспекты общей пространственной организации высокогорно-луговых сообществ. Как показано на орграфе, эти группы напрямую обусловлены множеством абиотических факторов. Сюда же примыкает третий, почвенно-морфологический ряд, признаки которого связаны преимущественно с литогенными условиями почвообразования.
Четвертый экологический ранг составляют различные парциальные структурные характеристики луговых сообществ, имеющие достаточно сильную сопряженность не только с це-нотическими и экологическими группами травостоя, но и с абиотическими условиями среды. На данном уровне заканчивается система наиболее значимых экологических связей ( К(А;В) > 0,140). Наконец, пятый, заключительный ряд образуют параметры основных фитомасс как дискретные показатели функционирования луговых экосистем. Эти показатели отличаются наиболее слабыми связями с вышестоящими по экологическому рангу природными компонентами, в том числе со структурными фитоценотическими признаками.
ПРИРОДНЫЕ МЕХАНИЗМЫ ФОРМИРОВАНИЯ ВЫСОКОГОРНОЛУГОВЫХ ЭКОСИСТЕМ
Влияние высотной климатической зональности. Анализ бинарной ординации начнем с высотно-зональных (фоновых) климатических и местных микроклиматических условий, которые представлены в информационной модели соответственно абсолютной высотой местности и солярной экспозицией склонов. Явным доминантом здесь выступает высотная климатическая зональность (поясность). Она определяет, прежде всего, термический режим почво-грунтов ( К(А;В) = 0,148). С ростом абсолютной высоты в интервале 2150 ^ 2840 м их температура на глубине 30 см ( 1 30) снижалась на 5—7о, хотя в нижней полосе субальп встречались и холодные почвы (рис. 2 а , табл. 2). Такой широкий спектр высотно-поясных значений термики высокогорных почв обеспечивает возможность появления в каждом поясе ин-тразоналъных , по определению [20], луговых сообществ под преломляющим воздействием локальных геоморфологических факторов (см.
ниже).
Почвенное влагосодержание связано с фоновыми климатическими условиями гораздо слабее ( К(А;В) = 0,072). В целом можно говорить о некоторой тенденции его снижения по мере подъема в горы. Однако повышенная влажность (например, в слое почвы 30-40 см от 30% до 65%) встречалась во всем изученном диапазоне абсолютных высот.
Высотно-зональнная дифференциация экологических и ценотических групп травостоя выражена весьма отчетливо (рис. 2 б, в ). С ростом высоты прослеживается общая тенденция смены более увлажненных эдафотопов (мезо-гигрофитных и мезофитных) менее увлажненными (ксеро-мезофитными и ксерофитными), что указывает на общий процесс ксерофитиза-ции луговой растительности. В альпийском же и субнивальном поясах на него накладывается также процесс психрофитизации, обусловленный низкими летними температурами. При этом травостой КМ и К экологических групп, произрастающий в условиях низкой влажности почв (для слоя почвы - 3040 см влажность W 30-40 = 10—20%), обнаруживает достаточно большой интервал высотной толерантности по всему субальпийскому поясу (см. рис. 2 б , табл. 2).
Высотные пояса мало отличаются друг от друга по флористическому разнообразию травостоя (см. табл. 2), что свидетельствует об определенном флорогенетическом единстве данного экорегиона. Однако ценотическое разнообразие весьма неоднородно. Наибольшими его значениями отличается средняя полоса субальп (2200-2300 м). С переходом же к альпийскому поясу ценотический спектр лугов резко обедняется. Это обусловлено, по-видимому, не только климатическими причинами, но и пастбищной дигрессией. В субальпийском поясе присутствуют все три дигрес-сионные стадии (см. выше), между тем как в альпийском и субнивальном поясах стадии Д2 и Д3 практически не встречаются.
Нижнюю полосу субальп, с наиболее благоприятными климатическими условиями, занимают разнотравно-пестроовсяницевые ассоциации (как высотно-зональный, коренной тип субальпийских лугов), а также осоковые, овся-ницево-осоково-манжетковые луга и бурьяни-стые сообщества — нитрофильные разнотравно-чемерице-крапивные луга (см. рис. 2 в ). При этом если коренные ассоциации сложены мезофитами и ксеромезофитами, а осоковые луга мезогигрофитны, то в умеренно дигрессионных плотнодернинно-манжетковых ассоциациях уже доминируют ксерофиты. ПМЛ совместно с бурьянистыми сообществами приурочены к наиболее теплым почвам ( 1 30 = 15—17о С), между тем как осоковые и пестроовсяницевые луга — к гораздо более холодным ( 1 30 = 8—11о С). В верхней полосе субальп (2400—2500 м)
на относительно теплых почвах ( 1 30 = 12-16оС) наиболее широко распространены слабо нарушенные выпасом ксеромезофитные манжетко-во-осоково-злаковые ассоциации (МПЛ), хотя коренными сообществами здесь уже выступают пустошные луга, сложенные почти исключительно психрофитами.
Альпийские и субнивальные поясы, с их наиболее низкими температурами ( 1 30 = 8-12оС, заняты преимущественно пустошными кобре-зиево-разнотравно-лишайниковыми лугами и альпийскими коврами — ксеро-мезофитными кобрезиево-разнотравно-сиббальдиевыми сообществами. В альпийском поясе встречаются также ассоциации МПЛ и ПМЛ.
Продукционные процессы в высокогорных лугах также предопределяются фоновыми климатическими условиями, хотя и в меньшей степени ( К(А;В) = 0,076-0,092), чем их фитоце-нотическая структура. Так, сырая зеленая масса ( В зел) по мере подъема в горы в целом уменьшается, и особенно резко в пределах субальпийского пояса — от 100-150 ц/га на высотах 2150-2130 м до 30-60 ц/га на отметках 2300-2500 м, где имеется определенная зона толерантности. В альпийском поясе разброс В зел весьма велик (в среднем от 12 до 48 ц/га), а субнивальный пояс отличается минимальными запасами сырой зеленой массы (не более 30 ц/га).
Экологическая роль локальных гидротермических контрастов. Фоновые климатические условия высокогорий преломляются местными микроклиматическими факторами. Среди них первостепенную роль играет солярная экспозиция склонов. Она определяет в первую очередь температурный режим почво-грунтов, что непосредственно сказывается на ценотиче-ской структуре луговых сообществ (рис. 2 г , д ). В направлении от теневых С, ССВ и ССЗ склонов к солнцепечным ЮЮЗ, ЮЮВ и Ю склонам, а затем к плакорам температура почвы на глубине 20 см ( 1 20) возрастала в среднем от 9,0—10,5оС до 16,5—18,5оС.
В соответствии с этим на плакорах формируется наиболее разнообразный спектр луговых ценозов, обусловленный как природными, так и антропогенными факторами (см. рис. 2 д ). Сюда входят группы ПОЛ, ПМЛ и МПЛ (см. табл. 1). Две альтернативные ценотические группы: осоковые ассоциации и альпийские ковры, занимающие крайние положения в высотнозональном ряду, тяготеют исключительно к теневым склонам. На склонах северной ориентации развиты также пустошные луга, свойственные альпийскому и субнивальному поясам.
Таким образом, локальные геоморфологические условия высокогорий, с их ярко выраженным солярным характером климата [3, 24], настолько сильно преломляют общий климатический фон, что в субальпах на теневых скло- нах возникают термоэдафотопы, аналогичные плакорным местообитаниям субнивального пояса. В то же время овсянице-осоково-манжетковые луга и бурьянистые сообщества, характеризующие стадии пастбищной дигрессии соответственно Д2 и Д3, оказываются наиболее свето- и теплолюбивыми, поскольку приурочены к субгоризонтальным поверхностям и склонам южной экспозиции.
Солярная экспозиция (СЭ) предопределяет также систему прямых и обратных связей между механическим составом почв, представленным в данном случае степенью защебненности поверхности почвы (ЗПП), влажностью почвы (ВП) и структурными характеристиками луговых сообществ — проективным покрытием и средней высотой травостоя (ППтр и Втр). Общая схема информационных потоков имеет следующий вид:
|
0.088 |
|||
|
k Jk |
|||
|
0.074 ПП ТР |
0.226 |
||
|
0.080 0.176 |
|||
|
СЭ |
З |
ПП В |
|
|
0.072 В ТР |
0.087 |
||
|
ц_____________ |
Г V |
||
|
0.073 |
|||
Здесь цифрами показаны значения параметра К(А;В). На солнцепечных склонах развивается наиболее высокий травостой (рис. 2 е ), с преобладающим ППтр = 75-100%, что и дает максимальную зеленую массу — 25-45 ц/га. В условиях же теневых склонов высота травостоя снижается в 5-10 раз, а его проективное покрытие сокращается до 65-50%, что вызывает 2,5-3-кратное уменьшение зеленой продукции (до 5— 10 ц/га).
Существенные отклонения от указанных доминирующих показателей, создающие весьма «размытые» экологические ниши параметра СЭ, вызваны пастбищной дигрессией, которая наиболее интенсивна на солнцепечных склонах, обладающих максимальными кормовыми ресурсами. На этих склонах и на плакорах процессы биохимического выветривания и почвообразования идут с наибольшей скоростью, и почвы здесь малощебнисты. Они содержат больше мелкозема с физической глиной и поэтому отличаются высоким влагосодержанием ( W2 0-30 = 30—65%). Эти факторы способствуют росту проективного покрытия травостоя (рис. 3 а ) и соответствующему увеличению продуктивности лугов.
На склонах же С, ССВ и ССЗ экспозиций гипергенные процессы развиты гораздо слабее, что подтверждается весьма низкими структурными и функциональными показателями лугов (см. рис. 2е), поэтому почвы здесь более ске-летны и примитивны. Низкое содержание мелкоземистых фракций не способствует накоплению почвенной влаги, которая просачивается в трещиноватые коренные породы. Почва остается не только холодной (см. рис. 2г), но и во многих случаях сухой (W30-40 <10-20%), и это существенно препятствует структурнофункциональному развитию луговых экосистем теневых склонов.
Весьма эффективным экологическим фактором локального порядка является состояние гидроэдафотопов, выражаемое экологическими группами травостоя. Прежде всего, с данным фактором тесно связаны сами ценотические группы (см. выше), что имеет далеко идущие последствия при пастбищной термоаридизации высокогорных почв (см. ниже).
От локальных гидротермических условий зависят также проективное покрытие травостоя (см. табл. 3) и его средняя высота, которые определяют запасы зеленой массы, т.е. продуктивность самого пастбища (см. рис. 1). Наиболее разреженный (ППтр = 30—55%) и низкий травяной покров свойственен ксерофитным лугам (рис. 3 в ). В психрофитных сообществах травостой немного выше, но его проективное покрытие колеблется в широких пределах (30— 80%). Наиболее высокие травы, с максимальным ППтр = 80-100%, развиваются в мезо-фитных и мезогигрофитных лугах. Ксеромезо-фитные сообщества имеют пониженную высоту травостоя при его высоком проективном покрытии (80-90%).
Наконец, следует отметить существенное влияние гидроэдафотопов на локальную дифференциацию продуктивности лугов (см. табл. 3). Наиболее продуктивны мезогигрофитные экотопы, приуроченные к аккумулятивным формам микрорельефа с наибольшим увлажнением. По запасам сухой зеленой массы к ним приближаются мезофитные луга. Ксерофиты же и психрофиты имеют одинаково низкую продуктивность.
Геоморфологическая локализация экосистемных параметров. Фоновые климатические условия высотных поясов существенно преломляются местными факторами: типом местоположения и формой микрорельефа, а также близостью к дневной поверхности коренных пород, определяющей степень скелет-ности почв. Эти факторы оказывают прямое дифференцирующее воздействие на влажность почвы и экологические группы луговых фитоценозов, причем в значительно большей степени, нежели фактор абсолютной высоты (см. рис. 1). Подтверждается известное в ландшафтной экологии положение [4, 13]: локальные экосистемные контрасты нередко существенно превышают размах их фоновой климатогенной дифференциации, что служит причиной формирования в каждом экорегионе так называемых региональных систем локальной зональности [12]. В данном случае такая система характеризуется экологическими и ценотиче-скими группами луговых фитоценозов, с их отдельными структурными и функциональными характеристиками.
Рельеф определяет прежде всего местное поле гидроэдафотопов, экологическая роль которых описана выше. Наиболее резко выражены контрасты между двумя экологическими плеядами лугов: с одной стороны, П, К и МК, а с другой, — М и МГ. Психро- и ксерофиты приурочены больше к ТЭ геотопам — гребням и бровкам террас (рис. 3г). На сравнительно сухих выпуклых и ровных склонах и на плако-рах преобладают ксеромезофиты (табл. 4). По нижним звеньям катен (ТА, А и Saq типам МП — ложбинам стока, подножьям крутых склонов и кресловидным формам) размещены мезофиты и мезогигрофиты, где почва в 1,5—2 раза влажнее. В свою очередь, увлажнение эдафотопов предопределяет формирование це-нотических групп высокогорных лугов (см. выше).
Микрорельеф не только преломляет фоновые климатические условия, но и регулирует пастбищную нагрузку на луга (см. ниже). В результате, имеем достаточно высокую сопряженность их ценотической структуры как с типами МП, так и с формами рельефа (рис. 3 д ). В нижней полосе субальпийского пояса (2150-2300 м) коренные разнотравно-пестроовсяницевые луга развиваются главным образом на ровных пологих склонах, осоковые и бурьянистые сообщества — по днищам ложбин и кресловин. Овсянице-осоково-манжетковые луга, имеющие более широкий высотный диапазон, тяготеют, скорее, к подножьям крутых склонов, хотя встречаются также на ТА и Saq геотопах (в западинах). Плотнодернинные осоково-злаковые фитоценозы, доминирующие у верхней границы субальп, приурочены к гребням и бровкам террас. Типичные для альпийского и субнивального поясов пустошные луга и альпийские ковры занимают два противоположных «полюса» катен: первые — Э умеренно влажные и ТЭ сухие гео-топы, вторые — Saq переувлажненные.
Интересно сопоставить общее и частное опосредованное (через гидроэдафотопы — ЭГ) влияние микрорельефа (МР) на формирование ценотических групп (ЦГ) высокогорных лугов. Схема информационных связей имеет вид с соответствующими значениями параметра К(А;В) :
МР
0.095
0.247
ЦГ ЭГ
0.367
Как видим, комплексный канал связи МР ^ ЦГ выражен очень слабо. По-видимому, обусловленные рельефом различные гидротермические факторы интерферируют с разным знаком, снижая суммарный эффект воздействия. Гораздо сильнее выражено парциальное влияние микрорельефа на ценотическую структуру лугов через эдафическое увлажнение. При этом фактор ЭГ оказывает на нее и самостоятельное влияние: значение параметра К(А;В) в паре ЭГ ^ ЦГ значительно больше, нежели в паре МР ^ ЭГ.
В условиях сильно расчлененного горного рельефа существенным экологическим фактором становится близость к дневной поверхно сти коренных пород, с чем тесно связаны за-щебненность поверхности почвы (ЗПП) и содержание литомасс (щебня) в самой почве (см. рис. 1 и табл. 4). Защебненность почвы воздействует главным образом на состояние гидроэдафотопов (рис. 3е). Несколько слабее выражено его влияние на ценотическую структуру лугов. При слабой защебненности (<5-10%) формируются М и МГ луговые сообщества, обладающие повышенным разнообразием ценотических групп (как ненарушенных, так и диг-рессионных), особенно в пределах субальпийского пояса (см. табл. 4).
Степень литоморфизации почвы обусловлена почти исключительно склоновыми процес- сами — главным образом солифлюкционным микротеррасированием склонов, которое приводит к разрывам дернины и обнажению коренных пород. По этой причине рост ЗПП от 0, до 50% сопровождается сменой экологических доминантов: мезофитов сначала на КМ, а затем и на К фитоценозы. При этом проективное покрытие травостоя сокращается с 80—100% до 30—50%, а покрытие лишайников возрастает от 0—10% до 35—50%. Начиная с уровня защеб-ненности 15—20%, ценотический набор луговой растительности резко обедняется. При ЗПП = 40—50% преобладают пустошные и манжетко-вые луга. В первом случае имеет место естественный уровень литоморфности почвы, а во втором — его явно дигрессионный характер. На второй стадии пастбищной дигрессии (Д2), когда ЗПП превышает 30%, происходит определенный скачок в антропогенной ксерофити-зация травяного покрова.
АНТРОПОГЕННЫЕ НАРУШЕНИЯ ВЫСОКОГОРНЫХ ЛУГОВ
Изменения гидротермического режима почв. Термоэдафотопы являются более существенным критическим фактором развития высокогорных лугов, нежели гидроэдафотопы (см. рис. 1). Экологические ниши сообществ по фактору влажности почвы сильно «размыты», причем преобладают отклонения от доминанта в сторону более низкой влажности почвы, что, несомненно, обусловлено пастбищной дигрессией лугов. Экологические группы фитоценозов сильнее связаны также с температурой почвы, нежели с ее влажностью. Наиболее велики температурные контрасты между мезогигрофитами и психрофитами, с одной стороны, и мезо- и ксерофитами, — другой (см. табл. 3). Характерно, что МГ, а иногда и М эдафотопы формируются в условиях низких температур почвы, приближающихся к температурам для психрофитов. В то же время К и КМ местообитаниям свойственны относительно высокие температуры почвы.
Ведущая экологическая роль термических условий отчетливо проявляется в высотнопоясной дифференциации двух основных групп коренных травянистых сообществ. Психрофит-ные пустошные луга альпийского и субнивального поясов развиваются в условиях наиболее холодных и сухих почв (ср. 130 < 9-10о С, W30-40 < 15-20%). Однако и в субальпах при локальном переувлажнении (W30_40 ~ 50-60%) встречаются столь же холодные эдафотопы, с мезогигрофитными осоковыми лугами. Почвы мезофитных разнотравно-пестроовсяницевых субальпийских лугов имеют умеренные температуры (ср. t30 = 10,5-12,5оС, редко до 14,0оС) и достаточно широкую зону толерантности по влажности (W30_40 = 25-60%). Таким образом, климатически обусловленная разница в летней температуре почвы между субальпийским и субнивальным поясами невелика — в пределах 3—4оС. Не столь значительными оказываются фоновые высотно-поясные контрасты и по влажности почвы.
Перевыпас вызывает резкое повышение температуры почвы. В верхних горизонтах почвы отчетливо выражены три положительных скачка температуры, соответствующие трем стадиям пастбищной дигрессии. Общий размах этих отклонений превышает максимальный фоновый перепад температуры между высотными поясами в регионе (рис. 4). В пределах субальп значительным повышение температуры почвы (итоговое A 1 30 > 4-5оС) вызвано антропогенной трансформацией ценотических групп (сменой дигрессионных стадий) по траектории ПОЁ(Д о ) ^ МПЛ(Д 1 ) ^ ПМЛ(Д 2 ) ^ БС(Д 3 ).
Одновременно происходит иссушение почвы. На траектории дигрессионных смен Д0 ^ Д 1 ^ Д2 ^ Д3 доминирующее значение влажности почвы в слое 20—30 см уменьшается с 45—65% до 10—20%, причем на бурьянистой стадии имеет место значительный разброс влажности (3—38%). На первых двух этапах иссушение почвы идет наиболее интенсивно (динамика таксономической нормы W 30 _ 40 = 55 ^ 34 ^ 26%).
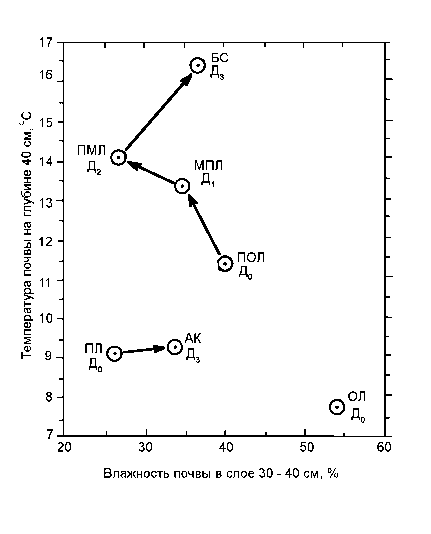
Рис. 4. Распределение ценотических групп травостоя, с их соответствующими дигрессионными стадиями (см. табл. 1), в поле средневзвешенных значений температуры почвы на глубине 40 см и влажности почвы в слое 30-40 см
Структурные преобразования луговых экосистем. Перевыпас вызывает смену самих экологических групп травостоя. Указанная выше дигрессионная траектория лугов сопровождается следующими переходами гидроэда-фотопов-доминантов: П, МГ ^ П, КМ ^ М, К ^ М, К. Мезофитная растительность сменяет- ся мезо-ксерофитной, а затем и ксерофитной (см. рис. 4). Лишь на бурьянистой стадии — влажность почвы несколько возрастает (норма Wзо-4о = 37%), что дает развитие вновь мезо-фитных, но уже рудеральных сообществ. Таким образом, пастбищная дигрессия субальпийских лугов (наиболее ценных в кормовом отношении для высокогорных районов) ведет к неизбежной термоаридизации почв и соответствующей ксерофитизации травостоя.
Аналогичная ситуация в отношении температуры складывается также в альпийском и субнивальном поясах. Почвы под альпийскими коврами, которые составляют здесь крайнюю степень пастбищной дигрессии (Д3) несколько теплее, чем под коренными пустошными лугами ( A 1 30 = 1,5-2,5оС по норме). Однако при этом они имеют более высокую влажность ( W 30-40 = 10—20 ^ 30-45%), что несколько напоминает дигрессионную смену ПМЛ(Д2) ^ БС(Д3) в субальпийском поясе.
Изменение продуктивности лугов. Пастбищная дигрессия снижает прежде всего проективное покрытие и среднюю высоту травостоя (см. табл. 1). В ненарушенном состоянии высокогорные луга имеют весьма широкий диапазон толерантности ППтр (от 30-50% до 90100%), что обусловлено описанными выше причинами локального порядка. Средняя высота травостоя также значительно варьирует, с доминирующими значениями 20—45 см. По мере усиления пастбищной дигрессии толерантная «зона» резко сужается и уже на стадии Д2 доминант ППтр сводится к 65-85%, а в состоянии Д3 снижается до 55-65%. При этом средняя высота травостоя резко падает уже на первой дигрессионной стадии — до 5-10 см, т.е. в 4-5 раз, что свидетельствует об интенсивном выедании травы скотом. В стадии Д3 травостой снижается до 1-3 см и только бурьянистые сообщества приобретают значительную высоту.
В итоге пастбищное нарушение травостоя приводит к общему снижению продуктивности лугов (см. табл. 1). В нижней полосе субальпийского пояса в ненарушенных разнотравно-пестроовсяницевых лугах (состоянии Д0) доминирующие значения сырой зеленой массы равны 100-150 ц/га в год. В пустошных лугах субнивального пояса они снижаются до 10-25 ц/га. При переходе пастбищ в дигрессионную стадию Д1 даже в субальпах производство зеленой массы падает до 30-55 ц/га, а в стадии Д2 уменьшается до 20-25 ц/га, т.е. приближается к естественной продуктивности субнивальных лугов. Таким образом, при чрезмерном выпасе пастбища приходят в деградированное состояние с одинаково низкой кормовой базой, независимо от климатических условий данного высотного пояса. Только рудеральные сообщества производят аномально большую зеленую массу (более 100 ц/га), которая не поедается скотом.
Пастбищная нагрузка проявляется в большей степени там, где луга дают наиболее продуктивны и с наилучшими кормовыми качествами. Производительность же лугов на локальном уровне тесно связана с влажностью почвы, которая, в свою очередь, определяется типом местоположения (см. табл. 4). В ксерофит-ных и ксеромезофитных лугах верхних и средних звеньев катен (Э, ТЭ и Т типы МП), с влажностью слоя почвы 20—30 см ( W 20-30) менее 15—20%, продуктивность сырой зеленой массы остается в пределах 1—20 ц/га, между тем как ТА и А геотопах, где W 20-30 = 55—60%, мезофитные и мезогигрофитные субальпийские сообщества дают до 100—150 ц/га и более.
Локальная дифференциация продуктивности лугов предопределяет их неодинаковую пастбищную нагрузку. Луга на геотопах первой указанной группы (особенно на узких гребнях, бровках террас и в верхних частях склонов) остаются почти в ненарушенном состоянии Д0, а сообщества плоских междуречных участков (плакоров) — в рабочем состоянии дигрессии Д 1 . Резким контрастом к ним выступают гораздо более производительные луга на геотопах второй группы (в нижних частях и у подножий склонов, на днищах ложбин водотоков). Здесь интенсивное выедание и вытаптывание травы приводит луговые экосистемы в крайнюю дигрессионную стадию (Д3). Несколько слабее (стадия Д2) нарушены переувлажненные Saq луга полузамкнутых западин и кресловин.
ЗАКЛЮЧЕНИЕ
Результаты проведенного моделирования позволили установить разномасштабные механизмы природных взаимодействий, формирующих пространственную организацию луговых экосистем высокогорий, а также дать количественную оценку разнообразному спектру их антропогенной динамики, которая вызвана лишь одной формой воздействия — пастбищной. По ранжированному множеству мер информационных связей смоделирована иерархическая структура межкомпонентных связей, которая раскрыла содержание известных представлений о высокогорных экосистемах как территориальных единствах с резко выраженной природной контрастностью и уязвимостью к внешним воздействием, с преобладанием внешних абиотических связей над связями внутренними, биотическими и со слабо выраженной автономностью биогеофизических режимов.
Список литературы Информационная модель моносистемной организации высокогорных лугов (на примере Приэльбрусья)
- Арманд А.Д. Информационные модели природных комплексов. М.: Наука, 1975. 126 с.
- Арманд А.Д., Таргульян В.О. Некоторые принципиальные ограничения эксперимента и моделирования в географии//Изв. АН СССР. Сер. геогр. 1974. № 4. С. 129-138.
- Вальтер Г. Растительность Земного Шара. Эколого-географическая характеристика. Т. III. М.: Прогресс, 1975. 428 с.
- Высоцкий Г.Н. Избранные труды. М.: Сельхозгиз, 1960. 435 с.
- Геренчук К.И., Топчиев А.Г. Информационный анализ структуры природных комплдексов//Изв. АН СССР. Сер. геогр., 1970. № 6. С. 132-140.
- Глазовская М.А. Геохимические основы типологии и методики исследований природных ландшафтов. М.: Изд-во МГУ, 1964. 230 с.
- Дьяконов К.Н. Информационный подход к анализу организации геосистем топологического уровня//Вопросы географии. Сб. 127. Моделирование геосистем. М.: Мысль, 1986. С. 111-122.
- Кавказ. Природные условия и естественные ресурсы СССР/Отв. ред. Н.В. Думитрашко. М.: Наука, 1966. 482 с.
- Кастлер Г. Азбука теории информации//Теория информации в биологии. М.: Изд-во иностр. лит., 1960. С. 9-53.
- Коломыц Э.Г. Информационно-статистический анализ структуры высокогорных экосистем и их антропогенных изменений//Тр. Высокогор. геофиз. ин-та. Вып. 58. М.: Гидрометеоиздат, 1984. С. 26-53.
- Коломыц Э.Г. Организация и устойчивость хвойнолесных экосистем на бореальном экотоне Русской равнины//Изв. РАН. Сер. геогр. 1995. № 3. С. 37-51.
- Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
- Крауклис А.А. Особенности географических градаций топического порядка//Топологические аспекты учения о геосистемах. Новосибирск: Наука, 1974. С. 87-137.
- Миркин Б.М. Теоретические основы современной фитоценологии. М.: Наука, 1985. 137 с.
- Преображенский В.С. О системе методов общей физической географии//Методы ландшафтных исследований. М.: Наука, 1969. С. 7-34.
- Пузаченко Ю.Г., Мошкин А.В. Информационно-логический анализ в медико-географиических исследованиях//Итоги науки. Медицинская география. Вып. 3. М.: ВИНИТИ, 1969. С. 5-74.
- Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: Системный анализ. М.: Наука, 1981. 275 с.
- Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971. 334 с.
- Сочава В.Б. Геотопология как раздел учения о геосистемах//Топологические аспекты учения о геосистемах. Новосибирск: Наука, 1974. С. 3-86.
- Спрыгин И.И. Материалы к познанию растительности Среднего Поволжья//Научное наследство. Т. 11. М.: Наука, 1986. 512 с.
- Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
- Урсул А.Д. Природа информации. Философский очерк. М.: Политиздат, 1968. 228 с.
- Федина А.Е. Ландшафтная структура Приэльбрусья. М.: Изд-во МГУ, 1971. 117 с.
- Хромов С.П., Мамонтова Л.И. Метеорологический словарь. Л.: Гидрометеоиздат, 1974. 568 с.
- Шифферс Е.Б. Растительность Северного Кавказа и его природные кормовые угодья. М.-Л.: Изд-во АН СССР, 1953. 400 с.
- Эшби У.Р. Введение в кибернетику. Пер. с англ. М.: Изд-во иностр. лит., 1959. 432 с.
