Interaction of bulk and nano titanium dioxide on antioxidant activity of Lallemantia iberica under drought conditions

Author: Shoarian Sattari Niloufar, Jamei Rashid, Pasban Eslam Bahman, Salehi Lisar Seyed Yahya
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 2 т.16, 2020.
Free access
This research was carried out to study the effect of Tio2 and nano Tio2 on some chemical compounds in a medicinal plant Lallemantia iberica under water deficit conditions. The experiment was conducted as a factorial arrangement in randomized complete block design with three replications. The first factor was the application of external materials as spraying with 3 levels of control, titanium dioxide and nano titanium dioxide, and the second factor was 100% (non stress), 75% (moderate stress) and 35% (severe stress) irrigation content. Stress was applied sixty days after planting to harvesting stage. The catalase and superoxide dismutase activities, total phenol, antioxidant activity with DPPH, total flavonoid and HPLC of phenolic compounds were evaluated in the samples. The results showed a moderate stress and the use of nTiO2 caused a significant increase in phenolic content, total flavonoid and antioxidant activity; severe drought stress significantly reducing these compounds...
Antioxidant activity, l. iberica, ntio2, phenolic
Short address: https://sciup.org/143171164
IDR: 143171164
Text of the scientific article Interaction of bulk and nano titanium dioxide on antioxidant activity of Lallemantia iberica under drought conditions
Drought is one of the most important stress factors that affect the growth and development of plants. Most plants respond to drought with physiological and metabolic mechanisms at the molecular, cellular, and organism levels (Osmolovskaya et al., 2018). Plants mainly have different mechanisms to confront the drought stress and adapt to drought stress by inducing a variety of physiological, biochemical and morphological responses (Mirzaee et al., 2013). At the germination stage, drought stress reduces in germination by decreasing the relative water content (Dadach et al., 2015), and in plant, with decreasing relative water content, the signals from the root cause the stomata to close and photosynthesis to decrease. These chemical signals are abscisic acid (Zhou et al., 2017). Plant resistance to various stresses is related to antioxidant capacity and increased antioxidant levels that can prevent damage ( or, 2003). A stress factor or combination of abiotic stressors causes an imbalance in cellular homeostasis due to the accumulation of reactive oxygen species such as H2O2 (Kaur et al., 2018). To eliminate reactive oxygen species and to protect homeostasis, plants have a single antioxidant defence system which includes antioxidant compounds (such as ascorbate, glutathione, phenolic compounds, etc.) and enzymes (such as superoxide dismutase and catalase, etc.) are involved in the ascorbate-glutathione cycle (Ahmad et al., 2018). Drought as abiotic stress by creating oxidative stress damages photosynthetic lipids, proteins and nucleic acids, enzymes and pigments; leading to disruption of natural metabolism (Salehi Shanjani et al., 2015). Flavonoids and other phenolic compounds are a large group of secondary metabolites that usually contain at least one hydroxyl group, most of which have antioxidant properties. These compounds are non-enzymatic antioxidants and have valuable antimutation, antimicrobial and anti-cancer properties. (Kumar et al., 2013; Ahmed et al., 2016). The concerted action of low molecular weight antioxidants, similar to polyphenols (Sgherri et al., 2003) and flavonoids (Hernandez et al., 2000), can effectively scavenge harmful radicals. Phenolic acids at high levels increase the activity of many antioxidant enzymes and lipid peroxidation (Hegab, 2005). Drought stress increases the activity of superoxide dismutase and catalase enzymes, although this activity depends on the susceptibility of different cultivars and the genetic potential of the species. (Masoumi et al., 2010). The role of these two enzymes in modulating the amount of oxygen free radicals has been proven and various researchers have considered superoxide dismutase as the primary defence mechanism against oxygen free radicals (Zandalinas et al., 2017). Research on safflower under drought stress to evaluate the activity of antioxidant enzymes showed an increase in catalase activity by increasing drought levels (Sirusmehr et al., 2015). Response of yield and composition of essential oils to water stress varies with duration and severity of stress. It is reported that the production of primary metabolites and the yield of volatile oils may decrease when plants are under water stress (Panrong et al., 2006).
The alangu (Lallemantia iberica) is an annual plant, herbaceous, short with opposite leaves, belonging to the Lamiaceae family and is distributed in southwest Asia and Europe (Ursu & urcean, 2012). The results of the study by Khosravi Dehaghi et al. (2016) identify alangu as a valuable medicinal plant, containing substances responsible for various pharmacological activities. The antioxidant activity of compounds isolated from L. iberica also has been confirmed. Despite the efforts made by agricultural scientists, crop yields and efficiency are still lower than their potential. This is due to the low yields of water and nutrients used by crops and the imposition of intense competition from pests and weeds on the plant. Nanotechnology is a new scientific approach that is able to break down these barriers and is expected to increase crop yield and efficiency in the coming years and respond to the challenges of human food security. Titanium dioxide nanoparticles are among the most widely used nanomaterials but their interaction with plants has not been much investigated (Mutlu et al., 2018). Some studies have shown that nTiO2 application has positive and stimulating effects on plants ( hosh et al., 2010);
The aim of this study was to investigate the effects of titanium dioxide nanoparticles application in comparison with titanium dioxide under drought conditions on some biochemical properties of Lallemantia iberica .
MATERIALS AND METHODS
Method of Cultivation and Treatment
The same seeds were planted after being sterilized with 3% sodium hypochlorite solution, in pots of 23 cm diameter 24 cm. Irrigation up to the three-leaf stage was the same for all the treatments. After the three-leaf stage irrigation of control plants, according to field capacity, for moderate drought 75% and for severe drought 35% was done. Stress was applied sixty days after planting to harvesting stage. Plants treatment was performed in eight-leaf stage after preparation of nano titanium dioxide anatase and bulk titanium dioxide solutions (0.03% concentration). Distilled water replaced bulk and nano titanium dioxide solutions for spraying of control plants. The second treatment was performed in the flowering stage and the plant samples were harvested 14 days after the second treatment.
Determination of Antioxidant Enzyme Activity
To measure antioxidant enzymes, 0.1 g fresh leaf tissue with 2 ml potassium phosphate buffer (50 mM) with pH = 7.8, after squashing, the extract was centrifuged at 10,000 g for 10 min and the upper phase of the solution was used for enzymatic assays.
Determination of Catalase (CAT EC 1.11.1.6) Activity
The catalase activity assay was performed on the basis of a decrease in absorbance of hydrogen peroxide at 240 nm. The reaction mixture consisted of 600 µl of potassium phosphate buffer (50 mM), 300 µl of hydrogen peroxide (33 mM) and 100 µl of enzyme extract. Adding hydrogen peroxide to the reaction mixture immediately initiated the reaction and the decrease in hydrogen peroxide uptake at 240 nm wavelengths every 30 s was measured with a spectrophotometer for 2 minutes. In order to calculate the enzymatic unit, extinction coefficient of 40 mM -1cm-1 was used (Simon et al., 1974).
Determination of Superoxide Dismutase (SOD EC 1.15.1.1) Activity
Preparation of Methanol Extract
Determination of Total Phenolic Content
The total phenolic content of the extracts was measured, using the folin-ciocalteu reagent (Meda et al. , 2005). 100 µl of the extract solution was mixed with 100 µl of folin-ciocalteu reagent (50% V / V) and 2 ml of sodium carbonate solution (2%). The samples were kept at environment temperature for 30 minutes after vortex. The absorbance of the solution was measured at 720 nm wavelength compared to the control. allic acid was used to draw the standard curve and the total amount of phenolic compounds in the extract was calculated using the standard curve equation. The results were expressed in mg allic acid /g of plant fresh weight. All measurements were performed in triplicate.
Determination of Total Flavonoid Content
The aluminum chloride colorimetric method was used for the measurment of the total flavonoid content with changes (Chang et al., 2002). The standard calibration curve of quercetin was used to determine the total flavonoid content in each sample. The amount of 1.5 ml of methanol (80%), 100 ml of aluminum chloride (10%), 100 ml of potassium acetate (1 M) and 2.8 ml of distilled water mixed with 500 µl of plant extract, then was incubated for 40 minutes at room temperature and the absorbance of the solution was measured at 415 nm compared to the control. Quercetin-free solution was used as control. The total flavonoid content of the extracts was measured by placing the samples in the standard curve equation and the total flavonoid content was expressed as mg quercetin equivalent (QE)/g of plant fresh weight.
DPPH Free Radical Assay
To determine the potency of the plant extract in trapping DPPH free radicals, 2 ml of the extract was mixed with 2 ml of methanolic DPPH solution (0.004%). The tubes were kept at room temperature and dark for 30 minutes. The absorbance of the samples was measured at 517 nm in comparison to the control. Control solution consisted of 2 ml DPPH and 2 ml methanol (80%). Percent DPPH radical scavenging activity was calculated by the following equation: Acontrol adsorbed control solution at 517 nm and Asample adsorbed sample at 517 nm (Miliauskas et al. , 2004).
% I = (Acontrol - Asample) / Acontrol × 100
Determination of Phenolic Compounds of Extracts by High Performance Liquid Chromatography
The amount and type of some phenolic compounds were determined by HPLC. For this purpose, the Knuer Model 2000 device with maxi-star k-1000 quadruple pumps and the C18 EUROSPHER-10 reverse phase column with 25 cm length and 4.8 mm diameter and 5 μm particle size and a detector UV-Vis, set at 280 nm, was used. The mobile phase was water, acetic acid (2% ( and acetonitrile with a flow rate of 0.8 mm / min and an injection volume of 20 μl. The final calculation was done in µg/g of dry weight of plant.
Statistical Analysis
The results were statistically analyzed by SPSS software version 25. The mean comparisons were performed with Duncan's multiple range test and randomized complete block design was used to compare the mean of interaction effects.
RESULTS AND DISCUSSION
Catalase (CAT EC 1.11.1.6) Activity
The Results of the analysis of variance showed that nTiO 2 application and drought stress and their interaction with catalase were significant on the statistical level of 1%. The groupings of treatments showed that in the samples with nTiO 2 treatment under moderate stress the highest catalase content was observed (Fig. 1). Catalase had an effective role in signal transduction in defence and germination responses. Catalase is a titrimetric enzyme that breaks down hydrogen peroxide into water and oxygen. Catalase activity is essential to remove toxic hydrogen peroxide from various stresses and to avoid oxidative stress-related damage. (He et al. , 2014) Increased catalase activity in Nigella sativa under drought stress has been reported ( orbanli et al. , 2011). Studies on red pepper and green algae have also shown that application of nTiO 2 increased catalase activity (Kuzel & Hruby, 2012). According to the study on basil plant under drought stress, the highest catalase activity was observed under moderate drought stress at 0.03% concentration of nTiO 2 , which is in agreement with the results of the present experiment.
Superoxide Dismutase (SOD EC 1.15.1.1) Activity
The results showed that the effect of nTiO 2 , stress, and their interaction were significant at 0.01 level on superoxide dismutase activity. The highest level of superoxide dismutase was observed in samples with nTiO 2 with moderate stress (Fig. 1). Investigation of the effect of titanium dioxide nanoparticles on antioxidant enzymes in onions showed that the presence of nTiO 2 increased the activity of superoxide dismutase. The study on the Lemna showed that the activity of superoxide dismutase significantly increased with increasing nTiO 2 concentration. Increased superoxide dismutase activity can improve the antioxidant system to remove reactive oxygen species (Movafeghi et al. , 2018). The increase of this enzyme in leaf and root of Mentha pulrgium has been shown under drought conditions (Niknam & Hassanpour, 2011).
Total Phenolic Content
The effect of nTiO2 and the effect of stress and the interaction of TiO2 and stress on total phenol were significant at 0.01level. In the samples treated with nTiO2, under moderate stress the highest total phenol content was observed (Fig. 2). ased on the measurements, the total amount of phenolic compounds in the shoot of L.iberica showed a significant increase with moderate drought stress, and the presence of nTio2 increased this trend, but severe stress (35% fc) reduced these compounds to moderate stress and the application of nTio2 partially prevented it from being reduced. Drought stress caused oxidative stress in the L. iberica and plant increased compounds that are mainly in the secondary metabolite group, especially phenolic compounds, to counteract damage induced by free radicals. In different plants, the content of these substances varies due to differences in drought resistance. According to the experiments by Morello et al., (2005), as the amount of olive tree irrigation increased, the total phenolics and PAL enzyme levels in the fruit decreased; therefore, the activity of this enzyme is strongly influenced by environmental conditions that play an important role in the control of total phenolic content. An important function of phenols is their role in defence mechanisms (Solar et al., 2006). The PAL enzyme has different activity at different stages of cellular and tissue differentiation and stress conditions (Morello et al., 2005). The phenolic content of cumin shoot under drought increased significantly ( attaieb et al., 2011). In addition, Agastian et al., (2000) reported an increase in the amount of polyphenols with increasing salinity in different plant tissues. However, the total phenolic content of red pepper increased at moderate salinity levels. In Salvia officinalis, total phenolic content increased in response to nanoTiO2. The highest total phenolic content was observed in nano-exposed plants at concentrations of 100 and 200 mg / l, respectively ( horbanpour, 2015).
Total Flavonoid Content
Total flavonoid content in the shoot was clearly dependent on the soil moisture level and was observed at 75% of field capacity and this indicates the involvement of flavonoid in the response of the tested plants to soil water deficit. The lowest flavonoid content was observed at 35% of field capacity. Analysis of variance showed that the effect of nTiO2 and the effect of stress on total flavonoids were significant on the statistical level of 0.05. ut the block effect on total flavonoid is not significant (Fig. 2). Under such conditions, the application of Tio2 nanoparticles caused flavonoid accumulation, indicating the activation of the antioxidant system in response to drought stress by nTio2. The presence of nTio2 in different irrigation regimes resulted in an increase in flavonoid content into the absence of nanoparticles. Flavonoid antioxidants have a protective effect during drought stress. Many flavonoids are active components of medicinal plants that play important roles in plants as physiologically active compounds, protective agents against stresses and uptake agents (Tattini et al., 2004). There is some evidence that flavonoids in plants play a role in reducing the effects of harmful radiation and toxic substances as well as the plant's response to stress through controlling auxin transport ( everidge et al., 2007).The results showed that the presence of nTio2 increased the amount of flavonoid compounds. According to experiments on maize and wheat, the maximum flavonoid content was observed at 40% of field capacity and at least 20% of field capacity. In such conditions, application of magnesium nanoparticles caused flavonoid accumulation.
Antioxidant Activity
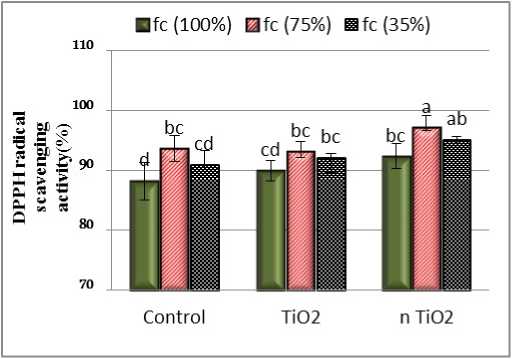
Results showed that the effect of stress and the presence of nTiO2 on antioxidant activity were significant at 0.05 level of probability. The DPPH test results showed an increase in free radical scavenging at moderate stress levels and foliar application of titanium dioxide nanoparticles improved this process (Fig. 3). In a similar study that investigated the effect of drought stress on biochemical constituents and antioxidant activity of Cumin, four system models were used to investigate the effect of drought stress on antioxidant properties. DPPH test showed that the highest IC50 was in the control and the lowest in severe drought stress. ut in the beta-carotene system model, the highest antioxidant activity was related to moderate stress treatment ( ettaieb Rebey et al., 2012). In other study, Ahmed et al., (2012) investigated the effect of drought and salinity on total phenolic, flavonoid and antioxidant activities of Lepidium sativum and similar to the present study used DPPH test to evaluate antioxidant activity. The results showed that the amount of total phenol, total flavonoid and free radical scavenging activity increased under moderate stress and decreased in severe salinity and drought stress. The antioxidant activity of plant extracts has been shown to be largely dependent on the concentration of phenolic compounds in the plant (Hiem et al., 2004). In the present study, it was found that the L. iberica was adapted to moderate drought, similar to Cumin cyminum L. ( ettaieb Rebey et al., 2012).
Results of Plant Tissue Analysis by HPLC
The effect of stress and nTiO 2 on allic acid was significant on statistical level of 1%. allic acid was measured despite moderate and even severe stress in the presence of nTiO 2 in the plant. However, in plants without nTiO 2 treatment, allic acid was not observed. p-hydroxybenzoic acid was not found in the tested plants (Table 1).
Results showed that the effect of stress on the rutin with statistical level of 0.01 was significant but the effect of TiO 2 was not significant The stress and application of nTiO 2 on rutin, which had the highest molecular weight among the compounds studied and had less antioxidant power than the other compounds, had no significant effect. Rutin was not observed in plants under severe stress, possibly due to the effect of severe drought on the enzymes of its biosynthetic pathway (Table 1).
allic acid with three hydroxyl groups showed good antioxidant activity related to the placement of the hydroxyl groups on the benzene ring. After deprotonisation, gallic acid anion is formed which has a large effect on free radical scavenging. And the results of research on antioxidant power of benzoic acid and cinnamic acid derivatives showed that gallic acid has antioxidant activity similar to caffeic acid and less than p-hydroxybenzoic acid (Velika et al., 2012). According to experiments performed by Szwajgier et al., (2005), the measurement methods used strongly influence the results obtained. In the DPPH free radical scavenging method, vanillic acid has higher antioxidant power than sinapic and ferulic acid, with the same scavenging power. Caffeic acid also appeared weaker than sinapic and ferulic acid. The results of HPLC analysis showed that vanillic acid was observed and measured only with nTiO2 in L. iberica plant. The effect of stress and nTiO2 on vanillic acid was significant at 0.01 level; As the level of drought stress increased, its content decreased and the highest amount was observed in nTiO2 treatment under non-stress conditions. Application of nTiO2 in L. iberica produced vanillic acid which it has higher antioxidant power than other compounds studied in the phenylpropanoid pathway (Table 2). The results indicated that the effect of stress and nTiO2 on caffeic acid was not significant. Caffeic acid, a polyphenol derived from hydroxycinnamic acid in the lignin biosynthesis pathway, leads to conversion of ferulic acid and Other structural components of lignin and conversion of caffeic acid to ferulic acid with the caffeate O-methyltransferase enzyme (2.1.1.68) ( oerjan et al., 2003). TiO2 nanoparticles probably converted caffeic acid to ferulic acid by affecting the activity of this enzyme in L. iberica under both stress and drought stress conditions. Ferulic acid was only observed in nTiO2-treated plants, and the effect of TiO2 and the interaction of stress and TiO2 was significant at 0.01 level (Table 2). Application of nTiO2 and the effect of stress on sinapic acid were significant at 0.01 level and the amount of sinapic acid which had lower antioxidant power than vanilic acid (Szwajgier et al.,, 2005) was increased in nTiO2 treated plants compared to control plants and the highest level was observed in the presence of nTiO2 in plants under moderate stress. Syringin was only measurable with presence of nTiO2 in comparison to untreated plants with TiO2 and the results showed that the effect of stress and nTiO2 was significant at 0.01 level on syringin. The highest content was similar to sinapic acid in nTiO2 treated plants and observed under moderate stress (Table 2).
Researches has shown that sinapic acid esters regulates the inhibition of germination, growth, and dormancy break by abscisic acid in Arabidopsis with a negative feedback cycle ( i et al. , 2017). nTiO 2 probably over-regulated the production of syringin, ferulic and sinapic acid by affecting the expression of biosynthesis pathway genes in phenylpropanoid compounds. Research on basil showed that the expression of 4-coumarate-CoA ligase (EC6.2.1.12) and cinnamate-4-hydroxylase (EC1.14.14.91) genes in the phenylpropanoid pathway decreased under severe stress and the expression of cinnamyl alcohol dehydrogenase (EC1.1.1.195) gene did not change (Abdollahi Mandoulakan, 2017). In a study by
Ongphimai et al., (2013), there was a direct relationship between total phenolic content and total antioxidant activity in phytochemical extracts of different fruits. The highest phenolic content in fruits resulted in the highest antioxidant activity, indicating that phenols may be involved in total antioxidant activity in fruits. The inhibitory potency of different extracts largely depends on the number and position of the hydroxyl groups and the molecular weight of the phenolic compounds. In lower molecular weight phenolic compounds, hydroxyl groups are easily available (Jung et al., 2006). A number of studies conducted to evaluate the antioxidant activity of benzoic acid and cinnamic acid derivatives have showed a relationship between structure and activity (Cuvelier et al., 1992). It was shown that the antioxidant potential depends on the replacement of the phenol ring with the hydroxyl groups at the ortho- and para positions and the methylation of the phenol ring in the ortho position relative to the hydroxyl groups also increases activity. The researchers also noted that the higher antioxidant activity of the cinnamic acid derivatives compared to benzoic acid could be attributed to the presence of the CH-CH-COOH group in these molecules.
Phenylpropanoids are produced as secondary metabolites of plants during developmental stages in response to stressful conditions. Among the active components of the plant defense system, aromatic phenylpropanoids play an important role in protecting plant cells against drought and heat. Drought stress can be suitable for the synthesis of phenolic compounds and indicates the regulation of the activity of related genes in response to drought. The increase in the accumulation of phenolic compounds, monoterpens, alkaloids and other compounds is consistent with the activity of key genes in their biosynthetic pathway under severe stress and is established in recent studies (Yadav et al. , 2013, Rastogi et al. , 2014, Nowak et al. , 2010). It can be concluded that the phenylpropanoid pathway in L. iberica plant is more likely to produce sinapic acid, and the application of nTiO 2 induces the expression of genes involved in the production of some enzymes of this pathway to produce syringin, sinapic and ferulic acid.
Table 1: Comparison of mean interactions of TiO 2 and nTiO 2 in L. iberica under drought stress on alic acid Rutin, Caffeic acid and Hydroxy benzoic acid.
|
Treatment |
Irrigation content |
Galic acid µg/gDw |
Rutin µg/gDw |
Caffeic acid µg/gDw |
Hydroxy benzoic acid µg/gDw |
|
no treatment |
100% FC |
15.1a±0.46 |
14.01a±0.11 |
0±0 |
0±0 |
|
75% FC |
0c±0 |
14.02a±0.14 |
0±0 |
0±0 |
|
|
35% FC |
0c±0 |
0b±0 |
0±0 |
0±0 |
|
|
TiO 2 |
100% FC |
13.62b±0.44 |
13.99a±0.14 |
0±0 |
0±0 |
|
75% FC |
0c±0 |
13.98a±0.11 |
0±0 |
0±0 |
|
|
35% FC |
0c±0 |
0b±0 |
0±0 |
0±0 |
|
|
nTiO 2 |
100% FC |
13.44b±0.46 |
14.08a±0.12 |
0±0 |
0±0 |
|
75% FC |
13.6b±0.26 |
13.95a±0.15 |
0±0 |
0±0 |
|
|
35% FC |
13.68b±0.18 |
0b±0 |
1±1.73 |
0±0 |
In each column the averages that have at least one letter in common do not differ significantly at the statistical level of 5%.
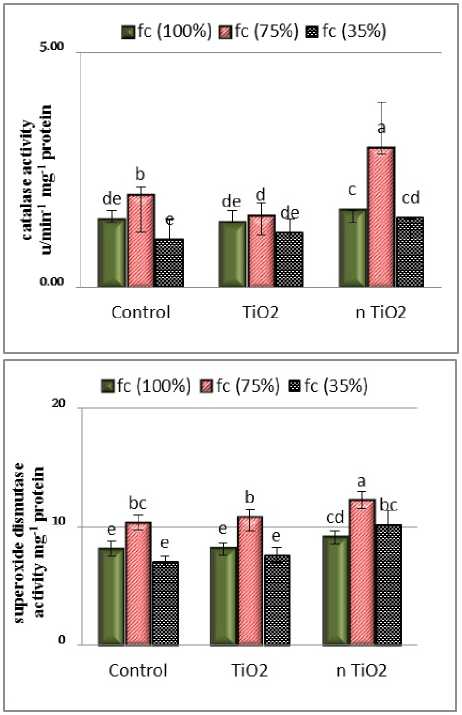
A
B
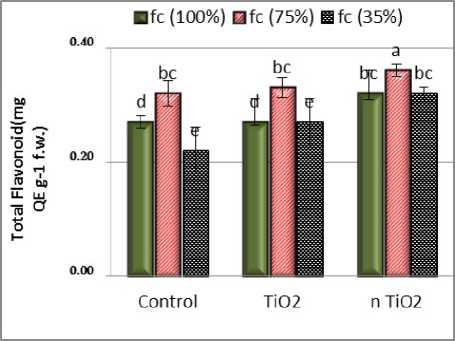
A
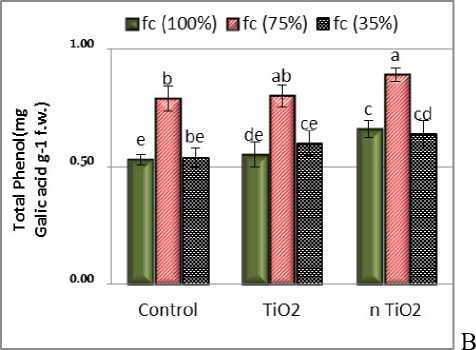
Figure 1: Interaction effects of titanium dioxide and nano titanium dioxide on catalase (A) and superoxide dismutase ( ) in shoot of L. iberica under drought stress. The averages with at least one letter in common did not differ significantly at the 0.05 level.
Figure 2: Interaction effects of titanium dioxide and nano titanium dioxide on total flavonoid (A) and total phenolic content ( ) in shoot of L. iberica under drought stress. The averages with at least one letter in common did not differ significantly at the 0.05 level.
Table 2. Comparison of mean interactions between TiO 2 and nTiO 2 in L.iberica under drought stress on vanilic acid, syringic acid, ferulic acid, and sinapic acid.
|
Treatment |
Irrigation content |
Vanilic acid µg/gDw |
Syringic acid µg/gDw |
Ferulic acid µg/gDw |
Sinapic acid µg/gDw |
|
no treatment |
100% FC |
0c±0 |
0d±0 |
0±0 |
10.33d±0.63 |
|
75% FC |
0c±0 |
0d±0 |
0±0 |
32.56b±3.06 |
|
|
35% FC |
0c±0 |
0d±0 |
0±0 |
7.59e±0.89 |
|
|
TiO 2 |
100% FC |
0c±0 |
0d±0 |
0±0 |
5.85e±0.88 |
|
75% FC |
0c±0 |
0d±0 |
0±0 |
21.72c±3.32 |
|
|
35% FC |
0c±0 |
0d±0 |
0±0 |
0f±0 |
|
|
nTiO 2 |
100% FC |
6.24a±0.28 |
44.47b±3.52 |
29.92a±2.92 |
12.96d±0.24 |
|
75% FC |
2b±0.25 |
56a±1.55 |
29.8a±0.6 |
59.89a±3.89 |
|
|
35% FC |
1.93b±0.08 |
35c±5.2 |
29.68a±3.68 |
10.53d±1.06 |
In each column the averages that have at least one letter in common do not differ significantly at the statistical level of 5%.
The amount of Potassium 208.67 ± 7.09 and amount of Sodium 17.33 ± 2.08 (Hegazy et al., 2013). The report was available showed comparison of nutrients in different medicinal plant and Na+ was found in minor level as compare to K+ (Rajurkar and Damame, 1998). Abolajiet al., (2007) reported that amount of sodium increased when fruit extract of Parinari polyandra level enhanced in mg/kg and level of potassium declined when fruit extract level of plant enhanced. The amount of vitamin C was found in Ficus palmata (2.93 ± 0.18) in the fruit (Saklani, Chandra and Mishra, 2011). haziet al., (2012) reported that content of ascorbic acid in Ficus carica 22.42 ± 0.01 mg ascorbic acid/100g and content of tocopherol found up to 1.9±0.57 mg tocopherol/100 mg. Doymaz (2005) reported that Vitamin C is abundant in figs.
CONCLUSION
This study showed that the foliar application of TiO2 nanoparticles had a significant effect on some of L. iberica 's traits, which may indicate that titanium dioxide nanoparticles penetrate the leaf more readily than titanium dioxide particles. The data presented show that application of titanium dioxide nanoparticles increased resistance of L. iberica to drought stress due to increased accumulation of protective antioxidants which in combination with increased antioxidant enzymes, total phenolic compounds, total flavonoids, antioxidant activity and produced some phenols under dry soil. Flavonoids and total phenolic compounds that were increased by moderate drought stress and the presence of TiO2 nanoparticles may indicate that L. iberica depends on large amounts of phenolic compounds to confront the negative effects of stress and antioxidant defence. Considering that L. iberica is cultivated for medicinal use and in order to achieve maximum biological yield, irrigation regime (75% FC) with 0.03 concentration of titanium dioxide nanoparticles was identified as the best treatment in this study.
References Interaction of bulk and nano titanium dioxide on antioxidant activity of Lallemantia iberica under drought conditions
- Abdollahi Mandoulakani B., Eyvazpour E. and Ghadimzadeh M. (2017). The effect of drought stress on the expression of key genes involved in the biosynthesis of phenylpropanoids and essential oil components in basil (Ocimum basilicum L.). Phytochemistry, 139, 1-7
- Agastian P., Kingsley S.J., Vivekanandan M. (2000). Effect of salinity on photosynthesis and biochemical characteristics in mulberry genotypes. Photosynthetica, 38, 287-290
- Ahmad P., Alyemeni M.N., Ahanger M.A., Wijaya L., Alam P., Kumar A., Ashraf M. (2018). Upregulation of antioxidant and glyoxalase systems mitigates NaCl stress in Brassica juncea by supplementation of zinc and calcium. J. Interact., 13, 151-162
- Ahmed A.R., Gabr A.M.M., AL- Sayed H.M.A. and Smetanska I. (2012). Effect of Drought and Salinity Stress on Total Phenolic, Flavonoids and Flavonols Contents and Antioxidant Activity in in vitro Sprout cultures of Garden cress (Lepidium sativum). Journal of Applied Sciences Research, 8(8), 3934-3942
- Bettaieb Rebey I, Jabri-Karoui I, Hamrouni-Sellami I, Bourgou S, Limam F. and Marzouk B.(2012). Effect of drought on the biochemical composition and antioxidant activities of cumin (Cuminum cyminum L.) seeds. Ind Crops Prod, 36(1), 238-245
- Beveridge C.B., Mathesius U., Rose R.J. and Gresshoff P.M. (2007). Common regulatory themes in meristem development and whole-plant homeostasis. Curr Opin Plant Biol., 10(1), 44-51
- Bi B., Tang J., Han S., Guo J. and Miao Y.(2017). Sinapic acid or its derivatives interfere with abscisic acid homeostasis during Arabidopsis thaliana seed germination. BMC Plant Biol. 17: 99
- Boerjan W., Ralph J. and Baucher M. (2003). Ligninbiosynthesis". Annual Review of Plant Biology, 54, 519-546
- Chang C.C., Yang M.H., Wen H.M. and Chern J.C.(2002). Estimation of total flavonoid content in propolis by two complementary colorimetric methods. Journal of Food and Drug Analysis, 10, 178-182
- Chinnamuthu C.R. and Murugesa Boopathi P. (2009). Nanotechnology and Agroecosystem, Madras Agricultural Journal, 96, 17-3
- Cuvelier M.E., Richard H. and Berset C. (1992). Comparison of the antioxidative activity of some acid-phenols: structure- activity relationship. Biosci. Biotech. Biochem. 56 (2), 324-325
- Dadach,M., Mehdadi,Z. and Latreche, A.( 2015). Germination Responses of Marrubium Vulgare L. under Various Waterstress Conditions. J. Appl. Environ. Biol. Sci., 5(9), 28-33
- Dixon R.A. and Paiva N. (1995). Stress-Induced phenylpropanoid metabolism. American Society of Plant Physiologist, 7, 1085-1097
- Ghorbanli M., Bakhshi Khaniki G. R., Salimi Elizei S. and Hedayati M. (2011). Effect of water deficit and its interaction with ascorbate on proline, soluble sugars, catalase and glutathione peroxidase amounts in Nigella sativa L. Iranian Journal of Medicinal and Aromatic Plants, 26(4), 466- 476
- Ghorbanpour M. (2015).Major essential oil constituents, total phenolics and flavonoids content and antioxidant activity of Salvia officinalis plant in response to nano-titanium dioxide. Ind J Plant Physiol, 20(3), 249-256
- Ghosh M., Bandyopadhyay M. and Mukherjee, A. (2010). Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels. plant and human lymphocytes. Chemosphere, 81, 1253-1262
- Giannopolitis, C.N. and Ries, S.K. (1977). Superoxide dismutase I. Occurrence in higher plants. Plant Physiol., 59, 309-314
- Haghighi M. and Teixeira da Silva, J.A. (2014). The effect of N-TiO2 on tomato, onion, and radish seed germination. Journal of Crop Science and Biotechnology, 17(4), 221-227
- He H., Vidigal D., Snoek L.B., Schnabel S., Nijveen H., Hilhorst H. and Bentsink L. (2014). Interaction between parental environment and genotype affects plant and seed performance in Arabidopsis. Journal of Experimental Botany, 65(22), 6603-6615
- Hegab M. M. (2005). Assessment of the Allelopathic Effect of Some Phenolic Compounds on Some Physiological Processes of Pea Plant (Pisum sativum). PH.D. Thesis, Faculty of Science, Beni-Sueif University, Egypt
- Hernandez J.A., Jimenez A., Mullineaux P. and Sevilla F. (2000). Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ., 23, 853-862
- Hiem K., Taigliaferro A. and Bobilya, D. (2002). Flavonoids antioxidants: chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem., 13, 572-584
- Hong F., Zhou J., Liu C., Yang F., Wu C., Zheng C. and Yang P. (2005). Effect of nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biological trace element research, 105, 269-279
- Jung C.H., Seog H.M. Choi I.W., Park M.W. and Cho H.Y. (2006). Antioxidant properties of various solvent extracts from wild ginseng leaves. LWT, 39, 266-274
- Kaur H., Sirhindi G., Bhardwaj R., Alyemeni M.N., Siddique K.H.M. and Ahmad P. (2018). 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep., 8(1), 8735
- Khosravi-Dehaghi N., Gohari A.R., Sadat-Ebrahimi S.S., Naghdi-Badi H. and Amanzadeh Y. (2016). Phytochemistry and antioxidant activity of Lallemantia iberica aerial parts. Research Journal of Pharmacognosy, 3(3), 27-34
- Kiapour H., Moaveni P., Habibi D. and Sani, B. (2015). Evaluation of the application of gibbrellic acid and titanium dioxide nanoparticles under drought stress on some traits of basil (Ocimum basilicum L.). International Journal of Agronomy and Agricultural Research (IJAAR), 6(4), 138-150
- Lei Z., Su M.Y., Wu X.C, Qu C.X. Chen L., Huang H., Liu X.Q. and Hong F.S. (2008). Antioxidant stress is promoted by nano-anatase in spinach chloroplasts under UV-Beta radiation. Biological Trace Element Research, 121(1), 69-79
- Masoumi H., Masoumi M., Darvish F., Daneshian J., Nourmohammadi G., and Habibi D. (2010). Change in several antioxidant enzymes activity and seed yield by water deficit stress in soybean (Glycine max L.) cultivars. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 38(3), 86- 94
- Meda A., Lamien C.E., Romito M., Millogo J. and Nacoulma O.G. (2005). Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chemistry, 91, 571-577
- Miliauskas G., Venskutonis P.R. and van Beek T.A. (2004). Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem., 85, 231-237
- Mirzaee M., Moieni A. and Ghanati F. (2013). Effects of drought stress on the lipid peroxidation and antioxidant enzyme activities in two canola (Brassica napus L.) cultivar. Journal of Agricultural Science and Technology, 15, 593-602
- Morello J.R., Romero M.P., Ramo T. and Motilva M.J. (2005). Evaluation of L-phenylalanine ammonia-lyase activity and phenolic profile in olive drupe (Olea europaea L.) from fruit setting period to harvesting time. Plant Science, 168, 65-72
- Movafeghi A., Khataee A., Abedi M., Tarrahi R., Dadpour M. and Vafaei F. (2018). Effects of TiO2 nanoparticles on the aquatic plant Spirodela polyrrhiza: Evaluation of growth parameters, pigment contents and antioxidant enzyme activities. Journal of Environmental Sciences, 64, 130-138
- Mutlu F., Yurekli F., Mutlu B., Bilge Emre F., Okusluk F. and Ozgul O. (2018). Assessment of Phytotoxic and Genotoxic Effects of Anatase Tio2 Nanoparticles On Maize Cultivar by Using Rapd Analysis. Fresenius Environmental Bulletin, 27 (1), 436-445
- NiknamV., Hassanpour H. (2011). Effects of penconazole and water deficit stress on physiological and antioxidative responses in pennyroyal (Mentha pulegium L.). Acta Physiol Plant, 34, 1537-1549
- Nowak M., Manderscheid R., Weigel H.J., Kleinwachter M. and Selmar D. (2010) Drought stress increases the accumulation of monoterpenes in sage (Salvia officinalis), an effect that is compensated by elevated carbon dioxide concentration. Journal of Applied Botany and Food Qualilty, 83, 133-136
- Ongphimai N., Lilitchan S., Aryusuk K., Bumrungpert A. and Krisnangkura K. (2013). Phenolic Acids Content and Antioxidant Capacity of Fruit Extracts from Thailand. Chiang. Mai J. Sci., 40(4), 636-642
- Osmolovskaya N., Shumilina J., Kim A., Didio A., Grishina T., Bilova T., Keltsieva O.A., Zhukov V., Tikhonovich I., Tarakhovskaya E., Frolov A. and Wessjohann L.A. (2018). Methodology of Drought Stress Research: Experimental Setup and Physiological Characterization. Int. J. Mol. Sci., 19(12), 4089
- Rastogi S.h., Kumar R., Chanotiya C.S., Shanker K., Gupta M.M., Nagegowda D.A. and Shasany A.K. (2013) 4-Coumarate: CoA Ligase Partitions Metabolites for Eugenol Biosynthesis. Plant Cell Physiology, 54, 1238-1252
- Sgherri C., Cosi E. and Navaro-Izzo F. (2003). Phenols and antioxidative status of Raphanus sativus grown in copper excess. Plant Physiol., 118, 21-28
- Simon J.R., Martin D.L. and Kroll M. (1974). Sodium-dependent efflux and exchange of GABA in synaptosomes, J. Neurochem., 23, 981-991
- Sirousmehr A., Bardel J. and Mohammadi S. (2015). Changes of Germination Properties, Photosynthetic Pigments and Anti Oxidant Enzymes Activity of Safflower as Affected by Drought and Salinity Stresses. Journal of Crop Ecophysiology, 8(4), 517-533
- Solar A., Colaric M., Usenik V. and Stampar F. (2006). Seasonal variations of selected flavonoids, phenolic acids and quinines in annual shoots of common walnut (Juglans regia L.). Plant Science, 170, 453-461
- Szwajgier D., Pielecki J. and Targonski Z. (2005). Antioxidant activities of Cinnamic and Benzoic acid derivatives. Acta Sci. Pol., Technol. Aliment, 4(2), 129-142
- Tattini M., Galardi C., Pinelli P., Massai R., Remorini D. and Agati, G. (2004). Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytologist, 163, 547-561
- Velika B. and Kron I. (2012).Antioxidant properties of benzoic acid derivatives against superoxide radical. Free Radicals and Antioxidants, 2(4), 62-67
- Yadav R., Sangwan R., Sabir F. and Srivastava A. (2013). Effect of prolonged water stress on specialized secondary metabolites, peltate glandular trichomes, and pathway gene expression in Artemisia annua L. Plant Physiology and Biochemistry, 74, 70-83
- Zandalinas S. I., Balfagón D., Arbona V. and Gómez-Cadenas A. (2017). Modulation of Antioxidant Defense System Is Associated with Combined Drought and Heat Stress Tolerance in Citrus. Frontiers in Plant Science, 8, article 953
- Zhou R., Yu X., Ottosen C. O., Rosenqvist E., Zhao L., Wang Y. and Wu Z. (2017). Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biology, 17(24), 1-13
