Инфузории (Ciliophora) планктона мелководной зоны водохранилищ Камского каскада
 планктона мелководной зоны водохранилищ Камского каскада")
Автор: Жариков Владимир Васильевич, Быкова Светлана Викторовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
Приводятся в сравнительном плане первые ориентировочные данные по составу, видовому разнообразию и количественным показателям свободноживущих инфузорий каскада водохранилищ Камы и Камской ветви Куйбышевского водохранилища. Анализируется характер изменений сообщества инфузорий планктона открытой литорали по продольной оси Камского каскада, начиная от незарегулированного верховья Камы до места слияния вод Камского и Волжского каскада водохранилищ. Указывается на тенденцию увеличения сверху вниз вдоль Камского каскада интегральных показателей видового разнообразия и волнообразное изменение параметров сообщества инфузорий планктона.
Инфузории, планктон, водохранилища, трансформация сообществ, видовое разнообразие
Короткий адрес: https://sciup.org/148201441
IDR: 148201441 | УДК: 574.52:593.17(282.247.415)
Planctonic ciliates (Ciliophora) of shallow zone from the Kama reservoirs cascade
This article presents data on the composition, species diversity and abundance of free-living ciliates from the Kama-river reservoirs and the Kama branch of Kuybyshev reservoir. The character of the changes in the ciliates plankton community of open littoral along the long axis of the Kama cascade is analysed. The tendency of increase in integrated parameters of species diversity as well as undulating change of abundance and biomass of ciliate plankton community down along the Kama reservoirs cascade are demonstrated.
Текст научной статьи Инфузории (Ciliophora) планктона мелководной зоны водохранилищ Камского каскада
И нтерес к исследованиям планктонных сообществ одноклеточных эукариот р. Камы и её притоков, собирающих воды с обширной горной и часто весьма заболоченной территории, продиктован полным отсутствием данных о видовом богатстве этих групп гидробиотов как до зарегулирования реки, так и после создания Камского каскада водохранилищ. В частности, данные о фауне и развитии свободноживущих инфузорий фактически до сих пор отсутствуют, принимая во внимание данные (всего 7 видов) 25-летней давности [8] по Нижнекамскому и Воткинскому водохранилищам. Первой попыткой заполнения этого «пробела» была маршрутная съемка в июле месяце 2004 г., проведенная В.В. Жариковым (не опубликовано) по всему Камскому каскаду, включая и его «речную» неза-регулированную часть. Основная цель нашей работы - получить представление о современном разнообразии свободноживущих инфузорий водохранилищ Камского каскада и выяснить, влияет ли современный гидрологический режим Камских водохранилищ на состав и структуру сообщества инфузорий планктона по продольной оси каскада для дальнейшей оценки вклада инфузорий водохранилищ Камы в биоразнообразие инфузорий каскада водохранилищ Средней и Нижней Волги. Полагаем, что полученные нами сведения положат начало более планомерному и тщательному исследованию свободноживущих инфузорий в данном районе.
РАЙОН ИССЛЕДОВАНИЯ И МЕТОДЫ
Исследования проводили в ходе маршрутной съемки в июле 2009 г., начиная с верховьев (п. Чепец) в незарегулированной части р. Камы и слияния с её притоком р. Вишера и далее – Камском, Воткинском, Нижнекамском водохранилищах и камской ветви Куйбышевского водохранилища (Камский и Волго-Камский плесы, до слияния с
Волжской ветвью). Протяженность маршрута исследования была чрезвычайно обширна и захватывала разные географические зоны: таежную и верхнюю границу лесостепной зоны. Морфометрические характеристики (объем, глубина, НПУ) водохранилищ Камы и их гидрологический режим заметно различаются (табл. 1). В составе водохранилищ Камского каскада лишь Нижнекамское имеет недельно-суточный режим регулирования стока.
Подробное описание физико-химических условий, химического состава воды и уровня продуктивности в период наших исследований в районе приведено ранее, в работе: [9]. Во время отбора проб в прибрежной части всех исследованных водохранилищ наблюдались гомотермия и, соответственно, аэробные условия на всех станциях. Цветность воды в целом снижалась от верховий Камского водохранилища вниз по каскаду. По уровню минерализации вода в камских водохранилищах соответствовала средне- и умеренно пресным водам [9]. Для Камского водохранилища было характерно чередование типов минерализации, связанное с поступлением низкоминерализованной гид-рокарбонатно-кальциевой воды из незарегулиро-ванной части р. Камы, стоков с болот, а также влиянием береговых выходов гипсов (сульфатный тип), месторождения калийных солей (хлоридно-кальциевый тип с повышенным содержанием натрия) и т.д. В нижележащих Воткинском, Нижнекамском и Куйбышевском водохранилищах преобладал кальций-гидрокарбонатный тип минерализации. Наибольшей продуктивностью обладали станции Воткинского водохранилища (эвтрофный и высокоэвтрофный уровень). Вместе с тем, по среднему содержанию фосфора прослеживался лишь слабо выраженный тренд увеличения его концентрации в направлении: Камское, Воткинское, Нижнекамское, Куйбышевское водохранилище (58, 61, 66 и 69 мг/м3, соответственно) [9].
Для исследования инфузорий отбор проб и их обработку осуществляли стандартными гидробиологическими и протозоологическими методами [3]. Для изучения горизонтального распределения ин- фузорий по водохранилищам каскада отбирали интегральные пробы (поверхность – дно) в прибрежной мелководной зоне, свободной от высшей водной растительности (глубина, в основном, до 3-4 м, реже до 7 м). Количественный учет проводили на материале, фиксированном сулемой. Видовую идентификации осуществляли с использованием цито-гистохимических методов (импрегнации серебром, окрашивания ядер по Фельгену). Для оценки биоразнообразия использовали коэффициенты видового разнообразия (индекс Шеннона, расчи-танные по численности (Hn) и биомассе (Hb), до- минирования (индекс Симпсона) и выровненности (индекс Пиелу). Ориентировочный расчет продукции инфузорий проводили «физиологическим» методом с внесением температурной поправки через коэффициент Вант – Гоффа Q10 = 2,25. Значение коэффициента К2 для инфузорий принимали равным 0,44. Для перехода от количества потребленного кислорода к тратам на обмен использовали оксикалорийный коэффициент = 4,86 кал/млО2. Калорийность сырого веса вещества инфузорий принимали равной 0,5 кал/мг [1, 6, 10].
Таблица 1. Основные морфометрические характеристики водохранилищ Камского каскада (по: [2, 7, 11])
Водохранилища
|
Параметры |
Камское |
Воткинское |
Нижнекамское |
Куйбышевское |
||
|
Камский плес |
Волгокамский плес |
в целом |
||||
|
НПУ, м |
108,5 |
89 |
62 |
- |
- |
53 |
|
Объем, км3 |
12,2 |
9,4 |
2,8 |
2,209 |
8,676 |
57,3 |
|
Площадь, км2 |
1915 |
1120 |
1000 |
539 |
1358 |
5900 |
|
Максимальная глубина, м |
30 |
28 |
14 |
- |
- |
41 |
|
Средняя глубина, м |
6,4 |
8,8 |
2,8 |
- |
- |
9,7 |
|
Длина, км |
300 |
365 |
270 |
200 |
65 |
510 |
|
Коэффициент водообмена |
4,2 |
5,7 |
6,6 |
- |
- |
4,2 |
|
Площадь мелководий,% |
19,4 |
11,3 |
49,8 |
- |
- |
16,8 |
|
Тип регулирования стока |
сезонный |
сезонный |
недельносуточный |
- |
- |
сезонный |
Примечание. «-» – отсутствие данных; НПУ – нормальный подпорный уровень
РЕУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Видовой состав инфузорий. Всего в открытом мелководье выявлено 102 вида инфузорий: в Камском водохранилище и незарегулированном участке р. Камы – 58 видов, в Воткинском – 63, в Нижнекамском – 65 и в камской ветви Куйбышевского водохранилища – 43 вида. Сходство видового состава инфузорий отдельных станций невелико (коэффициенты Серенсена – 47-49% в Камском; 3354% в Воткинском; 40-58% в Нижнекамском и 3436% в Камской ветви Куйбышевского водохранилища) и возрастает лишь на границах водохранилищ каскада, в районе плотин. Наибольшим сходством (Коэффициент Серенсена – 70%) отличается фауна инфузорий Воткинского и Нижне-Камского водохранилищ (рис. 1).
Из найденных 102 видов, ниже приведены лишь виды (табл. 2), входившие в состав доминантного (вклад в общую численность > 10%) и субдоминантного комплексов (> 1%).
Изменение видовой структуры: по каскаду водохранилищ проявляется ярче на уровне субдоминантов. Вместе с тем, в начале Камского каскада (Камское водохранилище) и в верховье Куйбышевского водохранилища (Камской ветви) – доминанты разные ( Codonella cratera , Rimostrobmidium humile и Tintinnopsis cylindrata , Balanion planctonicum, соответственно). В Воткинском и Нижнекамском водохранилищах в состав доминан-тов входят разные виды р. Urotricha и T. сylindrata .
А в незарегулированной части р. Камы – уже упомянутые вид К ща дополняю
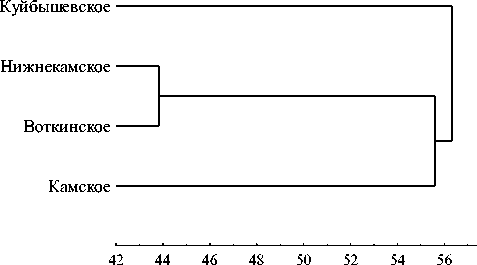
Рис. 1. Дендрограмма сходства фауны свободножи-вущих инфузорий водохранилищ Камского каскада и камской ветви Куйбышевского водохранилища
Для открытой литорали на всем каскаде массовыми (доля по численности >10 % и частота встречаемости > 25%) были следующие виды (табл. 2): разные виды р. Urotricha , мелкие виды р. Rimos-trombidium ( R. humile , R. hyalinum ); T. cylindrata , T. fluviatile , C. cratera , B. planctonicum , H. grandinella, P. natans . К редким (некоторые отмечаются впервые для водоемов Волжско-Камского каскада) можно отнести Leptopharynx costatus Mermod, 1914; Acineta flava ; Tintinnidium ephemeridium ; Membrani-cola tamari ; Zosterodasys sp., Staurophrya elegans
Таблица 2. Частота встречаемости, численность и вклад в общую численность доминантов (> 10%) и субдоминантов инфузорий (> 1%) в планктоне камских водохранилищ (июль 2009 г.)
|
Параметры |
Частота встречаемости, % |
Численность, тыс. экз/м3 |
Вклад от Nобщ.,% |
||||||||||||
|
Водохранилища Виды |
% 8 % |
=9 1 |
=9 =9 |
8 I Ei. |
=9 1 |
=9 =9 |
8 1 |
=9 1 |
=9 =9 |
9 |
|||||
|
Urotricha spp. (20-40 mk)* |
91 |
83 |
83 |
100 |
100 |
19,8 |
23,1 |
960,3 |
373,4 |
14,9 |
5,5 |
1,9 |
35,7 |
19,2 |
6,2 |
|
Rimostrobmidium spp.( R. humile (Penard, |
87 |
100 |
50 |
100 |
100 |
49,5 |
204,6 |
27 |
129,2 |
19,8 |
13,8 |
17,2 |
1,0 |
8,2 |
|
|
1922) + R. hyalinum (Mirabdulaev, 1985) |
|||||||||||||||
|
Urotricha spp.(< 20 mk) |
83 |
67 |
67 |
100 |
100 |
1,7 |
7,4 |
160,1 |
207,9 |
14 |
0,5 |
0,6 |
6,0 |
10,7 |
5,8 |
|
Tintinnidium fluviatile (Stein, 1863) |
74 |
67 |
83 |
100 |
25 |
1,7 |
38,8 |
35,8 |
41,5 |
5 |
0,5 |
3,3 |
1,3 |
2,1 |
2,1 |
|
Tintinnopsis cylindrata Kof. & Cam., 1892 |
74 |
50 |
67 |
100 |
75 |
1,7 |
14,9 |
11,6 |
315,9 |
28,9 |
0,5 |
1,3 |
0,4 |
16,2 |
12,0 |
|
Halteria grandinella (O.F. Muller, 1773) |
70 |
100 |
67 |
86 |
0 |
28,1 |
57,8 |
53,4 |
63,2 |
0 |
7,8 |
4,9 |
2,0 |
3,2 |
0 |
|
Pelagovorticella natans (Faure-Fremiet, 1924) |
65 |
67 |
100 |
43 |
50 |
1,7 |
28,9 |
124,9 |
111,3 |
9,1 |
0,5 |
2,4 |
4,6 |
5,7 |
3,8 |
|
Vorticella spp. |
65 |
67 |
100 |
29 |
75 |
1,7 |
17,3 |
172,2 |
18,4 |
5 |
0,5 |
1,5 |
6,4 |
0,9 |
2,1 |
|
Codonella cratera (Leidy, 1887) |
61 |
100 |
50 |
57 |
25 |
49,5 |
530,5 |
8,3 |
27,8 |
0,8 |
13,8 |
44,6 |
0,3 |
1,4 |
0,3 |
|
R. lacustris Petz & Foissner, 1992 |
61 |
83 |
33 |
86 |
25 |
23,1 |
52,8 |
30,8 |
46,7 |
0,8 |
6,5 |
4,4 |
1,1 |
2,4 |
0,3 |
|
Spathidium viride Kahl, 1926 |
61 |
50 |
83 |
71 |
25 |
0 |
6,6 |
47,3 |
38,2 |
0,8 |
0 |
0,6 |
1,8 |
2,0 |
0,3 |
|
Askenasia spp. |
57 |
67 |
50 |
86 |
0 |
1,7 |
14,9 |
106,2 |
19,3 |
0 |
0,5 |
1,3 |
4,0 |
1,0 |
0,0 |
|
Balanion planctonicum Foissner et al., 1994 |
57 |
50 |
33 |
57 |
100 |
1,7 |
2,5 |
3,3 |
5,7 |
24,8 |
0,5 |
0,2 |
0,1 |
0,3 |
10,3 |
|
Urotricha spp. (40-50 mk) |
57 |
50 |
50 |
71 |
50 |
3,3 |
4,1 |
263,5 |
34,9 |
5,8 |
0,9 |
0,3 |
9,8 |
1,8 |
2,4 |
|
Paradileptus conicus Wenrich, 1929 |
52 |
50 |
50 |
71 |
25 |
0 |
3,3 |
22,6 |
12,3 |
3,3 |
0 |
0,3 |
0,8 |
0,6 |
1,4 |
|
Cyclotrichium viride |
48 |
17 |
50 |
71 |
50 |
0 |
2,5 |
17,6 |
25 |
5,8 |
0 |
0,2 |
0,7 |
1,3 |
2,4 |
|
Didinium spp. |
48 |
17 |
50 |
71 |
50 |
0 |
2,5 |
2,8 |
45,3 |
10,7 |
0 |
0,2 |
0,1 |
2,3 |
4,5 |
|
Phascolodon vorticella |
48 |
33 |
67 |
57 |
25 |
0 |
3,3 |
103,4 |
28,8 |
1,7 |
0 |
0,3 |
3,8 |
1,5 |
0,7 |
|
Bursellopsis sp.1 |
43 |
33 |
67 |
57 |
0 |
1,7 |
0,8 |
11 |
33,9 |
0 |
0,5 |
0,1 |
0,4 |
1,7 |
0 |
|
Hypotrichia spp. 1 |
43 |
33 |
83 |
43 |
0 |
0 |
1,7 |
6,6 |
84,4 |
0 |
0 |
0,1 |
0,2 |
4,3 |
0 |
|
Limnostrombidium pelagicum (Kahl,1932) |
43 |
67 |
50 |
29 |
25 |
1,7 |
45,4 |
20,4 |
6,6 |
5 |
0,5 |
3,8 |
0,8 |
0,3 |
2,1 |
|
Pelagostrombidium falla x (Zach., 1895) |
43 |
50 |
50 |
57 |
0 |
5 |
5 |
102,3 |
8 |
0 |
1,4 |
0,4 |
3,8 |
0,4 |
0 |
|
Coleps hirtus viridis Ehrenberg, 1831 |
39 |
50 |
67 |
14 |
25 |
54,5 |
1,7 |
7,2 |
1,4 |
8,3 |
15,2 |
0,1 |
0,3 |
0,1 |
3,5 |
|
Enchelys simplex Kahl, 1926 |
39 |
50 |
50 |
14 |
50 |
1,7 |
3,3 |
20,9 |
0,5 |
14 |
0,5 |
0,3 |
0,8 |
0 |
5,8 |
|
Epistylis procumbens Zacharias, 1897 |
39 |
50 |
83 |
0 |
25 |
47,9 |
2,5 |
33,6 |
0 |
1,7 |
13,4 |
0,2 |
1,3 |
0 |
0,7 |
|
Lagynophrya sp. |
39 |
33 |
50 |
57 |
0 |
0 |
5 |
3,9 |
18,9 |
0 |
0 |
0,4 |
0,1 |
1,0 |
0 |
|
Pelagohalteria viridis (Fromentel, 1876) |
35 |
67 |
17 |
14 |
50 |
3,3 |
3,3 |
0,6 |
21,2 |
4,1 |
0,9 |
0,3 |
0 |
1,1 |
1,7 |
|
Foissner, Skogstad & Pratt, 1988 |
|||||||||||||||
|
Askenasia chlorelligera Krainer & Foissner, 1990 |
30 |
50 |
33 |
29 |
0 |
5 |
9,1 |
18,2 |
4,2 |
0 |
1,4 |
0,8 |
0,7 |
0,2 |
0 |
|
Cyclidium spp. 1 |
30 |
50 |
50 |
0 |
25 |
24,8 |
6,6 |
3,3 |
0 |
10,7 |
6,9 |
0,6 |
0,1 |
0 |
4,5 |
|
Haplocaulus sp. |
30 |
17 |
83 |
0 |
25 |
0 |
2,5 |
18,7 |
0 |
0,8 |
0 |
0,2 |
0,7 |
0 |
0,3 |
|
Sphaerophrya sp. |
30 |
33 |
67 |
0 |
25 |
0 |
9,9 |
7,7 |
0 |
0,8 |
0 |
0,8 |
0,3 |
0 |
0,3 |
|
Cyclidium spp. 2 |
26 |
67 |
0 |
29 |
0 |
1,7 |
3,3 |
0 |
11,8 |
0 |
0,5 |
0,3 |
0 |
0,6 |
0 |
|
Disematostoma spp. |
26 |
17 |
50 |
29 |
0 |
1,7 |
0 |
12,7 |
3,3 |
0 |
0,5 |
0 |
0,5 |
0,2 |
0 |
|
Monodinium chlorelligerum Krainer,1995 |
26 |
0 |
50 |
29 |
25 |
0 |
0 |
31,9 |
2,8 |
0,8 |
0 |
0 |
1,2 |
0,1 |
0,3 |
|
P. mirabile (Penard,1916) |
26 |
0 |
33 |
57 |
0 |
0 |
0 |
12,7 |
5,2 |
0 |
0 |
0 |
0,5 |
0,3 |
0,0 |
|
Cinetochilum margaritacium Perty, 1852 |
22 |
0 |
17 |
29 |
50 |
0 |
0 |
0,6 |
6,1 |
1,7 |
0 |
0 |
0 |
0,3 |
0,7 |
|
Didinium chlorelligerum Kahl, 1935 |
22 |
0 |
33 |
43 |
0 |
0 |
0 |
3,3 |
71,7 |
0 |
0 |
0 |
0,1 |
3,7 |
0 |
|
Hypotrichidium conicum Ilowaisky, 1921 |
22 |
0 |
67 |
0 |
25 |
0 |
0 |
14,3 |
0 |
1,7 |
0 |
0 |
0,5 |
0 |
0,7 |
|
Acaryophrya spp. |
17 |
0 |
50 |
14 |
0 |
0 |
0 |
28,6 |
1,4 |
0 |
0 |
0 |
1,1 |
0,1 |
0 |
|
Colpoda spp. |
17 |
0 |
33 |
14 |
25 |
0 |
0 |
29,7 |
0,9 |
0,8 |
0 |
0 |
1,1 |
0,0 |
0,3 |
|
Enchelys spp. |
17 |
0 |
33 |
14 |
25 |
0 |
0 |
9,4 |
0,5 |
7,4 |
0 |
0 |
0,3 |
0,0 |
3,1 |
|
Membranicola tamari Foissner et al, 1999 |
17 |
0 |
17 |
43 |
0 |
0 |
0 |
3,3 |
6,6 |
0 |
0 |
0 |
0,1 |
0,3 |
0 |
|
Mesodinium pulex (Clap.et L., 1859) |
17 |
0 |
33 |
29 |
0 |
0 |
0 |
6,1 |
1,4 |
0 |
0 |
0 |
0,2 |
0,1 |
0 |
|
Monodinium sp. |
17 |
0 |
17 |
43 |
0 |
0 |
0 |
2,8 |
47,1 |
0 |
0 |
0 |
0,1 |
2,4 |
0 |
|
Rabdoaskenasia minima Kr. & Foiss., 1990 |
17 |
0 |
50 |
14 |
0 |
0 |
0 |
6,6 |
4,7 |
0 |
0 |
0 |
0,2 |
0,2 |
0 |
|
Tintinnidium ephemeridium Hillard, 1968 |
17 |
0 |
50 |
14 |
0 |
0 |
0 |
3,9 |
0,9 |
0 |
0,1 |
0 |
0 |
||
|
Pelagovorticella mayeri (Faure-Fremiet, 1920) |
17 |
17 |
50 |
0 |
0 |
0 |
2,5 |
15,4 |
0 |
0 |
0 |
0,2 |
0,6 |
0 |
0 |
|
Bursellopsis sp. 2 |
13 |
0 |
33 |
0 |
25 |
0 |
0 |
23,7 |
0 |
0,8 |
0 |
0 |
0,9 |
0 |
0,3 |
|
Chilodonella sp. |
13 |
0 |
0 |
14 |
50 |
0 |
0 |
0 |
0,9 |
1,7 |
0 |
0 |
0,0 |
0 |
0,7 |
|
Pseudohaplocaulus infravacuolatus Foissner |
13 |
17 |
33 |
0 |
0 |
0 |
37,1 |
24,8 |
0 |
0 |
0 |
3,1 |
0,9 |
0 |
0 |
|
& Brozek, 1996 |
|||||||||||||||
|
R. velox (Faure - Fr., 1924) |
13 |
33 |
0 |
0 |
25 |
6,6 |
0,8 |
0 |
0 |
2,5 |
1,8 |
0,1 |
0 |
0 |
1,0 |
|
Strongylidium sp. |
13 |
33 |
17 |
0 |
0 |
1,7 |
0,8 |
1,1 |
0 |
0 |
0,5 |
0,1 |
0 |
0 |
0 |
|
Podophrya sp. |
13 |
0 |
33 |
0 |
25 |
0 |
0 |
3,9 |
0 |
0,8 |
0 |
0 |
0,1 |
0 |
0,3 |
|
Acineta flava Kellicott, 1885 |
9 |
0 |
0 |
14 |
25 |
0 |
0 |
0 |
0,5 |
1,7 |
0 |
0 |
0 |
0 |
0,7 |
|
Aspidisca lynceus (O.F. Muller, 1786) |
9 |
0 |
0 |
29 |
0 |
0 |
0 |
0 |
28,8 |
0 |
0 |
0 |
0 |
1,5 |
0 |
|
Ctedoctema acanthocrypta Stokes, 1884 |
9 |
17 |
0 |
0 |
25 |
3,3 |
0 |
0 |
0 |
0,8 |
0,9 |
0 |
0 |
0 |
0,3 |
|
Phialina sp. |
9 |
0 |
0 |
14 |
25 |
0 |
0 |
0 |
0,9 |
1,7 |
0 |
0 |
0 |
0 |
0,7 |
|
Spathidium sp. |
9 |
17 |
0 |
0 |
25 |
0 |
0,8 |
0 |
0 |
2,5 |
0 |
0,1 |
0 |
0 |
1,0 |
|
Urotricha spp.(> 50 mk) |
9 |
0 |
0 |
0 |
50 |
0 |
0 |
0 |
0 |
7,4 |
0 |
0 |
0 |
0 |
3,1 |
|
Epicarchesium pectinatum (Zacharias, 1897) |
4 |
17 |
0 |
0 |
0 |
0 |
7,4 |
0 |
0 |
0 |
0 |
0,6 |
0 |
0 |
0 |
|
Colpidium sp. |
4 |
17 |
0 |
0 |
0 |
1,7 |
0 |
0 |
0 |
0 |
0,5 |
0 |
0 |
0 |
0 |
|
Cyrtolophosis sp. |
4 |
0 |
17 |
0 |
0 |
0 |
0 |
3,3 |
0 |
0 |
0 |
0 |
0,1 |
0 |
0 |
|
Strobilidium caudatum (Fromentel, 1876) |
4 |
17 |
0 |
0 |
0 |
0 |
0,8 |
0 |
0 |
0 |
0 |
0,1 |
0 |
0 |
0 |
Прим.: * - виды расположены в порядке убывания частоты встречаемости по каскаду; жирным шрифтом выделены значения численности вида, доминировавшего (> 10%) в отдельных пробах ; р. Кама – незарегулированный участок р. Камы, включая и р. Вишеру, КамВ – Камское водохр., ВВ – Воткинское водохр., НКВ – Нижнекамское водохр.; КВ – Камская ветвь Куйбышевского водохр.
Zacharias , 1893; Nassulopsis elegans (Ehrb., 1833), Pelagovorticella mayeri , Pseudohaplocaulus in-fravacuolatus ; Cristigera pleuronemoides Roux,1901; Sphaerophrya sp . ; Rhabdoaskenasia minima Krainer& Foissner, 1990; Chilodontopsis vorax (Stokes, 1887).
Наши предварительные исследования водохранилищ Камского каскада позволяют предполагать, что с момента последних исследований (1987 г.) за истекшие 25 лет среди массовых видов [8] произошли некоторые изменения: нами не обнаружена Stokesia vernalis Wenzich, 1929; вероятна и замена видов среди олиготрих (например, замена Limnos-trombidium (= Strombidium) viride (Stein, 1932) Krainer, 1995 на Pelagostrombidium sp. и Rimostrom-bidium (= Strobilidium ) velox (Faure-Fr., 1924) Jankowski, 1978 на Rimostrombidium lacustris Petz & Foissner, 1992).
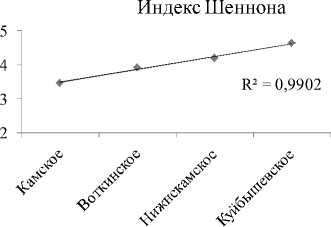
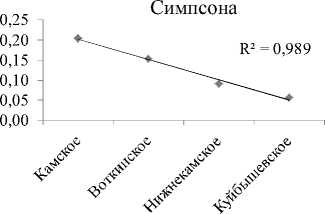
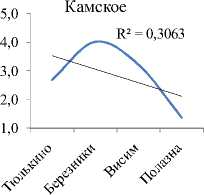
Видовое разнообразие. Интегральные индексы разнообразия Шеннона (Hn) и выровненности Пие-лу (E) с высокой достоверностью увеличиваются сверху вниз по каскаду (рис. 2А, табл. 3). В границах же отдельно взятого водохранилища все эти показатели (по каждой станции) показывают тенденцию снижения к плотине каждой ГЭС каскада (рис. 2В). На фоне этого, интегральный индекс доминирования (рис. 2А), напротив, по каскаду уменьшается сверху вниз. Косвенно это подтверждает то, что в Камском водохранилище, где индекс максимален, на долю 2-х первых доминантов приходится 56% от общей численности, в то время, как в Куйбышевском водохранилище – всего 22%. В целом же, изменения видового разнообразия сообщества инфузорий от станции к станции на Камском каскаде аналогичны таковым по каскаду Куйбышевское – Саратовское – Волгоградское водохранилища [4], носят волнообразный характер, и линейный тренд достоверно не отражает направления этих изменений по удельным характеристикам (рис. 3).
А
Индекс доминирования
1,00
0,80
0,60
0,40
0,20
0,00
Индекс выровненности Пиелу

R² = 0,916
В
Индекс Шеннона
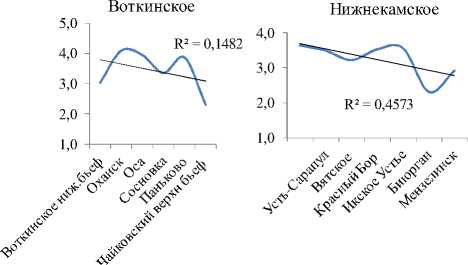
Рис. 2 . Тренды изменения интегральных (А) и удельных (В) показателей видового разнообразия сообществ инфузорий вдоль Камского каскада (А) и внутри отдельных водохранилищ (В)
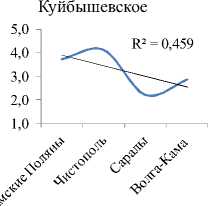
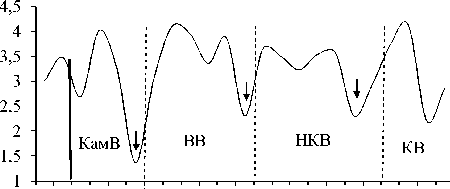
Изменение вдоль Камского каскада основных характеристик количественного развития сообществ инфузорий в планктоне. Максимальные значения численности, биомассы и продукции были зарегистрированы в водохранилище с наибольшей трофностью – Воткинском (табл. 3). В отличие от видового разнообразия, для этих показателей в целом характерна тенденция (тренд с незначительной достоверностью аппроксимации) снижения их средних значений (табл. 3) сверху вниз по каскаду водохранилищ, возрастания внутри каждого водохранилища по мере приближения к каждой ниже- лежащей плотине, а затем снижения в верхнем и нижнем бъефах каждой ГЭС (рис. 3).
Изменения численности и биомассы инфузорий в планктоне водохранилищ Камского каскада, как правило, несколько смещены во времени. В целом, схема динамики пространственного распределения инфузорий (рис. 3) аналогична для всех водохранилищ каскада и, по-видимому, имеет цикличный характер. В верхних бъефах плотин каждого вышележащего водохранилища (а также районе перехода от незарегулированного участка р. Камы непосредственно в Камское водохранилище) сообщест- ва инфузорий характеризуются повышением численности и биомассы, но с понижением видового разнообразия. Затем, в нижних бьефах плотин, где более высокая скорость течения и турбулентность, происходит резкое падение всех количественных показателей, но из-за перестройки сообщества начинает восстанавливаться видовое разнообразие.
Этот процесс продолжается и в средней части водохранилищ, где скорость течения снижается. Далее, по мере приближения к приплотинному участку (верхнему бъефу) каждой нижележащей ГЭС, у сообщества инфузорий вновь начинают повышаться показатели численности и биомассы, но опять с понижением видового разнообразия.
Таблица 3 . Средние по водохранилищам (± доверительный интервал) параметры развития сообщества инфузорий планктона (июль 2009 г.)
Индекс Шеннона
Численность, тыс. экз. /м3
Биомасса, мг / м3 ,
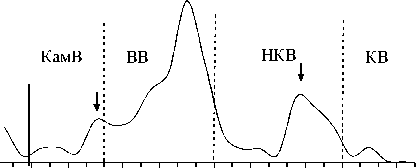
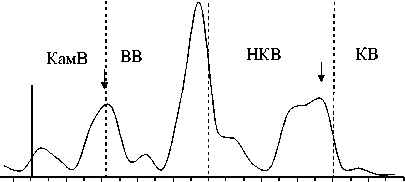
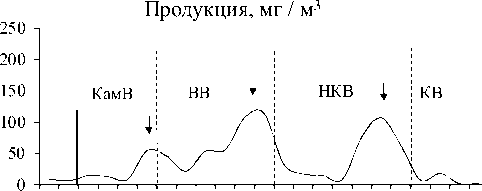
Рис. 3. Изменение основных показателей развития сообщества инфузорий по каскаду водохранилищ (пунктирная линия – граница между водохранилищами, плотина; сплошная линия отграничивает незарегулированную часть Камского водохранилища)
Сокращения: см. табл. 2
Чаще всего в условиях неустойчивого гидрологического режима (специфика водохранилищ) преимущество получает какая-либо экологически специализированная группа инфузорий, достигающая значительных численностей и биомасс (при этом степень доминирования – индекс Симпсона – увеличивается). Так, на станциях вблизи с плотинами, на долю первого доминанта приходится до 78% от общей численности, тогда как в середине водохранилища структура сообществ более выровнена и на долю первого доминанта приходится всего 17-39%.
Структурные перестройки сообществ инфузорий планктона по каскаду и внутри отдельных водохранилищ .
Каких-либо направленных изменений экотопи-ческой структуры сообщества инфузорий по каскаду водохранилищ в целом и внутри них нами не выявлено. Обычно такие изменения очень локальны и подчинены, скорее всего, специфике местных (гидрологических, гидрохимических, биотических и пр.) условий. Так, основу сообщества на протяжении всего каскада составляли эупланктонные виды (в среднем 89-93% от общей численности во всех водохранилищах). Отмечено увеличение роли эпипланктонных видов в Камском водохранилище (п. Висим) и Воткинском (п. Сосновка): 39% и 7% от общей численности. Увеличение вклада бентос- ных и перифитонных видов в сообществе инфузорий планктона выявлено лишь в Икском Устье (28%) и Камских Полянах (11%), т. е. точках, соответственно, на Нижнекамском и Куйбышевском водохранилищах. Но эти явления тоже локальны и их причины нами пока не установлены.
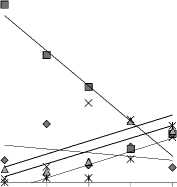
В трофической структуре прослеживаются направленные изменения сверху вниз по каскаду роли отдельных групп: наметилась тенденция (с различной достоверностью) снижения вклада в общую численность и биомассу бактериодетритофагов и альгофагов и увеличения неселективных всеядов, хищников и специфической экологической группы инфузорий-миксотрофов (табл. 4, рис. 4). Значительная роль бактеридетритофагов в верховьях Камского водохранилища и его незарегулирован-ной части обусловлена, вероятно, поступлением бактерий с болотными водами. В отличие от остальных трофических групп, для неселективных всеядов в пределах каждого водохранилища характерна повторяемость изменений: после каждой плотины их вклад в общую численность снижается и вновь увеличивается к следующей, но уже в большей степени, что и обусловливает общее увеличение роли данной группы сверху вниз по каскаду. Для других групп это наблюдается не всегда и не так отчетливо.
Таблица 4 . Изменение структурных показателей сообщества инфузорий по Камскому каскаду водохранилищ (июль 2009 г.)
|
Водоем |
Экотопическая структура |
Трофическая структура |
|||||||
|
Пл |
Эпл |
Пр |
Б |
А |
Б-Д |
М |
Н |
Х |
|
|
вклад по численности, % |
|||||||||
|
Незарегулированный участок Камы |
89,8 |
0,0 |
2,3 |
7,9 |
21,8 |
48,1 |
21,3 |
8,8 |
0,0 |
|
Камское водохранилище |
93,2 |
3,3 |
2,4 |
1,1 |
25,9 |
60,7 |
3,0 |
8,3 |
2,1 |
|
Воткинское водохранилище |
89,4 |
1,6 |
7,2 |
1,8 |
7,6 |
23,3 |
10,4 |
55,9 |
2,8 |
|
Нижнекамское водохранилище |
91,6 |
0,0 |
1,5 |
6,9 |
11,2 |
15,8 |
9,9 |
56,5 |
6,6 |
|
Куйбышевское водохранилище |
89,3 |
0,3 |
4,8 |
5,5 |
14,8 |
27,8 |
15,1 |
32,6 |
9,6 |
|
вклад по биомассе,% |
|||||||||
|
Незарегулированный участок Камы |
98,3 |
0,0 |
0,6 |
1,1 |
10,3 |
81,8 |
6,2 |
1,8 |
0,0 |
|
Камское водохранилище |
94,0 |
4,2 |
1,4 |
0,4 |
26,9 |
58,7 |
5,0 |
7,3 |
2,0 |
|
Воткинское водохранилище |
96,5 |
0,8 |
1,9 |
0,8 |
8,0 |
43,9 |
9,5 |
36,6 |
2,1 |
|
Нижнекамское водохранилище |
94,2 |
0,0 |
1,4 |
4,4 |
16,5 |
15,5 |
28,2 |
29,0 |
10,8 |
|
Куйбышевское водохранилище |
96,7 |
0,1 |
2,2 |
1,0 |
6,9 |
22,4 |
23,8 |
20,8 |
26,1 |
Примечание : Пл – планктонные, Эпл – эпипланктонные, Б-Д – бактериодетритофаги, М – миксотрофы (симбиотрофы), Н – неселективные всеяды, Х – хищники
Анализ полученных нами данных не выявил связи развития сообщества инфузорий с большинством абиотических факторов. Достоверные положительные корреляции численности и биомассы имеются лишь с температурой и содержанием кислорода. Полагаем, что характер изменения параметров сообщества инфузорий в большей степени определяется (как и на Волжском каскаде) совместным воздействием не только гидрохимических параметров среды, которые часто носят локальный характер в отдельных точках мелководной зоны, но и спецификой гидрологического режима каждого из водохранилищ каскада (местоположение в каскаде, режим регулирования стока, внутренняя динамика вод и уровенный режимы, прямые и обратные волны от ГЭС и т.д.).
Таким образом, трансформация сообщества свободноживущих инфузорий, четко проявляющаяся в изменении интегральных показателей видового разнообразия, затрагивает как видовую структуру (пусть не всегда явно), так и общие и структурные количественные показатели сообщества инфузорий планктона. Трансформация сообществ инфузорий внутри каждого водохранилища каскада происходит циклично, что, вероятно, обусловлено неустойчивостью системы, связанной с пульсационной работой ГЭС. При этом вышеуказанное пространственное распределение инфузорий по всему Камскому каскаду практически повторяет описанное для каскада водохранилищ Средней и Нижней Волги, где оно «всегда имеет характер непрерывной «волны», переходящей из одного водохранилища в другое с мощной деформацией в верхних и нижних бьефах каждой ГЭС» [4, 5].
90,0
80,0
70,0
60,0
50,0
40,0
30,0
20,0
10,0
%
0,0
□ Бактериодетритофаги R² = 0,7429
л Миксотрофы R² = 0,8977
X Неселективные всеядыR² = 0,7911
ж Хищники
ф Альгофаги R² = 0,1085
R² = 0,4226
Рис. 4. Тренды (с коэффициентами достоверности аппроксимации) изменения вклада в общую биомассу (%) основных структурных (трофи-ческих) групп инфузорий по каскаду
Обозначения: 1 – незарегулированный участок Камы, 2 – Камское, 3 –Нижнекамское, 4 – Воткинское, 5 – Куйбышевское водохранилища
В целом, наложение волнообразного изменения параметров сообщества инфузорий планктона вдоль каскада (рис. 3) и определенной направленности изменения интегральных показателей видового разнообразия (рис. 2) и количественных характеристик показывает «ступенчатое» (дискретное) их изменение сверху вниз. Вероятно, концепция речного континуума, выдвинутая для условий проточной незарегулированной реки [12], для каскадной системы водохранилищ не работает, и ей
можно противопоставить дискретность смены вариантов сообществ.
Список литературы Инфузории (Ciliophora) планктона мелководной зоны водохранилищ Камского каскада
- Винберг Г.Г. Температурный коэффициент Вант-Гоффа и уравнение Аррениуса в биологии//Журн. общ. биол. 1983. Т. 44, вып. 1. С. 31-42.
- Государственный водный кадастр. Разд. 1. Поверхностные воды. Сер. 3. Многолетние данные. Многолетние данные о режиме и ресурсах поверхностных вод суши. Ч. 2. Озера и водохранилища. Т. 1. РСФСР. Вып. 24. Бассейны рек Волги (среднее и нижнее течение) и Урала. Л., 1985. 517 с.
- Жариков В.В. Кадастр свободноживущих инфузорий водохранилищ Волги. Тольятти, 1996. 76 с.
- Жариков В.В. Свободноживущие инфузории Волги: состав, динамика и пространственно-временное распределение в условиях полного гидротехнического зарегулирования реки: Автореф. дис. … д-ра биол. наук. СПб, 1999. 45с.
- Жариков В.В. Специфика водохранилищ Волги как среды обитания гидробионтов (На примере свободноживущих инфузорий)//Теоретические проблемы экологии и эволюции (Третьи Любищевские чтения). Тольятти: ИЭВБ РАН, 2000. С.64-72.
- Ковальчук А.А. Некоторые вопросы экофизиологии свободноживущих инфузорий//Гидробиол. журн. 2001. Т. 38, № 3. С. 81-88.
- Куйбышевское водохранилище (научно-информационный справочник)/Отв. Ред. Г.С. Розенберг, Л.А. Выхристюк. Тольятти: ИЭВБ РАН, 2008. 123 с.
- Мыльникова З.М. Планктонные инфузории камских водохранилищ//Биология внутренних вод. Информ. бюл. Л., Наука. 1990. № 86. С. 38-41.
- Уманская М.В., Краснова Е.С., Горбунов М.Ю. Химический состав воды и трофический статус прибрежных участков водохранилища Камского каскада в 2009 г.//Самарская Лука: проблемы региональной и глобальной экологии: Бюлл. 2011. Т. 20, № 3. С. 39-49.
- Хлебович Т.В. Значение планктонных инфузорий в биологической трансформации вещества и энергии: Автореф. дисс.. канд. биол. наук. Л., 1986. 19 с.
- Эдельштейн К.К. Водохранилища России: экологические проблемы, пути их решения. М.: ГЕОС, 1998. 277 с.
- Vannote R.L., Minshall G.W., Cummins K.W., Sedell I.R., Cushing C. E. The river continuum concept//Can. J. Fich. Aquat. Sci., 1980, V. 37, N 1. P. 130-137.
