Инфузории озера Раифское (Волжско-Камский биосферный заповедник)
")
Автор: Быкова С.В., Жариков В.В.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Итоги научных исследований
Статья в выпуске: 3 т.18, 2009 года.
Бесплатный доступ
В работе приведены данные по изучению в заповедном оз. Раифском важной группы планктонного сообщества - инфузорий. Выявлено 94 вида. Показано, что видовой состав богаче и видовое разнообразие выше в сообществе инфузорий, развивающихся в зарослях макрофитов (58% всех зарегистрированных в водоеме видов инфузорий встречено только здесь). Однако численность и биомасса инфузорий в зарослях макрофитов ниже, чем в пелагиали. Проникновение видов изначально разной биотопической принадлежности в сообщество инфузорий в зарослях высшей водной растительности делает его структуру сложнее и разнообразней в таксономическом, видовом, трофическом и экологическом плане. Установлено, что сильнее отличаются по составу фауны от сообщества инфузорий пелагиали сообщества инфузорий, развивающихся в погруженных водных растениях. Таким образом, в изучении биоразнообразия гидробионтов водоема в целом важно уделять внимание исследованию сообществ инфузорий как пелагической, так и зарослевой частей водоема.
Инфузории, биоразнообразие, планктон, пелагиаль, перифитон, сообщества высших водных растений
Короткий адрес: https://sciup.org/148313226
IDR: 148313226 | УДК: 574.583:593.17
Текст научной статьи Инфузории озера Раифское (Волжско-Камский биосферный заповедник)
Материал обобщен в таксономической системе Смолла и Линна (Small and Lynn, 2000). Индексы сапробности ряда видов инфузорий-индикаторов скорректированы по: Foissner (1988).
РАЙОН ИССЛЕДОВАНИЙ
Оз. Раифское является центральным звеном единой гидросистемы, объединяющей поверхностные воды (р. Сумка и ее приток р. Сер-Булак и озера в их долинах) Раифского участка ВКЗ и его охранной зоны (рис. 1). Это достаточно крупный водоем: по данным 1995 г. его максимальная глубина - 19,6 м, площадь зеркала озера – 31,9 га, но эти показатели постепенно уменьшаются из-за наносов взвешенных частиц р. Сумкой и ее притоком (Унковская и др., 2002). Глубина озера в месте отбора проб – 17,5 м, площадь зарастания водоема высшей водной растительностью составляет примерно 7-10% (устное сообщение Е. Унковской)
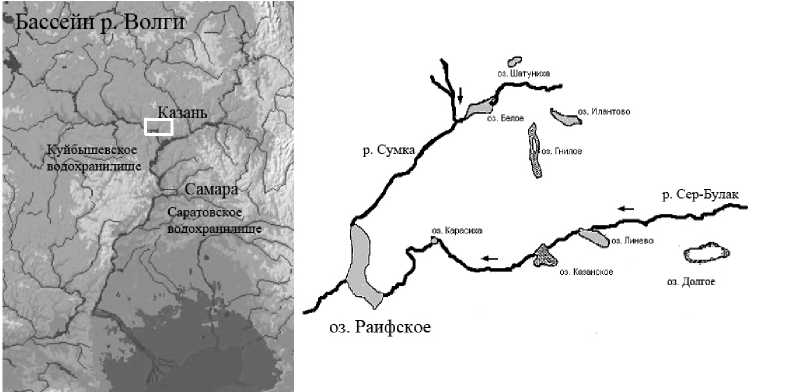
Рис. 1. Район исследований
Прозрачность воды варьирует от 0,3 м до 0,8 м. Температурный и газовый режимы озера соответствуют режиму глубоких стратифицированных водоемов. Летом температурный скачок отмечается между 3-4 м и 6 м, оксиклин – между 1,5 и 4 м. Градиент температуры в зоне скачка достигает 5,8-7,4° С на один метр, градиент кислорода в зоне оксиклина – 10,2 мг/м3 на 1 м (Уманская и др., 2007). Вода в озере кальций-гидрокарбонатного типа, средней минерализации, по общей жесткости относится к категории мягкой и умеренно-жесткой (Унковская и др., 2002). pH среды колеблется от 6,9 в придонных слоях до 8,2-8,7 в поверхностных слоях воды. Трофический статус по биомассе фитопланктона оценивается как меняющийся от олиго- до высокоэвтрофного (Палагушкина и др., 2002).
Исследования простейших на водоемах ВКЗ, в том числе и инфузорий, ранее никогда не проводились, в то время как первые упоминания о зоопланктоне относятся к 1921 г. С 1983 года изучение видового состава и количественных характеристик сообществ фитопланктона, зоопланктона и зообентоса оз. Раифское является частью экологического мониторинга (программа «Летопись природы») водоемов заповедника (Деревенская и др., 2002; Палагушкина и др., 2002; Унковская и др., 2002). В дополнение к ней ИЭВБ РАН в 2006-2007 гг. были осуществлены также первые исследования фитопланктона (Тарасова, 2008) и зоопланктона (Мухортова, 2008) зарослевой части водоема.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Таксономическое и видовое разнообразие. В результате наших исследований в оз. Раифское было зарегистрировано около 94 видов инфузорий (табл. 1), из которых 2 вида пока не удалось идентифицировать даже до рода. Выявленные инфузории относятся к 2 подтипам, 8 классам, 11 подклассам, 22 отрядам, 42 семействам, 61 роду.
В планктоне пелагической части озера зарегистрировано 43 вида, в планктоне зарослей макрофитов – 49 видов, в перифитоне (на стеклах обрастания и на естественных субстратах) – 28 видов (табл. 1). Удельное число видов (количество видов в отдельных пробах) на разных глубинах в пелагиали в 2006 г. варьирует от 2 до 17 (в среднем, 11 видов), в планктоне зарослей различных макрофитов – от 15 до 27 видов (в среднем, 20 видов), т. е. α-разнообразие сообщества инфузорий в зарослях макрофитов в 1,8 раза выше пелагического.
Это соотношение сохраняется и для β-разнообразия пелагического (27 видов со всех горизонтов в толще воды) и «зарослевого» (49 видов во всех исследованных в озере макрофитах) сообществ. Индекс видового разнообразия Шеннона (табл. 2) также выше в сообществе зарослевой части инфузорий (2,78-4,03). Лишь для сообщества инфузорий в зарослях кубышки (2,51), данный показатель близок к показателю пелагического планктона (2,52). Из 65 видов инфузорий, зарегистрированных в планктоне в 2006 г., 38 видов (58%) встречены только в зарослях макрофитов, 17 (26%) – только в пелагиали и 10 видов (15%) были общими для этих биотопов.
В пелагической части озера выявлено 27 эупланктонных видов инфузорий планктона, 5 перифитонных (эпифитных), 4 бентосных вида. Эпифитные виды, встречающиеся в пелагическом планктоне, представлены, в основном, Suctoria и Peritricha, сидящими на планктонных водорослях ( Microcystis, Ana-baena, Scenedesmus ) и ракообразных ( Daphnia ). Бентосные виды в планктоне оз. Раифское – сапропельные виды р. Brachonella, Metopus , поднимающиеся в вышележащие (вплоть до глубины 4 м) анаэробные слои толщи.
В сообществе макрофитов формируется комплекс подвижных форм инфузорий, промежуточный между сообществом планктона, перифитона и бентоса. Среди зарослей макрофитов зарегистрировано 28 эупланктонных видов инфузорий, 20 перифитонных, 1 бентосный вид. Большое число перифитон-ных видов в планктоне зарослей макрофитов обусловлено наличием твердого субстрата (листья, стебли растений).
Таблица 1
Видовой состав инфузорий оз. Раифское
|
№№ |
Экологические группы |
Планктон |
Перифитон |
||||||
|
Биотоп Видовой состав |
Заросли макрофитов |
Пела-гиаль |
Заросли + пела-гиаль |
||||||
|
н u я я о я к о го Я О & я н о о ч Рч |
=я S я н о я я о я о н н о d & |
а ю и |
и я я н о я я о о & |
40 о о Г1 |
о о Г1 |
40 О о Г1 |
о о Г1 |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Тип CILIOPHORA Doflein, 1901 |
|||||||||
|
П/тип POSTCILIODESMATOPHORA |
|||||||||
|
Кл. KARYORELICTEA Corliss, 1974 |
|||||||||
|
1. |
Loxodes striatus (Engelmann, 1862) |
+ |
|||||||
|
Кл. HETEROTRICHEA Stein, 1859 |
|||||||||
|
2. |
Stentor roeseli Ehrb., 1835 |
+ |
|||||||
|
3. |
Folliculina boltoni Kent, 1881 |
+ |
+ |
+ |
+ |
||||
|
П/тип INTRAMACRONUCLEATA Lynn, 1996 |
|||||||||
|
Кл. SPIROTRICHEA Butschli, 1889 |
|||||||||
|
4. |
Aspidisca cicada (O.F.Muller, 1786) |
+ |
+ |
||||||
|
5. |
A. lynceus (O.F.Muller, 1786) |
+ |
|||||||
|
6. |
Chaetospira remex (Hudson, 1875) |
+ |
+ |
||||||
|
7. |
Codonella cratera (Leidy, 1887) |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
8. |
Euplotes sp. |
+ |
|||||||
|
9. |
Halteria grandinella (O.F. Muller, 1773) |
+ |
+ |
+ |
|||||
|
10. |
Pelagohalteria viridis (Fromentel, 1876) Foissner, Skogstad & Pratt, 1988 |
+ |
+ |
||||||
|
11. |
Limnostrombidium viride (Stein, 1867) Krainer, 1995 |
+ |
+ |
||||||
Продолжение табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
||
|
12. |
Limnostrombidium pelagicum (Kahl, 1932) Krainer, 1995 |
+ |
+ |
||||||||
|
13. |
Oxytricha sp. |
+ |
+ |
+ |
|||||||
|
14. |
Pelagostrombidium fallax (Zacharias, 1895) Krainer, 1991 |
+ |
+ |
||||||||
|
15. |
Rimostrombidium humile (Penard, 1922) Petz & Foissner, 1992 |
+ |
+ |
||||||||
|
16. |
Rimostrombidium lacustris (Foissner, Skogstad & Pratt, 1988) Petz & Foissner, 1992 |
+ |
+ |
||||||||
|
17. |
Stichotricha aculeata Wrzesniowski., 1870 |
+ |
+ |
||||||||
|
18. |
Strobilidium caudatum (Fromentel, 1876) |
+ |
+ |
+ |
|||||||
|
19. |
Stylonychia mytilus (Ehrb., 1838) complex |
+ |
|||||||||
|
20. |
Tintinnidium fluviatile (Stein, 1863) |
+ |
+ |
+ |
+ |
||||||
|
21. |
Tintinnopsis cylindrata Kof. & Cam., 1892 |
+ |
+ |
||||||||
|
22. |
Uroleptus piscis (Mull.) Ehrb., 1831 |
+ |
|||||||||
|
23. |
Carchesium polypinum Linnaeus, 1758 |
+ |
|||||||||
|
24. |
Cinetochilum margaritacium Perty, 1852 |
+ |
+ |
||||||||
|
Sedis mutabilis в п/типе Intramacronucleata |
|||||||||||
|
25. |
Brachonella spiralis (Smith, 1897) |
+ |
+ |
||||||||
|
26. |
Brachonella sp. |
+ |
+ |
||||||||
|
27. |
Metopus es (O.F. Muller, 1776) |
+ |
|||||||||
|
28. |
Metopus sp. 1 |
+ |
|||||||||
|
29. |
Metopus sp. 2 |
+ |
|||||||||
|
30. |
Metopus sp. 3 |
+ |
|||||||||
|
Кл. LITOSTOMATEA Small & Lynn, 1981 |
|||||||||||
|
31. |
Acaryophrya sphaerica Foissner, 1983 |
+ |
+ |
||||||||
|
32. |
Amphileptus pleurosigma (Stokes, 1884) |
+ |
|||||||||
|
33. |
Askenasia acrostomia Krain.& Foissn., 1990 |
+ |
+ |
+ |
|||||||
|
34. |
A. chlorelligera Krainer & Foissner, 1990 |
+ |
+ |
||||||||
|
35. |
Chaenea teres (Dujardin, 1841) |
+ |
+ |
||||||||
|
36. |
Enchelys pupa (O.F.Muller, 1786) |
+ |
|||||||||
|
37. |
E. simplex Kahl, 1926 |
+ |
+ |
+ |
+ |
||||||
|
38. |
Lacrymaria olor (O.F. Muller, 1786) |
+ |
|||||||||
|
39. |
Lacrymaria sp. |
+ |
|||||||||
|
40. |
Lagynophrya acuminata Kahl, 1935 |
+ |
+ |
||||||||
|
41. |
Litonotus crystallinus ( Vuxanovici, 1960) |
+ |
|||||||||
|
42. |
L. cygnus (O.F.Muller, 1773) |
+ |
+ |
||||||||
|
43. |
Spathidium viride Kahl, 1926 |
+ |
+ |
||||||||
|
44. |
Spathidium spathula (O.F.Muller, 1773) |
+ |
|||||||||
|
45. |
Spathidium sp. 1 |
+ |
+ |
||||||||
|
46. |
Spathidium sp. 2 |
+ |
|||||||||
|
47. |
Trachelius ovum (Ehrenberg, 1831) |
+ |
|||||||||
|
48. |
Trachelophyllum vestitum (Stokes, 1884) |
+ |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Кл. PHYLLOPHARYNGEA de Puytorac et al., 1974 |
|||||||||
|
49. |
Chilodonella uncinat a (Ehrenberg, 1838) |
+ |
|||||||
|
50. |
Chilodonella sp. |
+ |
+ |
+ |
+ |
||||
|
51. |
Gastronauta membranaceus Engelmann, 1875 |
+ |
+ |
||||||
|
52. |
Metacineta cuspidata (Kellicott, 1885) |
+ |
|||||||
|
53. |
M. longipes var.septimfida Rieder, 1985 |
+ |
|||||||
|
54. |
M. mystacina v. brevipes Sand, 1900 |
+ |
|||||||
|
55. |
Trithigmostoma cucullulus (O.F.Mull.,1786) |
+ |
|||||||
|
Кл. NASSOPHOREA Small & Lynn, 1981 |
|||||||||
|
56. |
Furgasonia trichocystis (Stokes, 1894) |
+ |
|||||||
|
57. |
Nassula sp. |
+ |
+ |
||||||
|
Кл. PROSTOMATEA Schewiakoff, 1896 |
|||||||||
|
58. |
Coleps hirtus viridis Ehrenberg, 1831 |
+ |
+ |
+ |
+ |
+ |
|||
|
59. |
Holophrya sp. |
+ |
|||||||
|
60. |
Lagynus elegans (Engelmann, 1862) |
+ |
|||||||
|
61. |
Urotricha gen. sp. |
+ |
+ |
+ |
+ |
||||
|
Кл. OLIGOHYMENOPHOREA de Puytorac et al., 1974 |
|||||||||
|
62. |
Lembadion bullinum Perty, 1852 |
||||||||
|
63. |
Ophryoglena sp. 1 |
+ |
+ |
||||||
|
64. |
Ophryoglena sp. 2 |
+ |
|||||||
|
65. |
Paramecium bursaria (Ehrb.,1831) |
+ |
|||||||
|
П/кл. Peritrichia Stein, 1859 |
|||||||||
|
66. |
Astylozoon sp. |
+ |
|||||||
|
67. |
Carchesium polypinum Linnaeus, 1758 |
+ |
|||||||
|
68. |
Cinetochilum margaritacium Perty, 1852 |
+ |
+ |
||||||
|
69. |
Cothurnia annulata Stokes, 1885 |
+ |
+ |
+ |
+ |
||||
|
70 |
Ctedoctema acanthocrypta Stokes, 1884 |
+ |
+ |
+ |
|||||
|
71. |
Cyclidium citrullus Cohn, 1865 |
+ |
+ |
||||||
|
72. |
Cyclidium glaucoma (O.F.Mull., 1773) |
+ |
+ |
+ |
+ |
||||
|
73. |
Epistylis procumbens Zacharias, 1897 |
+ |
+ |
+ |
+ |
||||
|
74. |
Epistylis sp. |
+ |
+ |
||||||
|
75. |
Haplocaulus anabaena (Stiller, 1940) |
+ |
|||||||
|
76. |
Opercularia nutans (Ehrb., 1831) |
+ |
|||||||
|
77. |
Ophrydium versatile Muller, 1786 |
+ |
+ |
||||||
|
78. |
Platycola decumbens (Ehrb., 1830) |
+ |
+ |
+ |
|||||
|
79. |
Vorticella spp. (<30 мкм) |
+ |
|||||||
|
80. |
Vorticella natans (F.Fremiet, 1924) |
+ |
|||||||
|
81. |
V. campanula Ehrb., 1831 |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
82. |
V. convallaria (Linnaeus, 1758) |
+ |
+ |
+ |
|||||
|
83. |
V. octava complex Stokes, 1885 |
+ |
|||||||
|
84. |
Vorticella sp. 1 (на Anabaena) |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
85. |
Vorticella sp. 2 (на Daphnia) |
+ |
|||||||
|
86. |
Vorticella sp.3 (на Scenedesmus) |
+ |
Окончание табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
87. |
Vorticella sp. 4 (на Microcystis) |
+ |
|||||||
|
88. |
Vaginicola crystallina Entz., 1884 |
+ |
+ |
||||||
|
89. |
V. tincta Ehrenberg, 1830 |
+ |
|||||||
|
90. |
Zoothamnium arbuscula (Ehrb., 1831) |
+ |
+ |
||||||
|
91. |
Z. hentscheli Kahl, 1935 |
+ |
|||||||
|
92. |
Zoothamnium sp. |
+ |
|||||||
|
93. |
Неидентифицированные до рода Ciliatа 1 |
+ |
+ |
||||||
|
94. |
Неидентифицированные до рода Ciliatа 2 |
+ |
+ |
||||||
|
Итого: |
15 |
28 |
15 |
23 |
27 |
33 |
21 |
19 |
|
|
49 |
43 |
28 |
|||||||
Сходство фауны фитофильных инфузорий с фауной пелагиали в целом составляет 32% (только поверхностного слоя пелагиали – 43%), с фауной инфузорий перифитона (со стекол обрастаний) – 20%, Наибольшим сходством (53,8%) с фауной пелагической части и наименьшим с фауной перифитона (11,4%) характеризуется сообщество инфузорий в зарослях кубышки – высшего водного растения с плавающими листьями. Возможно, это обусловлено тем, что заросли кубышки хорошо промываются и имеют меньшую, по сравнению с погруженными растениями, площадь зарастания. Погруженные растения с мелкими листьями (рдесты, роголистник), напротив, предоставляют бόльшую площадь для обрастания организмам, что, соответственно, приводит к развитию бόльшего количества видов инфузорий-обрастателей в образуемых ими сообществах. Кроме того, плотные заросли, часто образуемые погруженными растениями, вероятно, способствуют формированию локальных сообществ инфузорий. Вследствие этого коэффициент сходства фауны инфузорий зарослей рдестов и роголистника с пелагической фауной ниже (15%-39%), чем в зарослях кубышки, а с фауной перифитона – выше (20%-22%). Кластерный анализ сходства фауны показал, что сообщества инфузорий планктона и перифитона явно обособлены друг от друга (рис. 2). В составе планктона в отдельные группы выделяются сообщества, развивающиеся в пе-лагиали и зарослях кубышки, с одной стороны, и погруженных макрофитах (рдесты и роголистник), с другой.
Доминанты пелагиали . В поверхностном слое по численности преобладают Vorticella sp. (на Anabaena), Askenasia sp. с зоохлореллами. У верхней границы оксиклина, максимумы численности образуют Vorticella sp, Enchelys sp., Codonella cratera , у нижней – миксотроф Coleps hirtus viridis . В анаэробной зоне доминируют по численности мелкие инфузории-бактериофаги. По биомассе в поверхностных слоях доминирует Epistylis procumbens , Nassula sp., Rimostrombidium lacustris , Ophryoglena sp. 1, а в анаэробной зоне – Metopus sp. 2, Codonella cratera .
Доминанты инфузории в зарослях макрофитов. По численности преобладают Vorticella sp. 1 (на Anabaena), V. campanula Ehrb., 1831, Codonella cratera, Cyclidium citrullus, а по биомассе – Folliculina boltoni, Litonotus cygnus, Strobilidium caudatum, Ophryoglena sp. 1 и sp. 2. Кроме того, при исследовании сообщества инфузорий зарослей рдеста пронзеннолистного и роголистника, в планктонных пробах нами обнаружены взрослые особи пресновод- ной Folliculina boltoni, ведущей обычно исключительно прикрепленный образ жизни. На домиках фолликулин сохранялся хорошо различимый даже без предварительного окрашивания прикрепительный слой («клей», collectoderm). Появление фолликулин в планктоне отмечалось в период массового развития F. boltoni на стеклах обрастания, где его численность была максимальной за все время исследований – 343 тыс. экз./м2. В зарослях других видов макрофитов находок F. boltoni не отмечалось. Факт присутствия фолликулин в планктонных пробах зарослевого планктона важен тем, что косвенно указывает на возможный характерный природный субстрат (рдест, роголистник) обитания F. boltoni в бассейне Волги.
п л а н к т о н
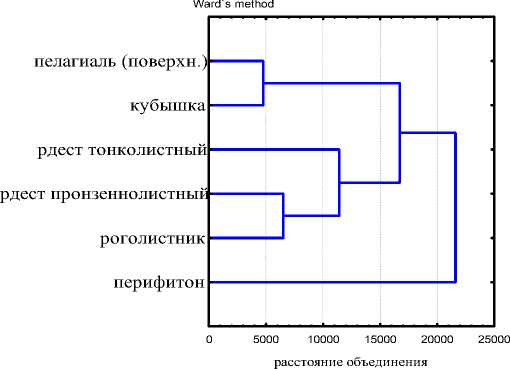
Рис. 2. Дендрограмма сходства сообществ инфузорий
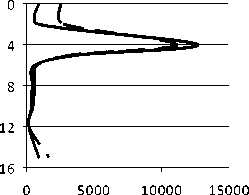
Количественное развитие . Сравнительный анализ количественных параметров развития сообщества инфузорий пелагической части водоема в 2006 и 2007 гг.. показал, одновременно с увеличением количества видов, наблюдается снижение общей численности в 2,5 раза, биомассы – в 2,4 в 2007 г. (рис. 3А).
АВ
2006 2007 2006 г . 2007 г.
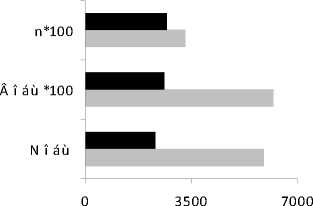
Рис. 3. Показатели количественного развития пелагического сообщества инфузорий в 2006-2007 гг.: А - общее количество видов (n), общая численность (N, тыс. экз./м3), биомасса (В, мг/м3); В - вертикальное распределение (N, B).
N общ В общ.*100
Максимумы численности (18414 тыс. экз./м3) и биомассы (153,4 мг/м3) регистрировались в 2006 г. на 10 м, в 2007 г. – на 4 м (соответственно, 12669 тыс. экз./м3 и 111,8 мг/м3) (рис. 3В).
В целом, за небольшим исключением, численность и биомасса инфузорий в зарослях макрофитов (N = 347 - 406 тыс. экз/м3 и В = 8,2 – 14,5 мг/м3) ниже, чем в пелагиали (табл. 2). Возможно, это связано с наличием пищевой конкуренции либо выеданием инфузорий массой развивающихся фитофиль-ных ракообразных, коловраток и мальков рыб.
Таблица 2
численность, тыс. экз./м3
пелагиаль макрофиты
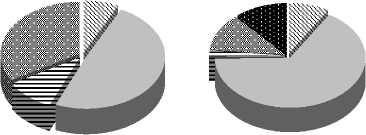
биомасса, мг/м3
пелагиаль макрофиты
Показатели развития инфузорий планктона в разных биотопах оз. Раифского в 2006 г.
|
биотоп |
Параметры |
||||||
|
n |
я |
N общ |
В общ |
P/B сут. |
W |
S |
|
Пелагиаль
|
0 м* |
11 |
2,51 |
603,9 |
14,7 |
2,25 |
0,024399 |
1,55 |
|
2 м |
12 |
1,98 |
3478,2 |
99,0 |
1,64 |
0,028454 |
1,53 |
|
4 м |
10 |
1,60 |
4286,7 |
80,2 |
1,03 |
0,018713 |
1,78 |
|
5 м |
12 |
2,05 |
2039,4 |
28,7 |
0,82 |
0,014089 |
1,66 |
|
6 м |
11 |
2,22 |
679,8 |
9,8 |
0,73 |
0,014403 |
2,44 |
|
8 м |
17 |
0,59 |
9655,8 |
79,5 |
0,70 |
0,008231 |
2,39 |
|
10 м |
10 |
0,15 |
18414,0 |
134,2 |
0,73 |
0,007290 |
1,98 |
|
12 м |
15 |
0,34 |
13404,6 |
108,7 |
0,65 |
0,008113 |
3,12 |
|
15 м |
2 |
0,06 |
435,6 |
3,0 |
0,58 |
0,007000 |
- |
|
Заросли макрофитов |
|||||||
|
Рдест пронзеннолист. |
16 |
2,78 |
346,5 |
14,56 |
1,64 |
0,042009 |
2,16 |
|
Рдест тонколистный |
27 |
4,03 |
1118,7 |
80,02 |
1,33 |
0,071253 |
2,43 |
|
Кубышка |
15 |
2,52 |
405,9 |
8,24 |
2,19 |
0,020305 |
2,18 |
|
Роголистник |
22 |
3,14 |
376,2 |
12,24 |
1,80 |
0,032350 |
1,90 |
Обозначения: n – число видов; н – индекс Шеннона, бит; N – общая численность, тыс. экз./м3; В – общая биомасса, мг/м3; W – средний индивидуальный вес особи, мг/м3; S – индекс сапробности среды; * - жирным шрифтом выделены сравниваемые показатели/ в альгофаги ■ бактериодетритофаги миксотрофы в неселективы и хищники
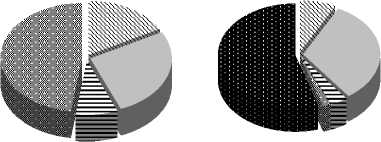
Рис. 4. Трофическая структура сообществ инфузорий в различных биотопах
Трофическая структура сообществ инфузорий пелагической и заросле-вой частей водоема различается. Особенностью сообществ инфузорий, развивающихся в зарослях макрофитов, является появление (в пелагической части их нет) хищников (до 11,5% от общей численности и 48% от общей биомассы), что указывает на усложнение трофической структуры данных сообществ.
На фоне этого снижается, по сравнению с сообществом пелагиали, вклад неселективных всеядов (с 33% до 12% по численности и с 48% до 1,2% по биомассе) и миксотрофов (с 10% до 2% (по N) и с 8% до 3% (по В)). Снижение доли от общих N и В последних обусловлено, возможно, худшими световыми условиями.
Размерная структура : в зарослях зарегистрировано больше крупных видов, о чем косвенно свидетельствуют больший, чем в сообществе инфузорий пелагиали средний индивидуальный вес и меньший Р/B-коэффициент (табл. 2).
Диапазон изменений индекса сапробности среды, определенный по численности инфузорий-индикаторов больше в пелагической части: от 1,55 (олиго-β-мезосапробная зона) на поверхности, до 3,12 (полисапробная зона) в придонных слоях (табл. 2). В макрофитах уровень органического загрязнения выше, чем в поверхностном горизонте пелагиали, о чем свидетельствуют индексы сапробности 2,16-2,43, соответствующие β-α-мезосапробной зоне.
ЗАКЛЮЧЕНИЕ
Таким образом, в оз. Раифское максимальное разнообразие инфузорий характерно для зарослей макрофитов, где обнаружено 73% всех выявленных видов, а 58% – только в зарослях макрофитов. Специфичность фауны отдельного водоема, вероятно, в значительной степени определяется именно фитофильной фауной, что необходимо учитывать в исследованиях. Наличие в толще воды среди высшей водной растительности видов инфузорий с явно разной биотопической принадлежностью делает структуру сообщества сложнее и разнообразней в таксономическом, трофическом и экологическом плане. Из этого следует важное обстоятельство: оценка биологического разнообразия инфузорий и других групп гидробионтов водоема лишь по видовому составу пелагической части водоема, без учета фауны его зарослевой части (с сильно варьирующей площадью развития) – некорректна и неправомерна.
Список литературы Инфузории озера Раифское (Волжско-Камский биосферный заповедник)
- Деревенская О.Ю., Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Структура сообществ зоопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны // Тр. Волжско-Камского государственного природного заповедника. 2002. Вып. 5. С. 52-70.
- Жариков В.В. Кадастр свободноживущих инфузорий водохранилищ Волги. Тольятти, 1996. 76 с.
- Мухортова О.В. Сообщества зоопланктона пелагиали и зарослей высших водных растений разнотипных водоемов Средней и Нижней Волги. Автореф. дис. … канд. биол. наук. Тольятти, 2008. 22 с.
- Палагушкина О.В., Бариева Ф.Ф., Унковская Е.Н. Видовой состав, биомасса и продуктивность фитопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны // Тр. Волжско-Камского государственного природного заповедника. 2002. Вып. 5. С.37-52.
- Тарасова Н.Г. Водоросли биоценозов высших водных растений озер Волжско-Камского заповедника // Природное наследие России в 21 веке. Материалы II международной научно-практической конференции. Башкирский государстенный анрарный университет, 25-27 сентября 2008 г. С. 380-384.
- Уманская М.В., Горбунов М.Ю., Унковская Е.Н. Бактериопланктон озер Раифы (Татарстан, Россия) // Изв. Самар. НЦ РАН, 2007. С. 987-995.
- Foissner W. Taxonomic and nomenclatural revision of Sladacek's list of ciliates (Protozoa: Ciliophora) as indicators of water quality. Hydrobiologia, 166, 1988. P. 1-64.
- Small, Lynn Phylum Ciliophora Doflein,1901 // An Illustrated guide to the protozoa. Second edition / Lee J.J., Leedale G.F., Bradbury Ph.(eds.). Lawrence, Kansas: Allen Press. 2000. P. 371-675.
