Ископаемые кости миоцен-голоценовых верблюдов Евразии (Россия): минералого-геохимические свойства и экологические реконструкции
: минералого-геохимические свойства и экологические реконструкции")
Автор: Силаев В. И., Титов В. В., Тесаков А. С., Филиппов В. Н., Васильев Е. В., Смолева И. В., Шанина С. Н., Киселева Дарья Владимировна, Хазов А. Ф., Макеев Б. А., Солошенко Н. Г., Окунева Т. Г.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 9 (345), 2023 года.
Бесплатный доступ
С использованием широкого комплекса минералого-геохимических и изотопно-геохимических методов исследована уникальная серия проб костных остатков ископаемых верблюдов, отобранных из местонахождений на территории Евразии от Северо-Западного Причерноморья до Западного Забайкалья и Монголии, датированных в хронологическом диапазоне от позднего миоцена (6 млн л.) до XIV века н. э. Полученные результаты указывают на минералого-геохимические свойства ископаемых костей как на весьма эффективный способ эколого-климатических реконструкций и расшифровки истории эволюции млекопитающих.
Ископаемые верблюды, евразия, минералого-геохимические свойства костей, экологические реконструкции
Короткий адрес: https://sciup.org/149144387
IDR: 149144387 | УДК: 569+54.021+549 | DOI: 10.19110/geov.2023.9.1
Fossil bones of the Miocene-Holocene camels of Eurasia (Russia): mineralogical and geochemical properties and ecological reconstructions
Using a wide complex of mineralogical-geochemical and isotope-geochemical methods we studied, a unique series of samples of fossil camels' bone remains taken from Eurasian localities from the northwestern Black Sea region to western Transbaikalia and Mongolia, dated in the chronological range from the Late Miocene (6 Ma) until the 14th century AD. The obtained results indicated the mineralogical and geochemical properties of fossil bones as very effective way of ecological and climatic reconstructions and interpretation of the evolutionary history of mammals.
Текст научной статьи Ископаемые кости миоцен-голоценовых верблюдов Евразии (Россия): минералого-геохимические свойства и экологические реконструкции
Современные верблюды являются обитателями крайне аридных условий. Благодаря этому при палеореконструкциях наличие какого-либо таксона мозоленогих в ископаемых фаунах Евразии некоторые исследователи используют в качестве показателя крайне засушливых ландшафтов. Верблюды также являются важными индикаторами палеогеографических событий на территории Евразии — межконтинентальных фау-
Посвящается Международному году верблюдовых (2024) нистических обменов, аридизации ландшафтов в среднем и позднем плейстоцене.
Проникновение представителей сем. Camelidae на территорию Евразии произошло в позднем миоцене (мессиний/понт; MN 13) около 6—5.5 млн л. н., когда в результате первого заметного похолодания в преддверии плейстоцена произошло падение уровня Мирового океана. После проникновения через Беренгийский мост верблюды, отнесённые к роду
Paracamelus , широко распространились по всей территории Старого Света: их костные остатки в составе гиппарионовых фаун известны от Средней Азии до Испании (Алексеева, 1974; Аубекерова, 1974; Made, Morales, 1999; Martini, Geraads, 2018; Titov, Logvinenko, 2006; Titov, 2008a; Liu et al., 2023). На данный момент описано два таксона позднемиоценовых верблюдов: P. aguirrei Morales, 1984 и P. giui Liu, Hou, Zhang, 2023.
В плиоцене верблюды рода Paracamelus активно адаптировались к особенностям существования на различных территориях Евразии и Африки, дав начало целому ряду видов. В частности, в Северном и Южном Причерноморье, Приазовье был обычным P. alexejevi Havesson, 1950 (Хавесон, 1954; Свистун, 1971). Известны также P. praebactrianus Orlov, 1927, P. longipes (Aubekerova, 1974), обнаруженные в разных частях Средней Азии, и P . sp. в Северной Африке (Orlov, 1929, 1930; Хавесон, 1954; Алексеева, 1974; Likius et al., 2003). На территории Индии из формации Пинжор верхнего сивалика описан «Camelus» sivalensis Falconer & Cautley, 1836, который, скорее всего, тоже является представителем рода Paracamelus .
В начале плейстоцена (гелазий, MNQ 17—18) на территории Причерноморья были достаточно обычными P. alutensis Stefanescu, 1895, являвшиеся измельчавшими потомками P. alexejevi. Из раннего плейстоцена Таджикистана описан P. trofimovi Sharapov, 1986. При этом достаточно широко в пределах Евразии на протяжении раннего плейстоцена от территории Китая до Северного Причерноморья был распространён P. gi-gas Schlosser, 1903.
Начало среднего плейстоцена на территории Северной Евразии характеризовалось значительными ландшафтными и растительными перестройками. В это время широкое распространение получают степные пространства, пришедшие на смену саванноподобных лесостепных ландшафтов. Предполагается, что в это время здесь повсеместно вымирают представители р. Paracamelus , и только в конце среднего плейстоцена в Азии и Восточной Европе становятся обычными верблюды рода Camelus . Самым известным представителем этого рода в Евразии является C. knoblochi Nehring, 1901, ареал распространения которого простирался от Северо-Восточного Китая и Забайкалья до Азовского моря (Titov, 2008b; Liu et al., 2023), а время существования охватывало период от конца среднего до начала позднего плейстоцена (MIS 9-5).
В период последнего оледенения верблюд Кноблоха в Северной Евразии сменился на двугорбого верблюда C. ferus Przewalski, 1878 (= C. bactrianus L., 1758). Западная граница распространения диких двугорбых верблюдов C. ferus в позднем плейстоцене, по имеющимся на данный момент данным, располагалась в Поволжье (Титов, Головачёв, 2020). В более западные районы на территорию Восточной Европы верблюды попали уже в одомашненном виде C. bactrianus с караванами из Средней Азии начиная с V—III вв. до н.э. (Титов, 2009; Titov, 2011). Не исключено, что проживание современных верблюдов в областях с аридным климатом является результатом их вытеснения из более комфортных по климату и кормовой базе областей более многочисленными и конкурентоспособными видами копытных (Titov, 2008). Считается, что ископаемые евразийские верблюды обитали не столько в ус- ловиях засушливых ландшафтов — полупустынь и пустынь, сколько в условиях саванн, степей и лесостепей (Titov, Logvinenko, 2006).
Несколько иначе проходила эволюция мозоленогих в Передней Азии и Африке. Уже в конце плиоцена — начале плейстоцена в Африке, судя по всему, появляются представители рода Camelus , базальным видом которого является C. grattardi Geraads, 2014 (Geraads, 2014; Geraads et al., 2021). После проникновения в Азию потомки этого вида, вероятно, и дали начало линии, ведущей к появлению двугорбых верблюдов C . ferus / bactrianus (Rowan et al., 2019; Geraads et al., 2021). В конце раннего и начале среднего плейстоцена на территории Северной Африки обитал C. thomasi Pomel, 1893, который, скорее всего, тоже не является прямым предком одногорбого верблюда C. dromedarius L., 1758. На протяжении плейстоцена в Африкано-Аравийском регионе шла активная эволюция мозоленогих. В частности, в отложениях плейстоцена Сирии обнаружены остатки до пяти разных представителей верблюдов (Martini et al., 2015). Есть мнение, что эволюционные ветви одногорбых и двугорбых верблюдов р. Camelus , которые дожили до наших дней, разошлись в эволюционном плане в промежутке между 1 и 2 млн лет назад (Geraads et al., 2020).
Несмотря на активное использование метода реконструкции условий существования древних животных по содержанию стабильных изотопов в их костях и зубах, для верблюдов Евразии этого не было сделано до сих пор. Есть лишь серия работ, в которых в той или иной степени рассматриваются вопросы реконструкции диеты североамериканских и южноамериканских Camelidae с использованием изотопно-геохимических данных (MacFadden and Cerling, 1996; MacFadden and Shockey, 1997; Connin et al., 1998; Feranec and MacFadden, 2000; Feranec, 2003; Kohn et al., 2005; Ruez, 2005; Feranec and MacFadden, 2006; Hoppe and Koch, 2006; Vetter, 2007; Higgins and MacFadden, 2009; Nunez et al., 2010; Domingo et al., 2012; Kohn and McKay, 2012; Pérez-Crespo et al., 2012; Kita et al., 2014; Trayler et al., 2015). В частности, было показано, что для наиболее распространённых трёх родов плейстоценовых американских верблюдовых Camelops , Hemiauchenia и Palaeolama было характерно питание побегами деревьев и кустарников (browser) или смешанное питание (mixed feeders) (Yann et al., 2016). Имеются также результаты изотопных исследований, полученные по эмали зубов современных одногорбых верблюдов и некоторых плиоплейстоценовых верблюдов Camelus Африки (Harris et al., 2010).
Основная проблема современной палеонтологии ископаемых верблюдов Евразии состоит в практически полном отсутствии исследований их костных остатков минералого-геохимическими методами, уже хорошо зарекомендовавшими себя в приложении к разнообразной фауне млекопитающих, существовавшей в весьма широком хронологическом диапазоне (Силаев и др., 2016; Silaev et al., 2017; Силаев и др., 2019, 2021, 2022, 2923). Объектами наших исследований послужили костные фрагменты верблюдов, отобранные в 13 местонахождениях от Северо-Западного Причерноморья до Западного Забайкалья и Монголии и в хронологическом диапазоне от позднего миоцена (6 млн л.) до XIV века н. э. (рис. 1).
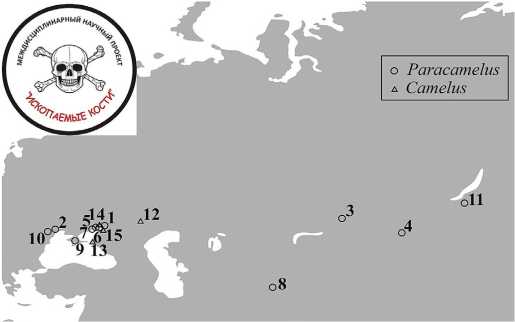
Рис. 1. Места находок костных остатков верблюдов, использованных в данном исследовании:
-
1 — Синявская, 2 — Одесские катакомбы, 3 — Острая Сопка, 4 — Санган Далай-Нур, 5 — Ливенцовка, 6 — Морская, 7 — Весёло-Вознесенка, 8 — Куруксай, 9 — Таврида, 10 — Порумбрей, 11 — Чикой, 12 — Нижнее Поволжье, Чёрный Яр, 13 — Волна-1, 14 — Танаис, 15 — Азак
Fig. 1 . Localities of camels’ bone remains used in this study: 1 — Sinyavskaya, 2 — Odessa catacombs, 3 — Ostraya Sopka, 4 — Sangan Dalai-Nur, 5 — Liventsovka, 6 — Morskaya, 7 — Veselo-Voznesenka, 8 — Kuruksay, 9 — Taurida, 10 — Porumbrey, 11 — Chikoy, 12 — Lower Volga, Cherniy Yar, 13 — Volna-1, 14 — Tanais, 15 — Azak
Топохронологически эту коллекцию можно подразделить на две серии: 1) топологическую — восточноевропейскую (местонахождения 1, 2, 5, 6, 7, 9, 10, 12, 13—15), среднеазиатско-западносибирскую (3, 8) и монголо-восточносибирскую (4, 11); 2) хронологическую — миоцен-плиоценовую (местонахождения 1, 10, 13), плейстоценовую (2, 4, 5, 7—9, 11, 12) и голоценовую (3—5, 6). Таким образом, исследуемая коллекция ископаемых костей охватывает большую часть евразийского пространства и практически все время существования на нем верблюдов, включая при этом несколько периодов значительных трансформаций экосистем и животных сообществ Евразии — в конце миоцена (6—5.5 млн л. н.), в конце раннего плейстоцена (0.7—0.8 млн л. н.) и позднего плейстоцена (71—11 тыс. л. н.).
Объекты и методы исследований
Непосредственными объектами исследований послужили фрагменты (21 экз.) трубчатых костей конечностей, нижних челюстей, ребер (табл. 1), варьирующих по окраске от бурых и буроватых (более древние) до светло-серых и бесцветных (рис. 2—4). Размеры фрагментов, представленные для анализа, имели несколько разные размеры, возрастающие в направлении от наиболее древних к наиболее молодым костям (мм):
Таблица 1 . Реестр исследованных костей ископаемых евразийских верблюдов
Table 1. Register of studied bones of fossil Eurasian camels
|
Обр. Sample |
Таксон Taxon |
Часть скелета Sceleton part |
Возраст Age |
Местонахождение Location |
|
Врб-1 |
Paracamelus cf . aguirrei |
метаподия metapodium |
поздний миоцен (понт) (5.5 млн л.) Late Miocene (Pontian) (5.5 Ma) |
Ростовская обл., ст. Синявская Rostov region, st. Sinyavskaya |
|
Врб-2 |
Paracamelus praebactrianus |
фаланга II phalanx II |
плиоцен (3.5 млн лет) Pliocene (3.5 Ma) |
Острая Сопка, р. Иртыш Ostraya Sopka, Irtysh riv. |
|
Врб-3 |
Paracamelus alexejevi |
диафиз большеберцовой кости tibial diaphysis |
плиоцен (3.5 млн лет) Pliocene (3.5 Ma) |
Одесские катакомбы (карстовые воронки) Odessa catacombs (sinkholes) |
|
Врб-4 |
Paracamelus sp. |
метаподия metapodium |
плиоцен (3.5 млн лет) Pliocene (3.5 Ma) |
Западная Монголия, Санган_Далай-Нур Western Mongolia, Sangan_Dalai-Nur |
|
3.1 |
Paracamelus cf. praebactianus |
передняя метаподия anterior metapodium |
плиоцен (3.5 млн лет) Pliocene (3.5 Ma) |
Острая Сопка, р. Иртыш Ostraya Sopka, Irtysh riv. |
|
Тит-1 |
Paracamelus alutensis |
большая берцовая кость tibia |
палеоплейстоцен (гелазий) (2.5—2.2 млн лет) Paleopleistocene (Gelasian) (2.5—2.2 Ma) |
Ростовская обл., Неклиновский р-н, с. Весёло-Вознесенка Rostov region, Neklinovsky district, village of Veselo-Voznesenka |
|
Тит-2 |
Paracamelus alutensis |
нижняя челюсть lower jaw |
» |
Ростовская обл., Неклиновский р-н, ст. Морская Rostov region, Neklinovsky district, st. Morskaya |
|
Тит-3 |
Paracamelus alutensis |
нижняя челюсть lower jaw |
» |
Ростовская обл., Мясниковский р-н, Ливенцовка Rostov region, Myasnikovsky district, Liventsovka |
|
Врб-6 |
Paracamelus cf. gigas |
передняя метаподия anterior metapodium |
» |
Ростовская обл., Мясниковский р-н, Ливенцовка Rostov region, Myasnikovsky district, Liventsovka |
|
Врб-7 |
Paracamelus alutensis |
нижняя челюсть lower jaw |
» |
Ростовская обл., Мясниковский р-н, Ливенцовка Rostov region, Myasnikovsky district, Liventsovka |

Окончание таблицы 1 / End of Table 1
|
Обр. Sample |
Таксон Taxon |
Часть скелета Sceleton part |
Возраст Age |
Местонахождение Location |
|
Врб-5 |
Paracamelus trofimovi |
задняя метаподия posterior metapodium |
» |
Таджикистан, Куруксай Tajikistan, Kuruksay |
|
Тавр-1 |
Paracamelus gigas |
задняя метаподия posterior metapodium |
ранний плейстоцен (калабрийский ярус) (1.8—1.5 млн л) Early Pleistocene (Calabrian stage) (1.8—1.5 Ma) |
Крым, пещера Таврида Crimea, Tavrida cave |
|
Тавр-2 |
Paracamelus gigas |
передняя метаподия anterior metapodium |
» |
Крым, пещера Таврида Crimea, Tavrida cave |
|
Врб-8 |
Camelus knoblochi |
диафиз плечевой кости humeral diaphysis |
конец среднего — начало позднего плейстоцена (120—80 тыс. л. н.) end of Middle — beginning of Late Pleistocene (120—80 thousand years ago) |
Нижнее Поволжье Lower Volga region |
|
Врб-9 |
Camelus knoblochi |
передняя метаподия front metapodium |
» |
Забайкалье, р. Чикой Transbaikalia, Chikoy riv. |
|
Врб-10 |
Paracamelus alexejevi |
метаподия metapodium |
плиоцен (3.5 млн л. н) Pliocene (3.5 Ma) |
Молдавия, Чимишлийский р-он, пос. Порумбрей Moldova, Cimislia district, village of Porumbray |
|
3.4 |
Camelus cf . ferus |
ребро rib |
поздний плейстоцен (42 тыс. л. н.) Late Pleistocene (42 thousand years ago) |
Астраханская обл., Чёрный Яр Astrakhan region, Black Yar |
|
Тит-4 |
Camelus bactrianus |
большеберцовая кость tibial bone |
голоцен (V век н. э.) Holocene (V A. D.) |
Ростовская обл., Неклиновский р-н, Танаис Rostov region, Neklinovsky district, Tanais |
|
Тит-5 |
Camelus bactrianus |
плечевая кость brachial bone |
голоцен (III век до н. э. — III век н. э.) Holocene (III B. C. — III A. D.) |
Таманский п-ов, Краснодарский край, Волна-1 Taman Peninsula, Krasnodar region, Volna-1 |
|
Тит-6 |
Camelus bactrianus |
нижняя челюсть lower jaw |
голоцен (XIV век н. э.) Holocene (XIV A. D.) |
Ростовская обл., г. Азов, Азак Rostov region, Azov, Azak |
|
Тит-7 |
Camelus bactrianus |
плечевая кость |
» |
Ростовская обл., г. Азов, Азак Rostov region, Azov, Azak |

Рис. 2. Образцы костей ископаемых верблюдов серии ВРБ: позднемиоценовых (ВРБ-1); плиоценовых (ВРБ-2, 3, 4); плейстоценовых гелазийских (ВРБ-5, 6, 7); плейстоценовых чибанийских (ВРБ-8, 9, 10)
Fig. 2 . Bone samples of fossil camels of the VRB series: Late Miocene (VRB-1); Pliocene (VRB-2, 3, 4); Pleistocene, Gelasian (VRB-5, 6, 7); Pleistocene, Chibanian (VRB-8, 9, 10)

Рис. 3. Образцы костей верблюдов плейстоцен-калабрийского возраста из пещеры Таврида
Fig. 3 . Pleistocene-Calabrian camel bone specimens age from Taurida cave

Рис. 4. Образцы костей ископаемых верблюдов серии ТИТ: плейстоцен-гелазийских (ТИТ-1, 2, 3) и голоценовых (ТИТ-4, 5, 6)
Fig. 4 . Samples of bones of fossil camels of the TIT series: Pleistocene-Gelasian (TIT-1, 2, 3) and Holocene (TIT-4, 5, 6)
миоцен-плиоценовые = (24 ± 110) × (15 ± 4) × (8 ± 2); палеоплейстоценовые (гелазийский и калабрийский века) = (36 ± 20) × (22 ± 10) × (11 ± 5); неоплейстоцено-вые (чибанийский и позднеплейстоценовый века) = (50 ± 7) × (17 ± 3) × (9 ± 2). Как видно из приведенных данных, размеры костных остатков в направлении удрев-нения сокращаются в 2—3 раза, сопровождаясь увеличением вариаций в 1.5—2 раза.
В ходе исследований костей ископаемых верблюдов применялся широкий комплекс современных аналитических методов: термический анализ (DTG-60А/60 АН, Shimadzu); определение содержания Сорг методом кулонометрического титрования; рентгенофлюоресцентный анализ (XRD-1800 Shimadzu); оптическая микроскопия (комплекс OLYMPUS BX51); рентгеновская дифрактометрия (XRD-6000); аналитическая растровая электронная микроскопия (JSM-6400 Jeol; Tescan Vega); определение нанопористости по кинетике ад-сорбции/десорбции азота (Nova 1200e, Quantachrome Instruments); масс-спектрометрия с индуктивно связанной плазмой (NexION 300S Perkin Elmer); инфракрасная спектроскопия (ФТ-2 Инфралюм); газохроматографическое определение аминокислотного состава в органических веществах (GC-17A); рамановская спектроскопия (Renishaw InVia, лазеры 514 и 785 нм); масс-спектрометрический анализ изотопного состава стронция (МК-ИСП-МС Neptune Plus), а также углерода, кислорода в биоапатите и углерода, азота в костном коллагене (Delta V Advantage с аналитическим комплексом, Thermo Fisher Scientific).
ИСП-МС-анализы на микроэлементы и определение изотопного состава стронция выполнены в ЦКП «Геоаналитик» Института геологии и геохимии УрО РАН в рамках госбюджетной темы № 123011800012-9. Спектры КР (рамановские) были получены в Санкт-Петербургском горном университете. Все остальные аналитические работы осуществлены в ЦКП «Геонаука» Института геологии ФИЦ Коми НЦ УрО РАН.
Микростроение и ультрапористость костей
Исследованные образцы костей голоценового возраста характеризуются хорошей сохранностью как компактной, так и губчатой частей костной ткани (рис. 5). Первая характеризуется плотным пластинчатым строением с хорошо сохранившейся тонкой гаверсовой системой с диаметром остеона в 150—200 мкм. Губчатая ткань имеет типичное ячеистое строение с хорошо сохранившимися трабекулами. Биоапатит не обнаруживает оптических преобразований. В более древних — плиоцен-плейстоценовых — костях плотная и губчатая костные ткани явно деградированы с образованием каверн и трещин, часто заполненных новообразованными минералами, особенно оксигидроксидами железа (рис. 6). Биоапатит в таких костях подвержен коллофа-низации с потерей прозрачности и приобретением на таких участках буроватой окраски.
Важной микроструктурной характеристикой костей как органоминерального композита является их ультрапористость в нанометровом диапазоне.

Рис. 5 . Микроскопическое строение костей голоценовых верблюдов. Оптико-микроскопические изображения в режимах проходящего света (a, c, e) и скрещенных николей (b, d, f)
Fig. 5 . Microscopic structure of the bones of Holocene camels. Optical-microscopic images in the modes of transmitted light (a, c, e) and crossed nicols (b, d, f)
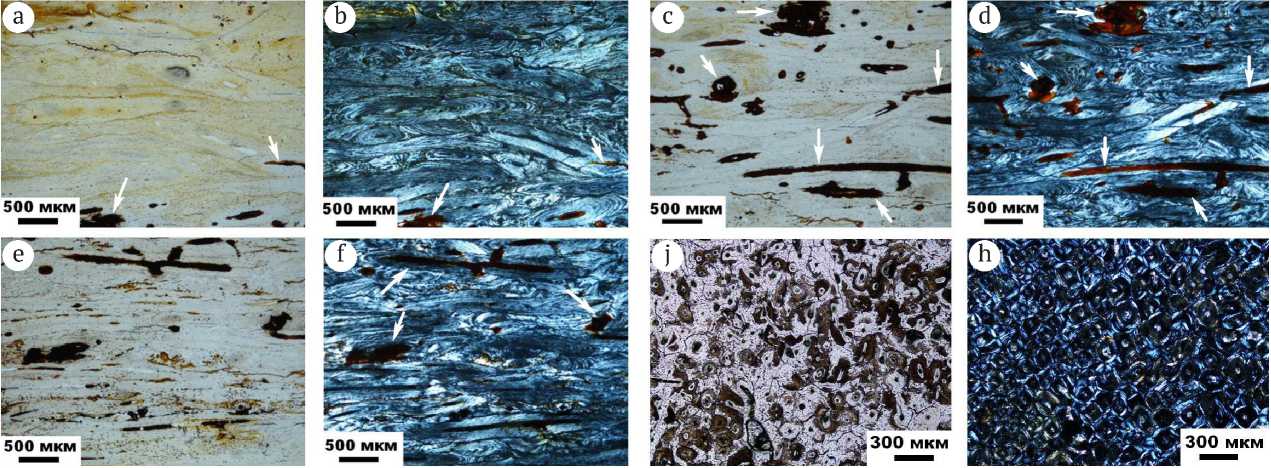
Рис. 6. Микроскопическое строение костей плиоцен-плейстоценовых верблюдов. Оптико-микроскопические изображения в режимах проходящего света (a, c, e, g) и скрещенных николей (b, d, f, h). Стрелками показаны выделения оксигидроксидов железа
Fig. 6 . Microscopic structure of bones of Pliocene-Pleistocene camels. Optical-microscopic images in the modes of transmitted light (a, c, e, g) and crossed nicols (b, d, f, h). The arrows indicate the release of iron oxyhydroxides
Проведенные исследования показали, что исследованные верблюжьи кости по состоянию нанопористости ближе всего согласуются с костями плейстоценовой мамонтовой фауны (рис. 7). Сравнение данных внутри коллекции верблюжьих костей разного возраста показывает, что кости плиоцен-плейстоценового возраста системно отличаются от голоценовых костей как большим совокупным объемом нанопор, так и пропорционально большим их условным числом (lgNo). Это демонстрирует результаты наноструктурной деградации костных остатков верблюдов в ходе их фоссилизации. Однако при этом в сравнении с близковозрастными и костями млекопитающих с Омского Прииртышья и с археологических стоянок Озерновской и Ушбулак кости верблюдов, включая наиболее древние, обнаруживают гораздо меньшую степень вторичных изменений.
Химический и нормативно-минеральный состав костей, микроминеральные примеси
В химическом составе исследованных костей (табл. 2) содержание неапатитовых примесей колеблется в довольно узких пределах — от 3.5 до 8.23 мас. %, возрастая по мере удревнения костей. В целом это значение в 1.2—3 раза уступает содержанию неапатито-
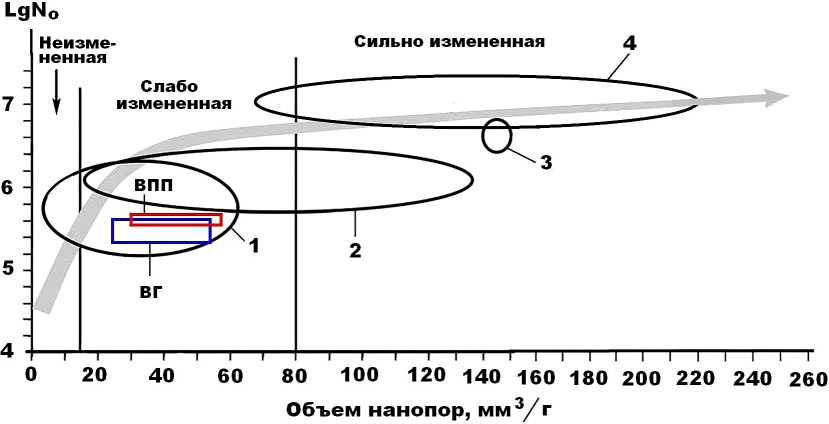
Рис. 7. Нанопористость ископаемых костей евразийских верблюдов (ВПП — плиоцен-плейстоценовых, ВГ — голоценовых) на фоне аналогичных данных, полученных нами для мамонтовой фауны с Печорского Приуралья (1) и Омского Прииртышья (2) для плейстоценовых лошадей со стоянки Заозерье в Пермской области (3) и плейстоценовых млекопитающих со стоянки Ушбулак в Восточном Казахстане (4). Заливкой показан генеральный тренд изменения нанопористости ископаемых костей в ходе их фоссилизации
Fig. 7. Nanoporosity of fossil bones of Eurasian camels (VPP — Pliocene-Pleistocene, VG — Holocene) against the background of similar data obtained by us for the mammoth fauna from the Pechora Cis-Urals (1) and the Omsk Irtysh region (2), for Pleistocene horses from the Zaozerye site in the Perm Region (3) and Pleistocene mammals from the Ushbulak site in East Kazakhstan (4). Shading shows the general trend of changes in the nanoporosity of fossil bones during their fossilization
Таблица 2. Валовый химический состав костей, мас. %
Table 2. Gross chemical composition of bones, wt. %
|
№ обр. / Sample No. |
SiO 2 |
TiO 2 |
Al2O3 |
Fe 2 O 3 |
Y 2 O 3 |
MnO |
ZnO |
MgO |
CaO |
BaO |
SrO |
Na2O |
K 2 O |
P 2 O 5 |
SO 3 (Сl) |
As2O3 |
UO 2 |
Ca/P ат |
|
Врб-1 |
0.31 |
н. о. |
0.17 |
1.12 |
н. о. |
0.67 |
н. о. |
0.58 |
60.97 |
н. о. |
0.23 |
0.67 |
0.05 |
34.77 |
0.46 |
н. о. |
н. о. |
2.22 |
|
Миоцен Miocene |
0.31 |
н. о. |
0.17 |
1.12 |
н. о. |
0.67 |
н. о. |
0.58 |
60.97 |
н. о. |
0.23 |
0.67 |
0.05 |
34.77 |
0.46 |
н. о. |
н. о. |
2.22 |
|
Врб-2 |
2.61 |
0.06 |
1.11 |
0.5 |
» |
н. о. |
н. о. |
1.13 |
57.39 |
» |
0.25 |
0.61 |
0.14 |
35.06 |
1.14 |
н. о. |
» |
2.07 |
|
Врб-3 |
0.19 |
н. о. |
н. о. |
0.5 |
» |
» |
» |
0.81 |
58.65 |
» |
0.2 |
0.81 |
0.04 |
37.2 |
2.03 |
» |
» |
2.0 |
|
Врб-4 |
0.29 |
» |
0.17 |
0.14 |
0.07 |
0.08 |
» |
0.68 |
59.93 |
» |
0.22 |
0.46 |
0.04 |
36.75 |
1.02 |
» |
0.15 |
2.07 |
|
Плиоцен Pliocene |
1.03 ± |
0.02 ± |
0.43 ± |
0.38 ± |
0.02 ± |
0.03 ± |
0.87 ± |
58.66 ± |
0.22 ± |
0.63 ± |
0.07± |
36.34 ± |
1.4 ± |
0.05± |
2.05 ± |
|||
|
1.37 |
0.03 |
0.6 |
0.21 |
0.04 |
0.05 |
0 |
0.23 |
1.27 |
0 |
0.03 |
0.18 |
0.06 |
1.13 |
0.55 |
0 |
0.09 |
0.04 |
|
|
(133) |
(150) |
(140) |
(55 ) |
(200) |
(167) |
(26) |
(2) |
(133) |
(29) |
(86) |
(3) |
(39) |
(180) |
(2) |
||||
|
Врб-5 |
0.63 |
н. о. |
0.25 |
0.09 |
н. о. |
0.04 |
н. о. |
0.75 |
58.4 |
0.27 |
0.38 |
0.51 |
0.05 |
37.93 |
0.69 |
н. о. |
0.01 |
1.95 |
|
Врб-6 |
2.36 |
» |
0.35 |
8.39 |
» |
1.51 |
» |
0.7 |
51.78 |
0.63 |
0.23 |
0.36 |
0.05 |
33.19 |
0.45 |
» |
н. о. |
1.98 |
|
Врб-7 |
1.57 |
» |
0.33 |
7.98 |
0.03 |
0.5 |
» |
0.67 |
52.04 |
0.23 |
0.35 |
0.62 |
0.03 |
34.38 |
1.23 |
» |
0.04 |
1.92 |
|
Тит-1 |
0.72 |
» |
0.22 |
5.98 |
0.12 |
0.2 |
» |
0.87 |
55.72 |
н. о. |
0.34 |
0.35 |
н. о. |
34.76 |
0.71 |
0.01 |
н. о. |
2.03 |
|
Тит-2 |
2.13 |
» |
0.4 |
5.01 |
н. о. |
0.59 |
» |
0.98 |
54.15 |
» |
0.26 |
0.73 |
0.05 |
34.53 |
1.16 |
0.01 |
» |
1.99 |
|
Тит-3 |
2.56 |
» |
0.35 |
2.67 |
0.02 |
0.8 |
» |
н. о. |
56.11 |
» |
0.31 |
0.52 |
0.04 |
36.11 |
0.51 |
н. о. |
» |
1.97 |
|
Тавр-1 |
1.89 |
» |
1.06 |
0.41 |
н. о. |
0.48 |
0.45 |
0.85 |
55.07 |
» |
0.26 |
0.7 |
0.09 |
37.61 |
н. о. (0.54) |
» |
» |
1.86 |
|
Тавр-2 |
0.5 |
» |
0.46 |
0.5 |
» |
0.06 |
0.42 |
0.63 |
56.78 |
» |
0.25 |
0.61 |
0.02 |
38.58 |
(0.»59) |
н. о. |
н. о. |
1.87 |
|
SiO 2 |
TiO 2 |
Al2O3 |
Fe 2 O 3 |
Y 2 O 3 |
MnO |
ZnO |
MgO |
CaO |
BaO |
SrO |
Na2O |
K 2 O |
P 2 O 5 |
SO3 (Сl) |
As2O3 |
UO 2 |
Ca/P ат |
|
|
Палеоплейстоцен |
0.59± |
|||||||||||||||||
|
(гелазий) |
1.55± |
0 |
0.43± |
3.88± |
0.02± |
0.52± |
0.11± |
0.68± |
55.01± |
0.14± |
0.3± |
0.55± |
0.04± |
35.89± |
0.46 |
0.003± |
0.01± |
1.95± |
|
Paleopleistocene |
0.82 |
0.27 |
3.42 |
0.04 |
0.48 |
0.2 |
0.3 |
2.28 |
0.23 |
0.05 |
0.14 |
0.03 |
1.97 |
(0.14± |
0.01 |
0.01 |
0.06 |
|
|
(Gelasian) |
0.26) |
|||||||||||||||||
|
Врб-8 |
0.63 |
н. о. |
0.2 |
5.57 |
н. о. |
0.73 |
н. о. |
0.69 |
54.29 |
н. о. |
0.18 |
0.65 |
0.04 |
35.5 |
1.52 |
н. о. |
н. о. |
1.94 |
|
Врб-9 |
0.21 |
» |
н. о. |
0.71 |
н. о. |
0.14 |
» |
0.73 |
61.18 |
» |
0.25 |
0.62 |
н. о. |
35.84 |
0.32 |
» |
» |
2.16 |
|
Врб-10 |
1.22 |
» |
0.2 |
6.79 |
0.02 |
0.16 |
» |
0.56 |
54.53 |
» |
0.16 |
0.46 |
0.02 |
35.34 |
0.56 |
» |
» |
1.96 |
|
Средний |
0.69± |
0.13 ± |
4.36 ± |
0.007 ± |
0.34 ± |
0.66 ± |
56.67 ± |
0.02± |
0.58 ± |
0.02± |
35.56 ± |
0.8± |
2.02± 0.12 |
|||||
|
плейстоцен |
0.51 |
0 |
0.11 |
3.22 |
0.011 |
0.33 |
0 |
0.09 |
3.91 |
0 |
0.05 |
0.1 |
0.02 |
0.25 |
0.63 |
0 |
0 |
|
|
Middle Pleistocene |
(74) |
(85) |
(74) |
(157) |
(97) |
(14) |
(7) |
(25) |
(17) |
(74) |
(1) |
(79) |
||||||
|
Тит-4 |
1.5 |
н. о. |
0.33 |
0.14 |
н. о. |
0.13 |
0.04 |
0.73 |
58.86 |
н. о. |
0.14 |
0.52 |
0.08 |
38.53 |
0.57 |
н. о. |
н. о. |
2.02 |
|
Тит-5 |
2.02 |
» |
0.69 |
0.26 |
» |
0.06 |
0.02 |
0.88 |
56.61 |
» |
0.24 |
0.45 |
0.15 |
38.62 |
0.63 |
» |
» |
1.89 |
|
Тит-6 |
2.77 |
» |
0.95 |
0.2 |
» |
н. о. |
0.02 |
0.79 |
55.26 |
» |
0.12 |
0.49 |
0.16 |
40.26 |
0.56 |
» |
» |
1.81 |
|
Тит-7 |
0.71 |
» |
0.3 |
0.25 |
» |
» |
н. о. |
0.97 |
59.65 |
» |
0.24 |
0.93 |
0.18 |
36.77 |
1.12 |
» |
» |
2.12 |
|
Голоцен Holocene |
1.75 ± |
0.57 ± |
0.21 ± |
0.05 ± |
0.02 ± |
0.84 ± 0.1 (12) |
57.95 |
0.18± |
0.6 ± |
0.14 ± |
38.54 |
0.72± |
1.96± 0.14 (7) |
|||||
|
0.87 (50) |
0 |
0.31 (54) |
0.06 (29) |
0 |
0.06 (120) |
0.02 (100) |
± 2.02 (3) |
0 |
0.06 (33) |
0.22 (37) |
0.04 (28) |
±1.43 (50) |
0.27 (37) |
0 |
0 |
Примечание: н. о. —не обнаружено. Note: н. о. — not detected.
вых примесей в костях ископаемых млекопитающих с территорий Печорского Приуралья, Пермской области (стоянка Заозерье), Северо-Восточного Казахстана (стоянка Ушбулак). При этом атомный модуль Ca/P в верблюжьих костях на 3—10 % выше, чем в сопоставимых по возрасту костях исследованных нами ранее плейстоценовых животных. Выявленная особенность говорит об аномально высокой степени кальцитиза-ции верблюжьих костей, возрастающей в направлении от голоценовых костей к позднемиоценовым (рис. 8). По этому показателю верблюжьи кости уступают только ископаемым костям, претерпевшим пещерную фос-силизацию. В последних значение модуля Ca/Pат достигает 2.35 (Силаев и др., 2020).
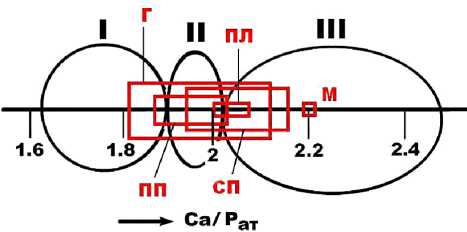
Рис. 8. Вариации значений карбонатапатитового модуля в костях ископаемых верблюдов: позднемиоценовых (М); плиоценовых (ПЛ); плейстоценовых, гелазийский век (ПП); плейстоценовых, калабрийский век (СП); голоценовых (Г). I, II, III — области ископаемых костей: соответственно некальцитизированные, слабокальцитизированные и сильно кальцитизированные
Fig. 8 . Variations in the values of the carbonate-apatite modulus in the bones of fossil camels: Late Miocene (M); Pliocene (PL); Pleistocene, Gelasian Age (PP); Pleistocene, Calabrian Age (SP); Holocene (G). I, II, III — fossil areas bones: respectively non-calcified, weakly calcified and highly calcified
Состав неапатитовых примесей в верблюжьих костях определялся рентгенофазовым и рентгеноспектральным микрозондовым методами. В результате проведенного анализа во всех исследованных образцах выявлены прежде всего минералы, иллювииро-ванные из вмещающих кости грунтов: кварц, альбит Na[Al1—1.01Si3O8]; ортоклаз K0.97—1.02[Al0.99—1Si3O8]; циркон Zr0.98—1.02[SiO4]; ильменит (Fe0.96—1.04Mn0—0.03)Ti1— 1.04O3; гидрослюды и хлориты состава соответственно (K0.44—0.97Ca0—0.03)0.43—0.97(Al1.39—1.56Fe0.18—0.55Mg0— 0.18Ti0—0.06)1.98—2.01[AlSi3O10](OH)1.33—1.47 и (Mg4.1—5Fe0— 0.09)5Al0.91—1.01[AlSi3O10](OH)7.9—8.1. Кроме этих примесей в верблюжьих костях выявлен ряд эпигенетических по отношению к биоапатиту минералов, образовавшихся аутигенно в ходе фоссилизации. Типичные энергодисперсионные спектры таких минералов приведены на рис. 9. В состав эпигенетических минералов входят: кальцит Ca[CO3]; поликомпонентный родохрозит состава (Mn0.54—0.94Ca0.03—0.27Fe0—0.33Ba0—0.04) [CO3]; барит (Ba0.72—1Sr0—0.14Ca0—0.36)[SO4]; барито-ан-гидрит (Ba0.41—0.54Ca0.32—0.52Sr0.08—0.14)1—1.01[SO4]; пирит (Fe0.92—1Co0—0.02Cu0—0.03)S2; гётит (Fe0.69—1Mn0— 0.31Cr0—0.03Al0—0.05)O(OH); манганит (Mn0.61—0.91Fe0.03— 0.35Ni0—0.05Ba0—0.03)O(OH). В отношении пирита следует добавить, что он представлен, как правило, фрамбоидальными формами субмикронных размеров, что прямо указывает на активное участие бактерий в фоссилизации костей.
Полученные данные по фазовому составу примесей в исследованных костях дают возможность рассчитать нормативный минеральный состав последних. Полученные результаты (табл. 3) показывают, что в хронологической последовательности от голоценовых верблюдов к позднемиоценовым доля биоапатита сокращается в среднем почти на 8 мол. %, но на 8.6 мол. % возрастает содержание карбонатов. Таким образом, выявляется факт прямой связи степени эпигенетической карбонатизации костей ископаемых верблюдов именно с их геологическим возрастом. Степень эпигенетического оксигидроксидного оже-лезнения костей тоже имеет общую тенденцию увеличиваться с возрастом костей, но при этом аномально проявляется в гелазийских и калабрийских плейстоценовых образцах. Последнее связано, возможно, с особенностями состава соответствующих костеносных грунтов.
Особое значение имеет анализ нормативно-минерального состава иллювиированной в кости терри-
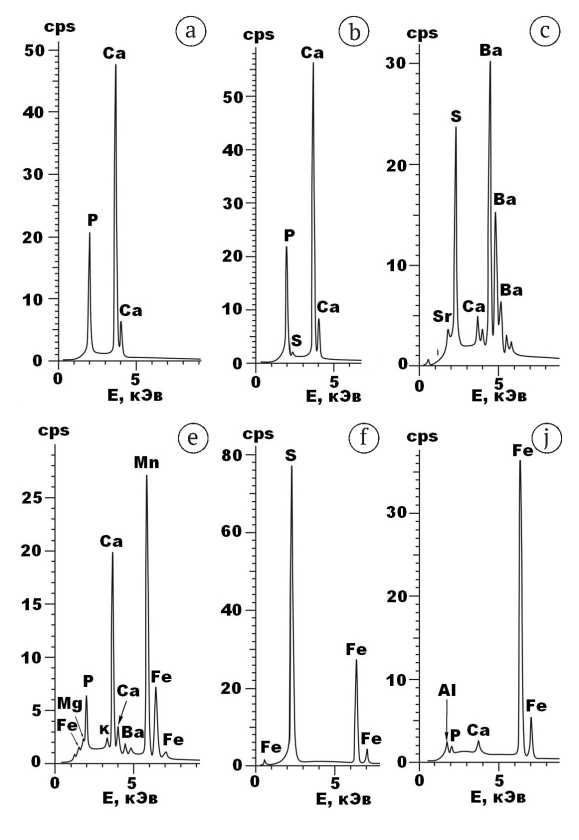
Рис. 9 . Типичные ЭД-спектры биоапатита и аутигенных минералов-примесей в исследованных костях ископаемых верблюдов: a, b — биоапатит (Тит-4, Врб-5), c — барит (Врб-7, Тит-2), d — родохрозит (Врб-3), e — пирит (Тит-6), f — гётит (Врб-6)
Fig. 9. Typical ED spectra of bioapatite and authigenic min-erals-impurities in the studied bones of fossil camels: a, b — bioapatite (Tit-4, Vrb-5), c — barite (Vrb-7, Tit-2), d — rhodochrosite (Vrb-3), e — pyrite (Tit-6), f — goethite (Vrb-6)
Таблица 3. Нормативно-минеральный состав костей, мол. % генной примеси. Анализ показал, что по литолого-минеральному составу терригенная примесь в верблюжьих костях варьируется от супесей к суглинкам и от слюдисто-хлорит-кварцевых до кварц-слюдисто-хлоритовых и хлоритовых (рис. 10). При этом наиболее глинистая и наименее кварцсодержащая примесь содержится в костях позднемиоценовых верблюдов,
а более песчаная и кварцевая — в костях плиоценовых верблюдов. В более молодых костях иллювиированная примесь изменяется от слюдисто-кварц-хлоритовой у плейстоценовых верблюдов до кварц-слюдисто-хлоритовой у голоценовых. В целом по составу терригенной примеси исследованные кости верблюдов очень своеобразны, отличаясь от ранее изученных ископае-
Table 3. Normative-mineral composition of bones, mol. %
|
№ обр. Sample No. |
Апатит Apatite |
Карбонаты Carbonates |
Кварц + полевые шпаты Quarts + Feldspar |
Гидрослюда Hydromica |
Хлориты Chlorites |
Гётит Goethite |
|
Врб-1 |
79.72 |
16.0 |
0 |
1.1 |
1.76 |
1.42 |
|
поздний миоцен Late Miocene |
79.72 |
16.0 |
0 |
1.1 |
1.76 |
1.42 |
|
Врб-2 |
77.66 |
11.6 |
5.8 |
1.24 |
3.33 |
0.37 |
|
Врб-3 |
88.49 |
10.91 |
0.2 |
0 |
0 |
0.4 |
|
Врб-4 |
84.8 |
12.65 |
0 |
0.28 |
2.09 |
0.18 |
|
плиоцен |
83.65 ± 5.5 |
11.72 ± 0.88 |
2 ± 3.29 |
0.51 ± 0.65 |
1.81 ± 1.68 |
0.32 ± 0.12 |
|
Pliocene |
(7) |
(8) |
(165) |
(127) |
(93) |
(37) |
|
Врб-5 |
87.84 |
9.29 |
0 |
0.47 |
2.3 |
0.1 |
|
Врб-6 |
78.69 |
9.07 |
1.6 |
0.48 |
2.21 |
7.95 |
|
Врб-7 |
82.22 |
7.68 |
0.9 |
0.27 |
2.14 |
6.79 |
|
Тит-1 |
85.45 |
7.06 |
0 |
0 |
2.69 |
4.8 |
|
Тит-2 |
81.32 |
9.69 |
1.09 |
0.41 |
3.06 |
4.43 |
|
Тит-3 |
84.79 |
9.53 |
2.5 |
0.39 |
0 |
2.79 |
|
Тавр-1 |
88.62 |
6.2 |
0.77 |
0.83 |
2.83 |
0.75 |
|
Тавр-2 |
90.76 |
6.65 |
0 |
0.17 |
1.98 |
0.44 |
|
плейстоцен (гелазий) Pleistocene (Gelasian) |
84.96 ± 4.06 |
8.15 ± 1.41 |
0.86 ± 0.89 |
0.38 ± 0.24 |
2.15 ± 0.94 |
3.51 ± 2.98 |
|
Врб-8 |
83.94 |
8.46 |
0 |
0.4 |
2.18 |
5.02 |
|
Врб-9 |
83.84 |
15.48 |
0.22 |
0 |
0 |
0.68 |
|
Врб-10 |
83.05 |
8.91 |
0.67 |
0.17 |
1.75 |
5.45 |
|
плейстоцен (калабрий) |
83.61 ± |
10.95 ± 3.93 |
0.3 ± 0.34 |
0.19 ± 0.2 |
1.31 ± 1.15 |
3.72 ± 2.64 |
|
Pleistocene (Calabrian) |
0.49 (1) |
936) |
(113) |
(105) |
(88) |
(71) |
|
Тит-4 |
87.58 |
8.75 |
0.54 |
0.72 |
2.19 |
0.22 |
|
Тит-5 |
88.75 |
6.27 |
0.67 |
1.37 |
2.69 |
0.25 |
|
Тит-6 |
92.09 |
2.43 |
1.47 |
1.45 |
2.41 |
0.15 |
|
Тит-7 |
83.24 |
12.06 |
0 |
1.6 |
2.91 |
0.19 |
|
голоцен |
87.91 ± |
7.38 ± 4.06 |
0.67 ± 0.61 |
1.28 ± 0.39 |
2.55 ± 0.32 |
0.2 ± 0.04 |
|
Holocene |
3.66 (4) |
(55) |
(91) |
(3) |
(12) |
(20) |
Рис. 10 . Треугольник нормативно-минерального состава иллюви-ированной в кости ископаемых верблюдов терригенной примеси. Верблюды: М — позднемиоценовые; ПЛ — плиоценовые; ПГ — плейстоценовые, гелазийский век; ПК — плейстоценовые, калабрийский век; Т — плейстоценовые, калабрийский век, пещера Таврида; Г — голоценовые. ЗОЗ — состав иллювиированной примеси в костях лошадей на стоянке Заозерье. Черные квадраты — средний состав литотипов. Области состава на треугольнике: 1—3 — пески соответственно кварцевые, хлорит-слюдисто-кварцевые и слюдисто-хлорит-кварцевые; 4, 5 — супеси соответственно хлорит-кварц-слюдистые и слюдисто-кварц-хлоритовые; 6—9 — суглинки и глины соответственно слюдистые, кварц-хлорит-слюдистые, кварц-слюдисто-хлоритовые и хлоритовые
Fig. 10. Triangle of normative-mineral composition of terrigenous admixture illuviated in the bones of fossil camels.
Camels: M — Late Miocene; PL — Pliocene; PG — Pleistocene, Gelasian age; PC — Pleistocene, Calabrian age; T — Pleistocene, Calabrian age,
КВАРЦ + ПОЛЕВЫЕ ШПАТЫ
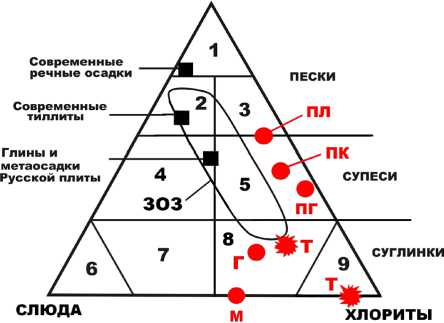
Taurida cave; G — Holocene. ZOS is the composition of the illuviated admixture in the bones of horses at the Zaozerye site. Black squares are average composition of lithotypes. Areas of composition on the triangle: 1—3 — quartz, chlorite-mica-quartz, and micaceous-chlorite-quartz sands, respectively; 4, 5 — chlorite-quartz-micaceous and micaceous-quartz-chlorite sandy loams, respectively; 6—9 — loams and clays, respectively, micaceous, quartz-chlorite-micaceous, quartz-micaceous-chlorite and chlorite
Таблица 4. Содержание микроэлементов в исследованных костях верблюдов, г/т
Table 4. The content of microelements in the studied camel bones, ppm
|
Элементы Elements |
Плейстоценовые / Pleistocene |
Голоценовые / Holocene |
|||||
|
Тит-2 |
Тит-3 |
Врб-6 |
Тит-4 |
Тит-5 |
Тит-6 |
Тит-7 |
|
|
Be |
2.1 |
0.6 |
1.2 |
0.007 |
0.01 |
0.015 |
0.004 |
|
Zn |
25 |
22 |
25 |
50 |
50 |
60 |
50 |
|
As |
68 |
12.8 |
18.3 |
4.4 |
4.7 |
3.8 |
3.3 |
|
Se |
2.2 |
0.54 |
1.01 |
0.41 |
0.37 |
0.38 |
0.33 |
|
Rb |
0.028 |
0.005 |
0.05 |
0.04 |
0.08 |
0.12 |
0.023 |
|
Mo |
4.5 |
0.6 |
3.7 |
1.5 |
4.3 |
2 |
3.7 |
|
Ag |
0.0048 |
0.0025 |
0.0119 |
0.016 |
0.0036 |
0.0053 |
0.019 |
|
Cd |
0.24 |
0.09 |
0.024 |
0.0002 |
0.0002 |
0.0002 |
0.0002 |
|
Pb |
1.6 |
1.4 |
5 |
0.35 |
0.9 |
0.6 |
0.2 |
|
Bi |
0.0005 |
0.00059 |
0.0032 |
0.0025 |
0.0049 |
0.0022 |
0.0005 |
|
Th |
0.016 |
0.024 |
0.06 |
0.01 |
0.021 |
0.026 |
0.006 |
|
Эссенциалы (Э) Essentials |
103.689 |
38.062 |
54.736 |
56.736 |
60.39 |
66.949 |
57.583 |
|
Li |
1.2 |
1.7 |
1.5 |
10 |
8 |
6 |
5 |
|
Ti |
700 |
800 |
800 |
700 |
700 |
800 |
700 |
|
V |
60 |
12 |
23 |
15 |
7 |
23 |
7 |
|
Cr |
80 |
6 |
9 |
1.7 |
0.9 |
1.9 |
1.7 |
|
Mn |
500 |
1000 |
1600 |
110 |
60 |
21 |
14 |
|
Co |
2.5 |
9 |
8 |
1.3 |
1.2 |
1.2 |
1 |
|
Ni |
34 |
38 |
40 |
32 |
31 |
33 |
29 |
|
Cu |
7 |
1.9 |
3 |
2.3 |
1 |
3.9 |
2.9 |
|
Ga |
2.2 |
2.1 |
2.4 |
1.8 |
1.8 |
2 |
1.6 |
|
Sr |
1200 |
1000 |
1200 |
500 |
700 |
600 |
600 |
|
Y |
190 |
21 |
70 |
0.19 |
0.3 |
0.3 |
0.13 |
|
Zr |
22.1 |
2.2 |
7 |
0.21 |
0.19 |
0.3 |
0.15 |
|
Sn |
0.003 |
0.003 |
0.003 |
0.003 |
0.003 |
0.003 |
0.003 |
|
Sb |
1 |
0.66 |
0.71 |
0.04 |
0.04 |
0.11 |
0.012 |
|
Cs |
0.0003 |
0.0003 |
0.0009 |
0.0003 |
0.,0061 |
0.008 |
0.0003 |
|
Ba |
260 |
1700 |
350 |
90 |
130 |
200 |
90 |
|
Hf |
0.18 |
0.031 |
0.09 |
0.003 |
0.004 |
0.007 |
0.00356 |
|
U |
100 |
130 |
80 |
0.34 |
5 |
0.44 |
0.21 |
|
Физиогенно-активные (ФА) |
3160.183 |
4724.594 |
4194.704 |
1464.886 |
1646.437 |
1693.168 |
1454.709 |
|
Physiogenic-active |
|||||||
|
B |
1.3 |
5 |
23 |
7 |
23 |
0.09 |
9 |
|
Sc |
5 |
0.5 |
2.1 |
0.25 |
0.25 |
0.31 |
0.21 |
|
Ge |
0.18 |
0.035 |
0.078 |
0.004 |
0.004 |
0.004 |
0.004 |
|
Nb |
0.05 |
0.0001 |
0.039 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
|
Te |
0.7 |
0.28 |
0.4 |
0.064 |
0.039 |
0.028 |
0.017 |
|
La |
27 |
5 |
13 |
0.08 |
0.17 |
0.19 |
0.04 |
|
Ce |
2.1 |
2.1 |
6 |
0.12 |
0.51 |
0.35 |
0.058 |
|
Pr |
5.1 |
1 |
2.6 |
0.019 |
0.038 |
0.042 |
0.009 |
|
Nd |
26 |
4.6 |
12 |
0.069 |
0.16 |
0.19 |
0.045 |
|
Sm |
6 |
1 |
2.8 |
0.017 |
0.035 |
0.036 |
0.008 |
Окончание таблицы 4 / End of Table 4
Микроэлементы и геохимические критерии фоссилизации
В составе исследуемых костных остатков обнаружен 51 микроэлемент (табл. 4), в том числе 11 элемен-тов-эссенциалов, 18 физиологически активных эле-
ментов и 22 элемента-антибионта. Элементы первой группы в костях наследуются от живого организма, концентрируясь в костном коллагене. Вторая группа объединяет элементы, которые могут быть по происхождению как унаследованными от организма, так и обусловленными фоссилизацией. Третья группа включает только элементы, которые по происхождению являются нацело следствием загрязнения костей в среде захоронения.
Суммарная концентрация микроэлементов в исследованных верблюжьих костях изменяется в диапазоне от 1500 до 5000 г/т, возрастая в 2—3 раза по мере их удревнения. Например, в проанализированных ко-
стях плейстоценовых и голоценовых верблюдов суммарное содержание микроэлементов колеблется в пределах соответственно 1520—1770 и 3400—4800 г/т. В сравнительном отношении верблюжьи кости по суммарному содержанию микроэлементов могут быть сопоставлены только с костями млекопитающих со стоянки Ушбулак. Все другие изученные нами костные остатки, например плейстоценовых лошадей со сто-
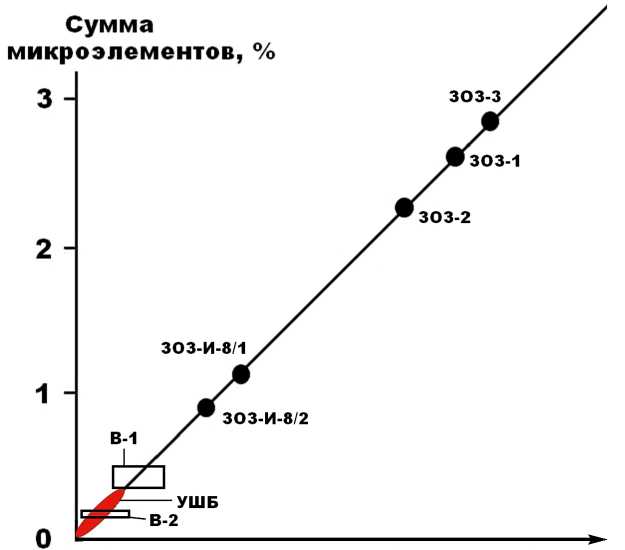
Продолжительность фоссилизации
Рис. 11 . Суммарная концентрация микроэлементов в костях ископаемых верблюдов плиоцен-плейстоцено-вого (В-1) и голоценового (В-2) возраста в сопоставлении с костями плейстоценовых лошадей со стоянки Заозерье (ЗОЗ) и плейстоценовых млекопитающих со стоянки Ушбулак (УШБ)
Fig. 11 . The total concentration of trace elements in the bones of fossil camels of the Pliocene-Pleistocene (B-1) and Holocene (B-2) age in comparison with the bones of Pleistocene horses from the Zaozerye site (ZOZ) and Pleistocene mammals from the Ushbulak site (USHB)
янки Заозерной, характеризовались многократно большим обогащением микроэлементами.
Ранее было показано, что пропорция между элементами эссенциалами и антибионтами хорошо отражает степень фоссилизации захороненных костей. В рассматриваемом случае отношение групповых концентраций ЭС/АБ изменяется от 15.35 ± 20.36 (многократное преобладание элементов-эссенциалов) в голоценовых костях до 0.93 ± 0.41 (преобладание эле-ментов-антибионтов) в костях плейстоценовых верблюдов. Эту закономерность отражает и отношение содержаний эссенциального цинка и физиогенно-ак-тивной меди, которое в хронологической последовательности изменяется от 26.09 ± 11.16 до 7.83 ± 4.93.
Дополнительным критерием степени фоссилиза-ции и относительного возраста ископаемых костей может служить суммарная концентрация лантаноидов, накапливающихся в костях именно в результате их взаимодействия с вмещающими грунтами. В рассматриваемом случае эта концентрация в хронологической последовательности возрастает практически на два порядка — от 0.71 ± 0.44 в голоценовых костях до 69.23 ± 57.96 г/т в плейстоценовых.
Костный биоапатит
Кости млекопитающих представляют собой органоминеральный композит, состоящий из органической матрицы и производимого ею минерального вещества — биоапатита. Известно, что в ходе фоссили-зации погребенных костей биоапатит подвергается изменениям, выражающимся прежде всего в увеличении степени кристалличности, которая обычно коррелируется с геологическим возрастом костей.
На рентгеновских дифрактограммах биоапатита исследованных костных образцов (рис. 12, 13) наблюдаются основные отражения, характерные для карбо-натапатита В-типа (Å): 3.41—3.46; 2.78—2.82 (121);
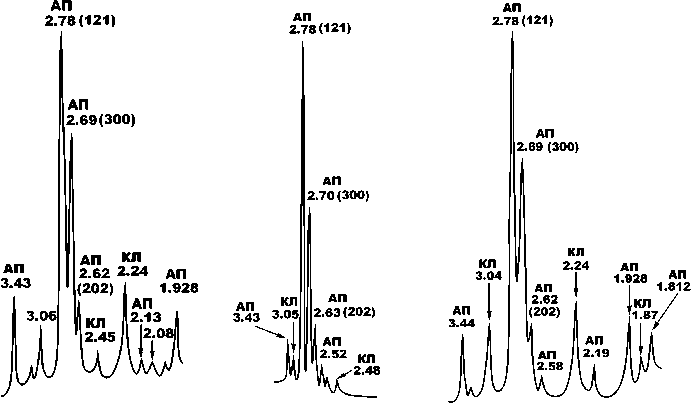
Рис. 12. Типичные рентгеновские дифрактограммы биоапатита в костях голоценовых верблюдов. Минералы: АП — биоапатит, КЛ — кальцит. Образцы слева направо: Тит-5 (III век до н. э. — III н. э.), Тит-4 (V век н. э.), Тит-7 (XIV век н. э.)
Fig. 12 . Typical X-ray diffraction patterns of bioapatite in the bones of Holocene camels. Minerals: AP — bioapatite, CL — calcite. Samples from left to right: Tit-5 (III century BC — III AD), Tit-4 (V century AD), Tit-7 (XIV century AD)
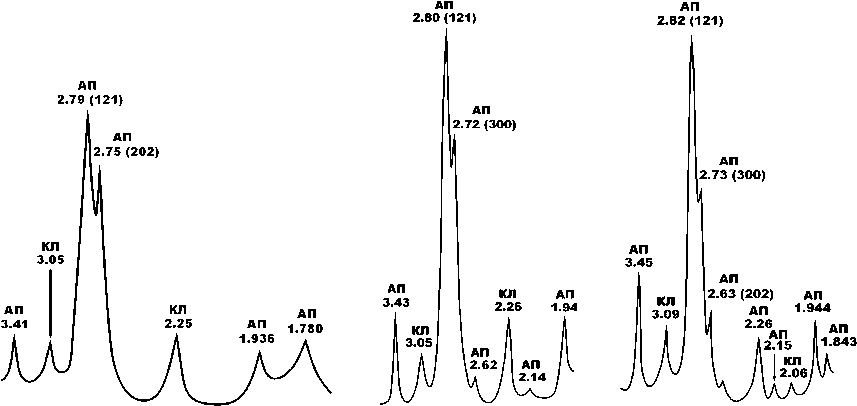
Рис. 13 . Типичные рентгеновские дифрактограммы биоапатита в костях плиоцен-плейстоценовых верблюдов. Образцы слева направо: Врб-3 (плиоцен), Тит-2 (плейстоцен, гелазий), Тавр-1 (плейстоцен, калабрий)
Fig. 13 . Typical x-ray diffraction patterns of bioapatite in bones Pliocene-Pleistocene camels. Samples from left to right: Vrb-3 (Pliocene), Tit-2 (Pleistocene, Gelasian), Tavr-1 (Pleistocene, Calabrian)
2.69—2.75 (300); 2.62—2.63 (202); 1.928—1.944; 1.843; 1.812; 1,780. Кроме апатитовых пиков на рентгенограммах регистрируется серия отражений, отвечающих кальциту: 3.05—3.09; 2.45—2.48; 2.24—2.27; 2.06— 2.08; 1.87. Все пики биоапатита демонстрируют значительное уширение (FWHM), а наиболее интенсивный из них — (121) — подвергается расщеплению с появлением двух дополнительных пиков малой интенсивности — (300) и (202). Именно эти эффекты мы используем для оценки степени кристалличности минерала. Для голоценовых костей FWHM составляет 0.51 ± 0.06. С удревнением костей эта величина последовательно сокращается до 0.42 ± 0.08 в плейстоцен-калабрийских, 0.35 ± 0.06 — в плейстоцен-гелазийских, 0.29 ± 0.04 — в плиоценовых и позднемиоценовых. Таким образом, выявляется устойчивая тенденция к значительному сокращению ширины рентгеновского пика вследствие
возрастания степени кристалличности костного биоапатита. Степень расщепления рентгеновского пика (121) мы оцениваем по отношению интенсивностей отражений I(300)/I(121). Для биоапатита в калабрийско-голоценовых костях величина этого отношения оценивается в 0.15 ± 0.09, в гелазийских костях — 0.32 ± 0.1, в плиоценовых — 0.27 ± 0.08, в позднемиоценовых не опускается ниже 0.4. Из приведенных данных следует, что в наплавлении от наиболее молодых костей к наиболее древним степень расщепления пика (121) последовательно возрастает, отражая, как и в случае с FWHM, сильную хронологическую тенденцию увеличения степени кристалличности.
В ИК-спектрах, полученных от верблюжьих костей (рис. 14, 15) отчетливо регистрируются основные полосы ν3- и ν4-поглощения на РО4-группах, а также полосы ν2—ν4-поглощения на группах СО3, изоморфно
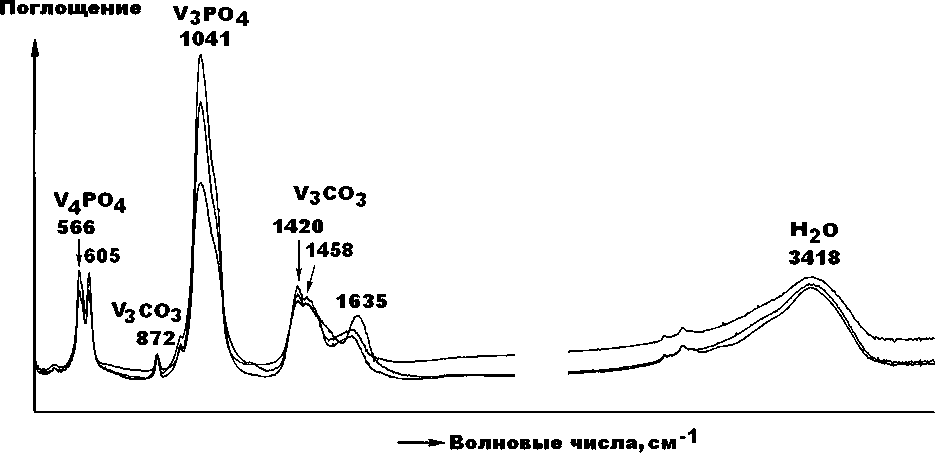
Рис. 14 . Спектры ИК-поглощения в относительно сильно карбонизированном биоапатите голоценовых верблюдов (обр. Тит-4, 5, 6)
Fig. 14 . IR absorption spectra in relatively highly carbonized bioapatite from Holocene camels (samples Tit-4, 5, 6)
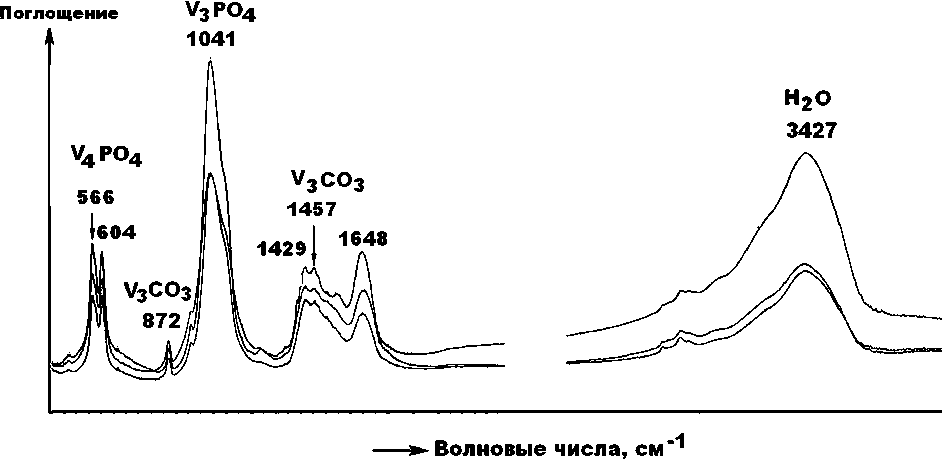
Рис. 15 . Спектры ИК-поглощения в относительно слабокарбонизированном биоапатите плиоцен-плейстоценовых верблюдов (обр. Врб-4; Тит-1, 2)
Fig. 15 . IR absorption spectra in relatively weakly carbonized bioapatite from Pliocene-Pleistocene camels (sample Vrb-4; Tit-1, 2)
замещающих фосфатные анионы. Признаков поглощения на группах СО3, замещающих ОН-группы (валентные анионы), не обнаружено. Таким образом, результаты ИК-спектроскопии тоже свидетельствуют о том, что биоапатит в костных остатках верблюдов нацело представлен гидроксилкарбонатапатитом В-типа.
В качестве критерия структурной карбонизации биоапатита мы использовали отношение интенсивностей поглощения в полосах при 1420 см–1 [ν3(СО3)] и при 1041 см–1 и [ν3(РО4]. Значения этого критерия для относительно слабокарбонизированного биоапатита в костях плиоцен-плейстоценовых верблюдов составляет 0.31 ± 0.06, а для относительно сильно карбонизированного биоапатита в костях голоценовых верблюдов — 0.35 ± 0.05. Из приведенных данных следует, что в направлении от молодых костей к более древним степень структурной карбонизации биоапатита сокращается, что согласуется с вышеотмеченной тенденцией к росту значений рентгеновского индекса его кристалличности.
Ранее подобный результат был получен по костям млекопитающих на стоянке Ушбулак (Силаев и др., 2022). Следует отметить, что полученные ИКС-данные 16
для верблюдов хорошо коррелируются с аналогичными данными для ископаемых костей, захороненных в открытых грунтах. Для костей, претерпевших фос-силизацию в пещерных условиях, ИКС-критерий карбонизации биоапатита достигает больших значений — 0.55 ± 0.22 (Силаев и др., 2020).
Химический состав биоапатита в исследуемых костях имеет сравнительно простой состав, характеризуясь структурной примесью Sr и Mn в катионной подрешетке и Si, S, C — в анионной. При этом наблюдаются некоторые хронологические вариации. В частности, биоапатит в голоценовых и плейстоцен-кала-брийских костях не содержит примесей в катионной подрешетке, меньше содержит примесей кремния и серы в анионной подрешетке, статистически является наиболее карбонизированным. Апатит в гела-зийско-миоценовых костях более примесный в обеих подрешетках и несколько менее карбонизированный. Таким образом, в биоапатите верблюжьих костей явно реализуется хронологическая тенденция усложнения состава, совпадающая с тенденцией сокращении степени карбонизации, как это было показано выше на примере рентгеноструктурных и ИК-спектроскопических свойств.
Таблица 5. Химический состав (мас. %) и эмпирические формулы биоапатита в костях ископаемых верблюдов Table 5. Chemical composition (wt %) and empirical formulas of bioapatite in the bones of fossil camels
|
№ обр. Sample No. |
CaO |
SrO |
MnO |
P 2 O 5 |
SO 3 |
SiO 2 |
Cl |
Ca/P ат |
Формулы Formulas |
|
Врб-1 |
|||||||||
|
1 |
61.23 |
н. о. |
н. о. |
38.77 |
н. о. |
н. о. |
н. о. |
2.0 |
Ca 10 [P 4.99 C 1.01 O 24 ](OH) 0.99 |
|
2 |
58.49 |
1.52 |
» |
38.86 |
» |
1.13 |
» |
1.91 |
(Ca 9.96 Sr 0.04 )[P 5.17 Si 0.18 C 0.65 O 24 ](OH) 1.17 |
|
3 |
59.74 |
н.о. |
» |
39.99 |
» |
н. о. |
0.27 |
1.89 |
Ca 10 [P 5.28 C 0.72 O 24 ]Cl 0.07 (OH) 0.99 |
|
4 |
60.13 |
» |
» |
39.87 |
» |
» |
н.о. |
1.91 |
Ca 10 [P 5.23 C 0.77 O 24 ](OH) 1.23 |
|
5 |
60.03 |
» |
» |
39.97 |
» |
» |
» |
2.07 |
Ca 10 [P 4.83 C 1.17 O 24 ](OH) 0.83 |
|
6 |
58.5 |
» |
» |
41.5 |
» |
» |
» |
1.79 |
Ca 10 [P 5.59 C 0.41 O 24 ](OH) 1.59 |
|
7 |
57.37 |
» |
» |
41.29 |
» |
1.08 |
0.26 |
1.76 |
Ca 10 [P 5.68 C 0.14 Si 0.18 O 24 ]Cl 0.07 (OH) 1.61 |
|
8 |
61.33 |
» |
» |
38.67 |
» |
н.о. |
н.о. |
2.01 |
Ca 10 [P 4.97 C 1.03 O 24 ](OH) 0.97 |
|
9 |
60.08 |
1.7 |
» |
38.22 |
» |
» |
» |
1.99 |
Ca 10 [P 5.02 C 0.98 O 24 ](OH) 1.02 |
|
10 |
59.66 |
н.о. |
» |
40.34 |
» |
» |
» |
1.87 |
Ca 10 [P 5.33 C 0.67 O 24 ](OH) 1.33 |
|
11 |
57.47 |
1.97 |
» |
40.56 |
» |
» |
» |
1.8 |
(Ca 9.82 Sr 0.12 )[P 5.46 C 0.54 O 24 ](OH) 1.46 |
|
12 |
58.63 |
н.о. |
1.18 |
38.82 |
» |
1.37 |
» |
1.92 |
(Ca 9.84 Mn 0.16 )[P 5.22 C 0.78 O 24 ](OH) 0.99 |
|
13 |
60.54 |
» |
0.62 |
37.55 |
» |
1.29 |
» |
2.04 |
(Ca 9.94 Mn 0.06 )[P 4.84 C 1.16 O 24 ](OH) 1.09 |
|
14 |
57.74 |
1.85 |
38.89 |
» |
1.52 |
» |
1.88 |
(Ca 10 Mn 0.00 )[P 5.32 C 0.68 O 24 ](OH) 1.32 |
|
|
поздний |
59.35 |
0.5± |
0.13± |
39.52 |
0 |
0.46 |
0.04 |
1.92 |
(Ca 9.82—10 Mn 0—0.16 Sr 0—0.18 ) 10 [P 4.83—5.68 |
|
Late Miocene |
±1.32 |
0.83 |
0.34 |
±1.16 |
±0.64 |
±0.1 |
±0.1 |
Si 0—0.18 C 0.14—1.17 O 24 ] Cl 0—0.07 (OH) 0.83—1.61 |
|
|
Врб-2 |
|||||||||
|
15 |
62.0 |
н.о. |
н.о. |
38.0 |
н.о. |
н.о. |
н.о. |
2.07 |
Ca 10 [P 4.83 C 1.17 O 24 ](OH) 0.83 |
|
16 |
59.92 |
» |
» |
40.08 |
» |
» |
» |
1.89 |
Ca 10 [P 5.28 C 0.72 O 24 ](OH) 1.28 |
|
17 |
61.81 |
» |
» |
36.62 |
1.57 |
» |
» |
2.14 |
Ca 10 [P 4.67 S 0.18 C 1.15 O 24 ](OH) 1.03 |
|
18 |
60.04 |
» |
» |
37.73 |
2.23 |
» |
» |
2.02 |
Ca 10 [P 4.96 S 0.26 C 0.78 O 24 ](OH) 1.03 |
|
19 |
59.0 |
» |
» |
39.37 |
1.63 |
» |
» |
1.9 |
Ca 10 [P 5.26 S 0.19 C 0.55 O 24 ](OH) 1.64 |
|
20 |
58.42 |
» |
» |
39.98 |
1.6 |
» |
» |
2.27 |
Ca 10 [P 4.4 S 0.19 C 0.41 O 24 ](OH) 1.78 |
|
21 |
59.06 |
» |
» |
39.13 |
1.81 |
» |
» |
1.91 |
Ca 10 [P 5.23 S 0.21 C 0.56 O 24 ](OH) 1.65 |
|
22 |
55.74 |
» |
» |
40.36 |
2.05 |
» |
» |
1.78 |
Ca 10 [P 5.61 S 0.25 C 0.14 O 24 ](OH) 2.11 |
|
23 |
57.86 |
» |
» |
40.09 |
2.05 |
» |
» |
1.83 |
Ca 10 [P 5.46 S 0.18 C 0.29 O 24 ](OH) 1.96 |
|
24 |
60.45 |
» |
» |
38.38 |
1.17 |
» |
» |
2.03 |
Ca 10 [P 4.93 S 0.13 C 0.94 O 24 ](OH) 1.19 |
|
25 |
58.73 |
» |
» |
38.81 |
2.46 |
» |
» |
1.92 |
Ca 10 [P 5.21 S 0.29 C 0.5 O 24 ](OH) 1.79 |
|
26 |
60.06 |
» |
» |
38.42 |
1.52 |
» |
» |
1.98 |
Ca 10 [P 5.05 S 0.18 C 0.77 O 24 ](OH) 1.41 |
|
27 |
57.26 |
» |
» |
39.47 |
2.78 |
» |
0.49 |
1.84 |
Ca 10 [P 5.44 S 0.34 C 0.22 O 24 ]Cl 0.19 (OH) 1.41 |
|
28 |
57.72 |
» |
» |
40.37 |
1.91 |
» |
н. о. |
1.81 |
Ca 10 [P 5.52 S 0.23 C 0.25 O 24 ](OH) 1.98 |
|
29 |
60.06 |
» |
» |
38.27 |
1.67 |
» |
» |
1.99 |
Ca 10 [P 5.03 S 0.19 C 0.78 O 24 ](OH) 1.41 |
|
30 |
60.3 |
» |
» |
37.53 |
2.17 |
» |
» |
2.04 |
Ca 10 [P 4.91 S 0.58 C 0.51 O 24 ](OH) 2.07 |
|
31 |
60.44 |
» |
» |
37.67 |
1.89 |
» |
» |
2.03 |
Ca 10 [P 4.92 S 0.22 C 0.86 O 24 ](OH) 1.36 |
|
32 |
60.47 |
» |
» |
37.11 |
2.42 |
» |
» |
2.07 |
Ca 10 [P 4.84 S 0.28 C 0.88 O 24 ](OH) 1.4 |
|
33 |
60.08 |
» |
» |
37.87 |
2.05 |
» |
» |
2.01 |
Ca 10 [P 4.97 S 0.24 C 0.79 O 24 ](OH) 1.04 |
|
34 |
60.38 |
» |
» |
37.41 |
2.21 |
» |
» |
2.04 |
Ca 10 [P 4.89 S 0.25 C 0.86 O 24 ](OH) 1.39 |
|
35 |
60.26 |
» |
» |
38.05 |
1.59 |
» |
» |
2.01 |
Ca 10 [P 4.98 S 0.18 C 0.84 O 24 ](OH) 1.34 |
|
36 |
60.89 |
» |
» |
37.34 |
1.77 |
» |
» |
2.07 |
Ca 10 [P 4.84 S 0.2 C 0.96 O 24 ](OH) 1.24 |
|
37 |
59.58 |
» |
» |
37.96 |
2.46 |
» |
» |
1.99 |
Ca 10 [P 5.02 S 0.29 C 0.69 O 24 ](OH) 1.6 |
|
38 |
60.13 |
» |
» |
37.08 |
2.79 |
» |
» |
2.06 |
Ca 10 [P 4.86 S 0.32 C 0.82 O 24 ](OH) 1.5 |
|
39 |
61.11 |
» |
» |
37.1 |
1.79 |
» |
» |
2.11 |
Ca 10 [P 4.74 S 0.2 C 1.01 O 24 ](OH) 1.19 |
|
40 |
61.27 |
» |
» |
36.37 |
2.36 |
» |
» |
2.14 |
Ca 10 [P 4.68 S 0.27 C 1.05 O 24 ](OH) 1.22 |
|
ВРБ-3 |
|||||||||
|
41 |
59.88 |
н.о. |
н.о. |
37.49 |
2.63 |
н.о. |
н.о. |
2.02 |
Ca 10 [P 4.94 S 0.31 C 0.75 O 24 ](OH) 1.56 |
|
42 |
59.12 |
» |
» |
36.12 |
4.4 |
» |
0.36 |
2.08 |
Ca 10 [P 4.82 S 0.52 C 0.66 O 24 ]Cl 0.1 (OH) 1.76 |
|
43 |
59.12 |
» |
» |
37.69 |
3.19 |
» |
н.о. |
1.99 |
Ca 10 [P 5.03 S 0.38 C 0.99 O 24 ](OH) 1.79 |
|
44 |
58.79 |
» |
» |
38.87 |
2.34 |
» |
» |
1.92 |
Ca 10 [P 5.22 S 0.28 C 0.5 O 24 ](OH) 1.79 |
|
45 |
57.73 |
» |
» |
39.93 |
2.34 |
» |
» |
1.83 |
Ca 10 [P 5.46 S 0.28 C 0.26 O 24 ](OH) 2.02 |
|
46 |
57.3 |
» |
» |
42.14 |
н.о. |
» |
0.56 |
1.72 |
Ca 10 [P 5.8 C 0.2 O 24 ]Cl 0.16 (OH) 1.64 |
|
47 |
60.16 |
» |
» |
37.7 |
2.14 |
» |
н.о. |
2.02 |
Ca 10 [P 4.94 S 0.25 C 0.81 O 24 ](OH) 0.44 |
|
48 |
58.04 |
» |
» |
39.23 |
2.73 |
» |
» |
1.88 |
Ca 10 [P 5.33 S 0.33 C 0.34 O 24 ](OH) 1.99 |
|
49 |
57.94 |
» |
» |
38.87 |
3.19 |
» |
» |
1.89 |
Ca 10 [P 5.29 S 0.38 C 0.33 O 24 ](OH) 2.02 |
|
50 |
58.68 |
» |
» |
37.04 |
4.28 |
» |
» |
2.01 |
Ca 10 [P 4.98 S 0.51 C 0.51 O 24 ](OH) 2 |
|
51 |
56.58 |
» |
» |
39.53 |
2.92 |
0.97 |
» |
1.82 |
Ca 10 [P 5.51 S 0.16 C 0.36 O 24 ](OH) 1.95 |
|
52 |
58.84 |
» |
» |
39.33 |
1.83 |
н.о. |
» |
1.92 |
Ca 10 [P 5.27 S 0.36 C 0.37 O 24 ](OH) 1.99 |
Продолжение таблицы 4 / Continuation of Table 4
|
№ обр. Sample No. |
CaO |
SrO |
MnO |
P 2 O 5 |
SO 3 |
SiO 2 |
Cl |
Ca/P ат |
Формулы Formulas |
|
53 |
58.23 |
» |
» |
38.96 |
2.81 |
» |
» |
1.89 |
Ca 10 [P 5.28 S 0.34 C 0.38 O 24 ](OH) 1.96 |
|
54 |
58.65 |
» |
» |
37.56 |
3.4 |
» |
0.39 |
1.98 |
Ca 10 [P 5.05 S 0.41 C 0.54 O 24 ]Cl 0.11 (OH) 1.76 |
|
55 |
56.66 |
» |
» |
40.22 |
3.12 |
» |
н. о. |
1.79 |
Ca 10 [P 5.6 S 0.39 C 0.01 O 24 ](OH) 2.38 |
|
56 |
56.63 |
» |
» |
40.96 |
2.12 |
» |
0.29 |
1.75 |
Ca 10 [P 5.7 S 0.26 C 0.04 O 24 ]Cl 0.08 (OH) 2.14 |
|
57 |
57.57 |
» |
» |
40.02 |
2.41 |
» |
н. о. |
1.82 |
Ca 10 [P 4.68 S 0.27 C 1.05 O 24 ](OH) 1.22 |
|
58 |
54.76 |
2.4 |
» |
35.68 |
6.68 |
» |
0.54 |
1.95 |
Ca 10 [P 5.14 S 0.85 C 0.01 O 24 ]Cl 0.16 (OH) 1.22 |
|
59 |
57.44 |
н. о. |
» |
39.52 |
3.04 |
» |
н. о. |
1.84 |
Ca 10 [P 5.43 S 0.37 C 0.2 O 24 ](OH) 2.17 |
|
Врб-4 |
|||||||||
|
60 |
61.07 |
н. о. |
н. о. |
37.21 |
1.72 |
н. о. |
н. о. |
2.08 |
Ca 10 [P 4.81 S 0.2 C 0.99 O 24 ](OH) 1.21 |
|
61 |
58.99 |
» |
» |
39.02 |
1.99 |
» |
» |
1.92 |
Ca 10 [P 5.22 S 0.24 C 0.54 O 24 ](OH) 1.7 |
|
62 |
60.33 |
» |
» |
37.36 |
2.31 |
» |
» |
2.05 |
Ca 10 [P 4.88 S 0.27 C 0.85 O 24 ](OH) 1.42 |
|
63 |
61.12 |
» |
» |
37.12 |
1.76 |
» |
» |
1.95 |
Ca 10 [P 5.13 S 0.22 C 0.65 O 24 ](OH) 1.57 |
|
64 |
60.13 |
» |
» |
38.17 |
1.7 |
» |
» |
2.0 |
Ca 10 [P 5.01 S 0.2 C 0.79 O 24 ](OH) 1.41 |
|
65 |
60.71 |
» |
» |
37.82 |
1.47 |
» |
» |
2.04 |
Ca 10 [P 4.91 S 0.17 C 0.92 O 24 ](OH) 1.25 |
|
66 |
61.26 |
» |
» |
37.33 |
1.41 |
» |
» |
2.08 |
Ca 10 [P 4.81 S 0.16 C 1.03 O 24 ](OH) 1.13 |
|
67 |
58.73 |
» |
» |
38.94 |
2.33 |
» |
» |
1.91 |
Ca 10 [P 5.23 S 0.28 C 0.49 O 24 ](OH) 1.79 |
|
68 |
57.36 |
2.26 |
» |
37.9 |
2.48 |
» |
» |
1.92 |
(Ca 9.79 Sr 0.21 ) 10 [P 5.1 S 0.3 C 0.6 O 24 ](OH) 1.7 |
|
69 |
59.71 |
1.76 |
» |
36.57 |
1.96 |
» |
» |
2.07 |
(Ca 9.84 Sr 0.16 ) 10 [P 4.76 S 0.37 C 0.2 O 24 ](OH) 1.22 |
|
70 |
58.41 |
1.41 |
» |
38.06 |
2.12 |
» |
» |
1.95 |
(Ca 9.87 Sr 0.13 ) 10 [P 5.07 S 0.25 C 0.68 O 24 ](OH) 1.57 |
|
плиоцен |
59.16 |
0.14 |
0 |
38.37 |
2.23 |
0.02 |
0.05 |
1.97 |
(Ca 9.79—10 Sr 0—0.21 ) 10 [P 4.4—5.8 S 0—0.85 |
|
Pliocene |
±1.57 |
±0.52 |
±1.32 |
±1.02 |
±0.13 |
±0.14 |
±0.11 |
C 0.1—1.17 O 24 ] Cl 0—0.19 (OH) 0.44—2.17 |
|
|
Врб-5 |
|||||||||
|
71 |
59.92 |
1.62 |
н. о. |
37.39 |
1.07 |
н. о. |
н. о. |
2.03 |
(Ca 9.86 Sr 0.14 ) 10 [P 4.85 S 0.12 C 1.03 O 24 ](OH) 1.09 |
|
72 |
59.04 |
1.71 |
» |
37.74 |
1.51 |
» |
» |
1.98 |
(Ca 9.85 Sr 0.15 ) 10 [P 4.96 S 0.18 C 0.86 O 24 ] (OH) 1.32 |
|
73 |
59.65 |
н. о. |
» |
40.35 |
» |
» |
1.87 |
Ca 10 [P 5.34 C 0.66 O 24 ](OH) 1.34 |
|
|
74 |
60.71 |
» |
» |
39.29 |
» |
» |
1.96 |
Ca 10 [P 5.1 C 0.9 O 24 ](OH) 1.1 |
|
|
75 |
60.44 |
» |
» |
37.71 |
1.45 |
» |
0.4 |
2.03 |
Ca 10 [P 4.92 S 0.17 C 0.91 O 24 ]Cl 0.11 (OH) 1.05 |
|
76 |
61.32 |
» |
» |
38.68 |
» |
» |
н. о. |
2.01 |
Ca 10 [P 4.98 C 1.02 O 24 ](OH) 0.98 |
|
77 |
60.89 |
» |
» |
37.35 |
1.38 |
» |
0.38 |
1.99 |
Ca 10 [P 5.01 S 0.16 C 0.83 O 24 ]Cl 0.1 (OH) 1.23 |
|
78 |
57.87 |
» |
» |
40.72 |
» |
» |
н.о. |
1.96 |
Ca 10 [P 5.09 S 0.17 C 0.74 O 24 ](OH) 1.43 |
|
79 |
57.48 |
» |
» |
40.54 |
1.51 |
» |
0.47 |
1.8 |
Ca 10 [P 5.56 S 0.18 C 0.26 O 24 ]Cl 0.13 (OH) 1.79 |
|
80 |
58.62 |
» |
» |
40.12 |
1.26 |
» |
н. о. |
1.85 |
Ca 10 [P 5.4 S 0.15 C 0.45 O 24 ](OH) 1.7 |
|
81 |
61.84 |
» |
» |
37.71 |
» |
» |
0.45 |
2.08 |
Ca 10 [P 4.81 C 1.19 O 24 ]Cl 0.12 (OH) 0.69 |
|
82 |
58.79 |
» |
» |
37.78 |
1.84 |
1.59 |
н.о. |
1.97 |
Ca 10 [P 5.07 Si 0.25 S 0.22 C 0.46 O 24 ](OH) 1.51 |
|
83 |
58.88 |
» |
» |
37.75 |
2.9 |
н.о. |
0.47 |
1.98 |
Ca 10 [P 5.06 S 0.34 C 0.6 O 24 ]Cl 0.13 (OH) 1.61 |
|
84 |
57.89 |
» |
» |
38.11 |
1.91 |
1.53 |
0.56 |
1.93 |
Ca 10 [P 5.19 Si 0.25 S 0.23 C 0.33 O 24 ]Cl 0.15 (OH) 1.5 |
|
85 |
60.18 |
» |
» |
38.61 |
1.21 |
н. о. |
н. о. |
1.98 |
Ca 10 [P 5.06 S 0.14 C 0.8 O 24 ]Cl 0.13 (OH) 1.34 |
|
86 |
58.97 |
1.53 |
» |
37.39 |
2.11 |
» |
» |
2.0 |
(Ca 0.99 Sr 0.01 ) 10 [P 4.93 S 0.02 C 1.05 O 24 ] Cl 0.13 (OH) 0.97 |
|
87 |
59.74 |
н.о. |
» |
38.55 |
1.71 |
» |
» |
1.96 |
Ca 10 [P 5.09 S 0.2 C 0.71 O 24 ]Cl 0.13 (OH) 1.48 |
|
Врб-6 |
|||||||||
|
88 |
61.78 |
н.о. |
н.о. |
37.3 |
0.92 |
н. о. |
н. о. |
2.1 |
Ca 10 [P 4.76 S 0.1 C 1.14 O 24 ](OH) 0.96 |
|
89 |
60.55 |
» |
» |
38.12 |
1.33 |
» |
» |
2.01 |
Ca 10 [P 4.96 S 0.15 C 0.89 O 24 ](OH) 1.26 |
|
90 |
60.84 |
» |
» |
38.15 |
1.01 |
» |
» |
2.02 |
Ca 10 [P 4.95 S 0.12 C 0.93 O 24 ](OH) 1.19 |
|
91 |
59.9 |
1.69 |
» |
37.14 |
1.27 |
» |
» |
2.04 |
(Ca 9.85 Sr 0.15 )[P 4.82 S 0.15 C 1.03 O 24 ](OH) 1.12 |
|
92 |
59.99 |
н. о. |
» |
38.82 |
1.19 |
» |
» |
1.96 |
Ca 10 [P 5.1 S 0.14 C 0.76 O 24 ](OH) 1.38 |
|
93 |
61.25 |
» |
» |
38.75 |
н. о. |
» |
» |
2.0 |
Ca 10 [P 4.99 C 1.01 O 24 ](OH) 0.99 |
|
94 |
60.31 |
» |
» |
38.59 |
1.1 |
» |
» |
1.98 |
Ca 10 [P 5.05 S 0.13 C 0.82 O 24 ](OH) 1.31 |
|
95 |
61.34 |
» |
» |
37.56 |
1.1 |
» |
» |
2.07 |
Ca 10 [P 4.83 S 0.13 C 1.04 O 24 ](OH) 1.09 |
|
Врб-7 |
|||||||||
|
96 |
59.11 |
н. о. |
н. о. |
37.49 |
1.57 |
1.36 |
0.47 |
2.0 |
Ca 10 [P 5 Si 0.22 S 0.19 C 0.59 O 24 ]Cl 0.13 (OH) 1.25 |
|
97 |
59.06 |
» |
» |
38.36 |
2.58 |
н. о. |
н. о. |
1.95 |
Ca 10 [P 5.12 S 0.31 C 0.57 O 24 ](OH) 1.74 |
|
98 |
59.41 |
» |
» |
37.56 |
3.33 |
» |
» |
2.01 |
Ca 10 [P 4.99 S 0.39 C 0.62 O 24 ](OH) 1.77 |
|
99 |
59.12 |
» |
» |
38.75 |
2.13 |
» |
» |
1.93 |
Ca 10 [P 5.17 S 0.25 C 0.58 O 24 ](OH) 1.67 |
|
Тит-1 |
|||||||||
|
100 |
63.52 |
н. о. |
н. о. |
36.48 |
н. о. |
н. о. |
н. о. |
2.21 |
Ca 10 [P 4.53 C 1.47 O 24 ](OH) 0.53 |
|
101 |
60.52 |
» |
» |
38.15 |
1.33 |
» |
» |
2.01 |
Ca 10 [P 4.97 S 0.15 C 0.88 O 24 ](OH) 1.26 |
|
102 |
60.72 |
» |
1.4 |
36.69 |
1.19 |
» |
» |
2.14 |
(Ca 9.98 Mn 0.02 ) 10 [P 4.67 S 0.13 C 1.2 O 24 ] (OH) 0.93 |
|
103 |
59.69 |
1.52 |
н.о. |
37.68 |
1.11 |
» |
» |
2.03 |
(Ca 9.99 Sr 0.01 ) 10 [P 4.91 S 0.13 C 0.96 O 24 ] (OH) 1.17 |
|
104 |
61.15 |
н. о. |
» |
37.66 |
1.19 |
» |
» |
2.06 |
Ca 10 [P 4.86 S 0.01 C 1.13 O 24 ](OH) 0.88 |
Продолжение таблицы 4 / Continuation of Table 4
|
№ обр. Sample No. |
CaO |
SrO |
MnO |
P 2 O 5 |
SO 3 |
SiO 2 |
Cl |
Ca/P ат |
Формулы Formulas |
|
105 |
61.0 |
» |
» |
37.67 |
1.33 |
» |
» |
1.93 |
Ca 10 [P 5.17 S 0.14 C 0.69 O 24 ](OH) 1.45 |
|
106 |
61.0 |
» |
» |
39.0 |
н. о. |
» |
» |
1.98 |
Ca 10 [P 5.04 C 0.96 O 24 ](OH) 1.04 |
|
Тит-2 |
|||||||||
|
107 |
60.83 |
н.о. |
н.о. |
36.87 |
2.3 |
н.о. |
н.о. |
2.09 |
Ca 10 [P 4.78 S 0.26 C 0.96 O 24 ](OH) 1.3 |
|
108 |
62.07 |
» |
» |
35.92 |
2.01 |
» |
» |
2.19 |
Ca 10 [P 4.56 S 0.23 C 1.23 O 24 ](OH) 1.1 |
|
109 |
61.17 |
» |
» |
36.99 |
1.84 |
» |
» |
2.1 |
Ca 10 [P 4.77 S 0.21 C 1.02 O 24 ](OH) 1.19 |
|
Тит-3 |
|||||||||
|
110 |
61.35 |
н. о. |
н. о. |
36.82 |
1.83 |
н. о. |
н. о. |
2.11 |
Ca 10 [P 4.73 S 0.21 C 1.06 O 24 ](OH) 1.15 |
|
111 |
59.28 |
» |
» |
39.46 |
1.26 |
» |
» |
1.9 |
Ca 10 [P 5.25 S 0.15 C 0.6 O 24 ](OH) 1.55 |
|
112 |
60.5 |
» |
» |
37.7 |
1.8 |
» |
» |
2.02 |
Ca 10 [P 4.94 S 0.21 C 0.85 O 24 ](OH) 1.36 |
|
113 |
58.92 |
1.37 |
» |
38.82 |
0.89 |
» |
» |
1.85 |
(Ca 9.88 Sr 0.12 ) 10 [P 5.35 S 0.1 C 0.55 O 24 ] (OH) 1.55 |
|
114 |
62.07 |
н. о. |
» |
36.9 |
1.03 |
» |
» |
2.13 |
Ca 10 [P 4.69 S 0.12 C 1.19 O 24 ](OH) 0.93 |
|
Таврида / Tavrid |
a |
||||||||
|
115 |
58.39 |
н. о. |
н.о. |
40.96 |
н.о. |
н. о. |
0.65 |
1.81 |
Ca 10 [P 5.53 C 0.47 O 24 ]Cl 0.18 (OH) 1.35 |
|
116 |
57.33 |
» |
» |
41.28 |
0.9 |
» |
0.49 |
1.76 |
Ca 10 [P 5.68 S 0.11 C 0.21 O 24 ]Cl 0.14 (OH) 1.76 |
|
117 |
57.47 |
» |
» |
40.57 |
1.35 |
» |
0.61 |
1.8 |
Ca 10 [P 5.57 S 0.16 C 0.27 O 24 ]Cl 0.17 (OH) 1.72 |
|
118 |
56.83 |
» |
» |
41.64 |
1.1 |
» |
0.43 |
1.73 |
Ca 10 [P 5.78 S 0.14 C 0.08 O 24 ]Cl 0.12 (OH) 1.94 |
|
119 |
57.43 |
» |
» |
40.65 |
1.38 |
» |
0.54 |
1.79 |
Ca 10 [P 5.58 S 0.17 C 0.25 O 24 ]Cl 0.15 (OH) 1.77 |
|
120 |
56.61 |
» |
» |
41.67 |
1.34 |
» |
0.38 |
1.72 |
Ca 10 [P 5.81 S 0.17 C 0.02 O 24 ]Cl 0.11 (OH) 2.04 |
|
121 |
58.46 |
» |
» |
41.08 |
н. о. |
» |
0.46 |
1.72 |
Ca 10 [P 5.43 C 0.57 O 24 ]Cl 0.13 (OH) 1.3 |
|
122 |
57.67 |
» |
» |
40.77 |
0.92 |
» |
0.64 |
1.79 |
Ca 10 [P 5.58 S 0.11 C 0.31 O 24 ]Cl 0.18 (OH) 1.62 |
|
123 |
59.4 |
» |
» |
38.63 |
1.47 |
» |
0.5 |
1.95 |
Ca 10 [P 5.13 S 0.17 C 0.7 O 24 ]Cl 0.31 (OH) 1.16 |
|
124 |
57.26 |
» |
» |
40.5 |
1.63 |
» |
0.61 |
1.79 |
Ca 10 [P 5.57 S 0.2 C 0.23 O 24 ]Cl 0.17 (OH) 1.8 |
|
125 |
57.99 |
» |
» |
40.61 |
1.0 |
» |
0.4 |
1.81 |
Ca 10 [P 5.52 S 0.12 C 0.36 O 24 ]Cl 0.26 (OH) 1.5 |
|
126 |
58.75 |
» |
» |
40.73 |
н.о. |
» |
0.52 |
1.83 |
Ca 10 [P 5.47 C 0.53 O 24 ]Cl 0.15 (OH) 1.32 |
|
127 |
58.01 |
» |
» |
40.07 |
1.43 |
» |
0.49 |
1.83 |
Ca 10 [P 5.45 S 0.17 C 0.38 O 24 ]Cl 0.13 (OH) 1.9 |
|
128 |
55.56 |
1.76 |
» |
40.93 |
1.33 |
» |
0.42 |
1.72 |
(Ca 9.83 Sr 0.17 ) 10 [P 5.7 S 0.16 C 0.14 O 24 ]Cl 0.18 (OH) 1.62 |
|
129 |
57.56 |
н. о. |
» |
40.67 |
1.41 |
» |
0.36 |
1.79 |
Ca 10 [P 5.57 S 0.17 C 0.26 O 24 ]Cl 0.1 (OH) 1.81 |
|
130 |
59.62 |
» |
» |
38.83 |
1.55 |
» |
н. о. |
1.95 |
Ca 10 [P 5.14 C 0.86 O 24 ](OH) 1.14 |
|
плейстоцен |
|||||||||
|
(гелазий) |
59.55 |
0.19 |
0.02 |
38.75 |
1.22 |
0.07 |
0.18 |
1.95 |
(Ca 9.86—10 Sr 0—0.14 ) 10 [P 4.53—5.81 Si 0—0.25 |
|
Pleistocene (Gelasian) |
±1.61 |
±0.52 |
±0.18 |
±1.51 |
±0.74 |
±0.33 |
±0.24 |
±0.12 |
S 0—0.39 C 0.02—1.47 O 24 ]Cl 0—0.31 (OH) 0.53—2.04 |
|
Врб-8 |
|||||||||
|
131 |
57.91 |
н.о. |
н.о. |
42.09 |
н.о. |
н.о. |
н.о. |
1.74 |
Ca 10 [P 5.73 C 0.27 O 24 ](OH) 1.71 |
|
132 |
57.12 |
» |
» |
41.41 |
1.47 |
» |
» |
1.75 |
Ca 10 [P 5.72 S 0.18 C 0.1 O 24 ](OH) 2.08 |
|
133 |
57.39 |
» |
» |
40.23 |
2.38 |
» |
» |
1.81 |
Ca 10 [P 5.53 S 0.29 C 0.18 O 24 ](OH) 2.01 |
|
134 |
57.08 |
» |
» |
41.75 |
1.17 |
» |
» |
1.73 |
Ca 10 [P 5.77 S 0.14 C 0.09 O 24 ](OH) 2.05 |
|
Врб-9 |
|||||||||
|
135 |
59.69 |
н.о. |
н.о. |
40.31 |
н.о. |
н.о. |
н.о. |
1.88 |
Ca 10 [P 5.33 C 0.67 O 24 ](OH) 1.33 |
|
136 |
57.46 |
» |
» |
42.54 |
» |
» |
» |
1.71 |
Ca 10 [P 5.84 C 0.16 O 24 ](OH) 1.84 |
|
137 |
59.75 |
» |
» |
39.26 |
0.99 |
» |
» |
1.93 |
Ca 10 [P 5.18 S 0.12 C 0.7 O 24 ](OH) 1.42 |
|
138 |
57.15 |
» |
» |
41.18 |
1.67 |
» |
» |
1.76 |
Ca 10 [P 5.68 S 0.2 C 0.12 O 24 ](OH) 0.4 |
|
139 |
60.33 |
» |
» |
39.67 |
н.о. |
» |
» |
1.93 |
Ca 10 [P 5.19 C 0.81 O 24 ](OH) 1.19 |
|
140 |
60.26 |
» |
» |
38.57 |
» |
» |
» |
1.99 |
Ca 10 [P 5.05 Si 0.18 C 0.77 O 24 ](OH) 1.05 |
|
141 |
56.43 |
» |
» |
37.71 |
» |
5.86 |
» |
1.9 |
Ca 10 [P 5.27 Si 0.36 C 0.37 O 24 ](OH) 1.27 |
|
142 |
57.13 |
» |
» |
41.43 |
1.44 |
» |
» |
1.75 |
Ca 10 [P 5.72 S 0.18 C 0.1 O 24 ](OH) 1.08 |
|
143 |
59.36 |
» |
» |
38.04 |
н.о. |
2.6 |
» |
1.98 |
Ca 10 [P 5.05 Si 0.41 C 0.54 O 24 ](OH) 1.05 |
|
Врб-10 |
|||||||||
|
144 |
57.51 |
н. о. |
н.о. |
40.85 |
1.64 |
н. о. |
н. о. |
1.78 |
Ca 10 [P 5.6 S 0.2 C 0.2 O 24 ](OH) 2 |
|
145 |
58.67 |
» |
» |
39.84 |
1.49 |
» |
» |
1.87 |
Ca 10 [P 5.36 S 0.18 C 0.46 O 24 ](OH) 1.72 |
|
146 |
59.36 |
» |
» |
39.37 |
1.27 |
» |
» |
1.91 |
Ca 10 [P 5.23 S 0.15 C 0.62 O 24 ](OH) 1.53 |
|
147 |
60.77 |
» |
» |
37.83 |
1.4 |
» |
» |
2.04 |
Ca 10 [P 4.91 S 0.16 C 0.93 O 24 ](OH) 1.18 |
|
148 |
59.01 |
» |
» |
39.24 |
1.75 |
» |
» |
1.91 |
Ca 10 [P 5.24 S 0.4 C 0.36 O 24 ](OH) 2.04 |
|
149 |
57.58 |
» |
» |
40.62 |
1.8 |
» |
» |
1.8 |
Ca 10 [P 5.56 S 0.22 C 0.22 O 24 ](OH) 2 |
|
150 |
59.12 |
» |
» |
39.33 |
1.55 |
» |
» |
1.87 |
Ca 10 [P 5.34 S 0.19 C 0.47 O 24 ](OH) 1.72 |
|
151 |
60.91 |
» |
» |
37.67 |
1.42 |
» |
» |
2.05 |
Ca 10 [P 4.88 S 0.16 C 0.96 O 24 ](OH) 1.2 |
|
плейстоцен (калабрий) |
58.57 |
0 |
0 |
39.95 |
1.02 |
0.4 |
0 |
1.86 |
Ca 10 [P 4.88—5.84 Si 0—0.41 S 0—0.4 C 0.09—0.93 O 24 ] |
|
Pleistocene (Calabrian) |
±1.39 |
±1.48 |
±0.79 |
±1.37 |
±0.1 |
(OH) 0.4—2.08 |
|||
Окончание таблицы 4 / End of Table 4
|
№ обр. Sample No. |
CaO |
SrO |
MnO |
P 2 O 5 |
SO 3 |
SiO 2 |
Cl |
Ca/P ат |
Формулы Formulas |
|
Тит-4 |
|||||||||
|
152 |
62.26 |
н.о. |
н.о. |
36.6 |
1.14 |
н.о. |
н.о. |
2.15 |
Ca 10 [P 4.64 S 0.13 C 1.23 O 24 ](OH) 0.9 |
|
153 |
58.71 |
» |
» |
39.97 |
1.32 |
» |
» |
1.86 |
Ca 10 [P 5.37 S 0.15 C 0.48 O 24 ](OH) 1.67 |
|
154 |
59.86 |
» |
» |
38.75 |
1.39 |
» |
» |
1.96 |
Ca 10 [P 5.11 S 0.19 C 0.7 O 24 ](OH) 1.49 |
|
155 |
56.78 |
» |
» |
41.08 |
2.14 |
» |
» |
1.75 |
Ca 10 [P 5.71 S 0.26 C 0.03 O 24 ](OH) 2.23 |
|
156 |
59.45 |
» |
» |
38.51 |
2.04 |
» |
» |
1.96 |
Ca 10 [P 5.11 S 0.24 C 0.65 O 24 ](OH) 1.59 |
|
157 |
58.68 |
» |
» |
40.18 |
1.14 |
» |
» |
1.85 |
Ca 10 [P 5.4 S 0.14 C 0.46 O 24 ](OH) 1.68 |
|
158 |
60.88 |
» |
» |
37.21 |
1.91 |
» |
» |
2.08 |
Ca 10 [P 4.8 S 0.22 C 0.98 O 24 ](OH) 1.24 |
|
Тит-5 |
|||||||||
|
159 |
58.59 |
н.о. |
н.о. |
40.24 |
1.17 |
н.о. |
н.о. |
1.84 |
Ca 10 [P 5.42 S 0.14 C 0.44 O 24 ](OH) 1.7 |
|
160 |
57.19 |
» |
» |
41.57 |
1.24 |
» |
» |
1.74 |
Ca 10 [P 5.73 S 0.15 C 0.12 O 24 ](OH) 2.03 |
|
161 |
63.46 |
» |
» |
35.25 |
1.29 |
» |
» |
2.28 |
Ca 10 [P 4.38 S 0.14 C 1.48 O 24 ](OH) 0.66 |
|
Тит-6 |
|||||||||
|
162 |
58.2 |
н.о. |
н.о. |
39.38 |
2.42 |
н.о. |
н.о. |
1.88 |
Ca 10 [P 5.33 S 0.28 C 0.39 O 24 ](OH) 1.89 |
|
163 |
60.14 |
» |
» |
37.76 |
2.1 |
» |
» |
2.02 |
Ca 10 [P 4.95 S 0.23 C 0.82 O 24 ](OH) 1.41 |
|
164 |
58.02 |
» |
» |
40.57 |
1.31 |
» |
» |
1.81 |
Ca 10 [P 5.52 S 0.17 C 0.31 O 24 ](OH) 1.86 |
|
166 |
56.22 |
» |
» |
41.1 |
2.68 |
» |
» |
1.73 |
Ca 10 [P 5.77 S 0.23 O 24 ](OH) 2.23 |
|
167 |
59.35 |
» |
» |
39.55 |
1.1 |
» |
» |
1.9 |
Ca 10 [P 5.26 S 0.13 C 0.61 O 24 ](OH) 1.52 |
|
168 |
56.82 |
» |
» |
41.42 |
1.76 |
» |
» |
1.74 |
Ca 10 [P 5.75 S 0.22 C 0.03 O 24 ](OH) 2.19 |
|
169 |
64.95 |
» |
» |
35.05 |
н.о. |
» |
» |
2.35 |
Ca 10 [P 5.04 C 0.96 O 24 ](OH) 0.25 |
|
170 |
56.43 |
» |
» |
42.23 |
1.34 |
» |
» |
1.69 |
Ca 10 [P 5.9 S 0.1 O 24 ](OH) 2.1 |
|
171 |
60.83 |
» |
» |
36.3 |
2.87 |
» |
» |
2.12 |
Ca 10 [P 4.71 S 0.33 C 0.96 O 24 ](OH) 1.37 |
|
155 |
58.89 |
» |
» |
39.0 |
2.11 |
» |
» |
1.91 |
Ca 10 [P 5.22 S 0.25 C 0.53 O 24 ](OH) 1.72 |
|
Тит-7 |
|||||||||
|
156 |
56.62 |
н.о. |
н.о. |
40.22 |
3.16 |
н.о. |
н.о. |
1.78 |
Ca 10 [P 5.6 S 0.16 S 0.39 C 0.01 O 24 ](OH) 2.38 |
|
голоцен |
59.16 |
0 |
0 |
39.14 |
1.7 |
0 |
0 |
1.92 |
Ca 10 [P 4.38—5.9 S 0—0.33 C 0—1.48 O 24 ] |
|
Holocene |
±2.34 |
±2.09 |
±0.73 |
±0.18 |
OH) 0.25—2.38 |
||||
Примечание: н. о. —не обнаружено.
Note: н. о. — not detected.

Органический матрикс
Присутствие органического вещества в ископаемых костях легко выявляется термическим методом (Смирнов и др., 2009; Votyakov et al., 2010). На полученных нами кривых нагревания в диапазоне 20—600 °С зарегистрированы все три характерных для костей термических эффекта, но весьма различно проявившихся у костей разного возраста. В случае костей плиоцен-па-леоплейстоценовых верблюдов на кривых нагревания
регистрируются два малоинтенсивных термических эффекта (рис. 16): 1) эндотермический с экстремумом при 105—115 °С, обусловленный низкотемпературной дегидратацией (абсорбированная вода) кости; 2) экзотермический с экстремумом при 300—329 °С, отражающий выгорание органического вещества с потерей веса 1—2.5 %. Кроме того, часто наблюдаются дополнительные эндотермические пики в области 700—800 °С, обусловленные термодиссоциацией карбонатов.
Рис. 16 . Типичные данные ДТА костей плиоценовых (a, b) и плейстоценовых (c) верблюдов: 1, 2 — кривые соответственно нагревания и потери веса. Цифры на кривых нагревания — температуры экстремумов в °С. Проанализированные образцы: a — Тит-2, b — Тит-3, c — Тит-1
Fig. 16 . Typical DTA data on the bones of Pliocene (a, b) and Pleistocene (c) camels: 1, 2 — heating and weight loss curves, respectively. The numbers on the heating curves are the extreme temperatures in °C. Analyzed samples: a — Tit-2, b — Tit-3, c — Tit-1 20
В случае костей верблюдов голоценового возраста на кривых нагревания присутствуют три термических эффекта (рис. 17): 1) эндотермический пик с экстремумом при 100—112 °С, обусловленный низкотемпературной дегидратацией; 2) преобладающий по интенсивности экзотермический пик В1 с экстремумом при 320—340 °С; 3) дополнительный экзотермический пик В2 с экстремумом при 340—380 °С. Оба экзотермических эффекта отражают выгорание органического вещества с потерей от 7 до 17 % исходной массы и регистрируемым выделением Н2О, NO, CO2.
Согласно полученным данным, термические свойства костей строго коррелируются с геологическим возрастом ископаемых верблюдов. Термические эффекты выгорания костного органического вещества хорошо реализуются только в результатах анализа голоценовых костей. В случае более древних костей эти эффекты проявляются незначительно. Очевидно, что выявленная разница результатов ДТА обусловлена резким сокращением содержания коллагена с удрев-нением костей. Последнее подтверждается непосредственным определением содержания Сорг. В костях
наиболее молодых — голоценовых — верблюдов содержание Сорг оценивается в (9.78 ± 1.46) мас. %, в костях калабрийских верблюдов оно снижается в 1.5— 2 раза — до (5.67 ± 3.01) мас. %, а в костях миоцен-ге-лазийских верблюдов падает еще в 2.5 раза — до (2.24 ± 1.6) мас. %.
Выделение костного коллагена из исследуемых костей осуществлялось методом химической деминерализации. Наиболее высокий показатель выхода костного органического вещества — (13.98 ± 5.92) мас. % — показали кости голоценовых верблюдов. В случае костей плейстоценовых верблюдов этот выход сокращается почти в 2 раза — до (7.2 ± 3.23) мас. %, а в случае миоцен-плиоценовых уменьшается еще в 2.5 раза — до (2.79 ± 3.25) мас. %. Полученный костный коллаген широко варьируется по окраске от бурого и темно-бурого в случае плиоцен-плейстоценовых костей до буровато-желтого и желтого в голоценовых (рис. 18). Это обусловлено окислением и структурной деградацией органического вещества в ходе фоссилизации захороненных костей, степень которых, разумеется, возрастает с геологическим возрастом костей.
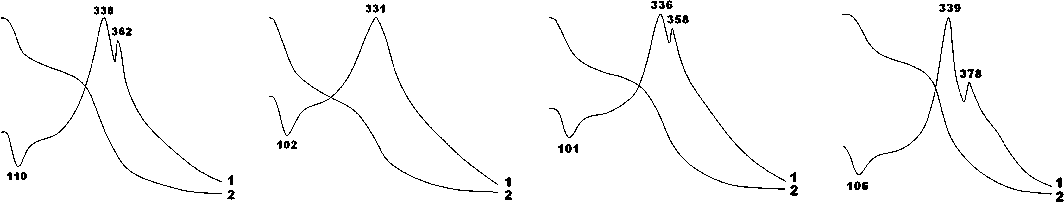
Рис. 17 . Типичные данные ДТА костей голоценовых верблюдов: 1, 2 — кривые соответственно нагревания и потери веса. Цифры на кривых нагревания — температуры экстремумов в °С. Проанализированные образцы, слева направо: Тит-4, Тит-5, Тит-6, Тит-7
Fig. 17 . Typical DTA data for the bones of Holocene camels: 1, 2 — heating and weight loss curves, respectively. The numbers on the heating curves are the extreme temperatures in °C. Analyzed samples, from left to right: Tit-4, Tit-5, Tit-6, Tit-7

Рис. 18 . Внешний вид коллагена, выделенного из костей раннеплейстоценовых (гелазийских) (Тит-1, 2, 3) и голоценовых (Тит-4, 5, 6, 7) верблюдов. Стрелками показаны микровключения минеральных частиц
Fig. 18. Appearance of collagen isolated from bones of Early Pleistocene (Gelasian) (Tit-1, 2, 3) and Holocene (Tit-4, 5, 6, 7) camels. Arrows show microinclusions of mineral particles
Таблица 6 . Химический состав примесей в коллагене ископаемых верблюдов, мас. %
Table 6. Chemical composition of admixtures in fossil camel collagen, wt. %
|
Объекты / Objects |
SiO 2 |
Al2O3 |
Fe 2 O 3 |
CaO |
P 2 O 5 |
CO 3 |
SO 3 |
Cl |
Сумма Sum |
|
Верблюды плиоцен-плейстоценовые Pliocene-Pleistocene camels |
1.56 ± 1.4 |
0.69 ± 0.6 |
25.77 ± 15.9 |
19.29 ± 12.25 |
3.57 ± 1.28 |
11.6 ± 8.74 |
0.84 ± 0.55 |
н. о. |
63.33 ± 14.74 |
|
Верблюды голоценовые Holocene camels |
0.13 ± 0.25 |
0.08 ± 0.21 |
0.08 ± 0.23 |
0.47 ± 1.06 |
0.04 ± 0.14 |
0.33 ± 0.78 |
0.43 ± 0.24 |
2.08 ± 1 |
3.63 ± 1.88 |
Содержание углерода и азота в органической части коллагена колеблется в пределах соответственно 26—32 и 10.2—11.66 мас. %, снижаясь в направлении от голоценовых костей к палеоплейстоценовым. Атомное отношение С/N изменяется в рамках 3.1—3.43. Анализ содержания неорганических примесей указал на обогащение ими органического матрикса в направлении от молодых костей к древним в 15—20 раз (табл. 6). При этом для некоторых компонентов установлены аномально высокие расхождения. Так, при переходе от относительно молодых костей к древним содержание СaO и СО2 возрастает в 35—40 раз, а содержание Fe2O3 и вовсе подскакивает более чем в 300 раз. Очевидно, что такие скачки концентрации обусловлены отмеченной
выше эпигенетической карбонатизацией и ожелезне-нием, степень которых сильно возрастает по мере удрев-нения костей. Особый интерес вызывают вариации содержания в коллагене серы, которая изначально является важным компонентом аминокислот (Анищенко, Шанина, 2008). В нашем случае валовое содержание серы в органическом матриксе костей при переходе от молодых костей к древним не снижается, как можно было ожидать, а возрастает в два раза.
Аминокислотный состав костного коллагена удалось проанализировать в костях позднеплейстоценового и голоценового возраста. В результате проведенного анализа в составе органических веществ обнаружены 15 аминокислот (табл. 7), представляющих обе
Таблица 7. Аминокислотный состав костного коллагена верблюдов позднеплейстоценового (ВРБ-9) и голоценового (остальные в последовательности омоложения) возраста, мг/г (%)
Table 7. Amino acid composition of bone collagen of camels of Late Pleistocene (ВРБ-9) and of Holocene (rest in the rejuvenation sequence) ages, mg/g (%)
Полученные данные показали, что в составе большинства выявленных аминокислот присутствует только L-форма. Исключение представляют аланин, глутаминовая и аспарагиновая кислоты. Как известно, появление D-форм в аминокислотах обусловлено рацемизацией — самопроизвольным неферментативным превращением первичной L-формы в D-форму, которое при жизни происходит очень медленно, но ускоряется после смерти животного и может служить важным критерием как биологического возраста, так и степени фоссилизации ископаемого биоорганического вещества в геологических объектах (Анищенко, Шанина, 2009; Анищенко, Шанина, 2010; Котельникова, 2014; Каткова, Шанина, 2019). В случае исследуемых верблюдов величина весовой пропорции D/L в упомянутых выше аминокислотах, служащая критерием степени рацемизации, колеблется в пределах 0—0.07, коррелируясь с возрастом костей только в аспарагиновой кислоте. В целом полученные значения, во-первых, соответствуют относительно молодым биологически животным, а во-вторых, свидетельствуют о сравнительно хорошей сохранности их остатков (Yoyce, 2006).
Общее содержание аминокислот колеблется в пределах 700—915 мг/г, не коррелируясь с геологическим возрастом костей. Полученные высокие содержания аминокислот, а также доминирование среди индивидуальных аминокислот глицина, аланина, пролина и гидроксипролина подтвержают коллагеновый состав деминерализованных костных остатков и его хорошую сохранность. Пропорция между незаменимыми и заменимыми кислотами колеблется в узких пределах от 0.17 до 0.26, не обнаруживая корреляции с геологическим возрастом верблюдов. По распределению групп и отдельных аминокислот исследованные образцы так-
же довольно однообразны (в направлении геологического омоложения костных остатков).
ВРБ-9 : глицин (А) > пролин (И) > аланин (А) > гидроксипролин (И) > глутаминовая кислота (К) > серин (Г) > аспарагиновая кислота (К) > лейцин (Л) > тирозин (АР) > треонин (Г) > валин (А) > фенилаланин (АР) > лизин (О) > изолейцин (А) > метионин (СС). НЗ/З = 0.24.
ТИТ-5 : глицин (А) > пролин (И) > аланин (А) = гидроксипролин (И) > глутаминовая кислота (К) > аспарагиновая кислота (К) > серин (Г) > треонин (Г) > тирозин (АР) > лейцин (А > метионин (СС) > валин (А) > фенилаланин (АР) > лизин (О) > изолейцин (А). НЗ/З = 0.26.
ТИТ-4 : глицин (А) > пролин (И) > аланин (А) = гидроксипролин (И) > глутаминовая кислота (К) > треонин (Г) > серин (Г) > аспарагиновая кислота (К) > лейцин (А > тирозин (АР) > лизин (О) > фенилаланин (АР) > метионин (СС) > валин (А) > изолейцин (А). НЗ/З = 0.24.
ТИТ-6 : глицин (А) > пролин (И) > гидроксипролин (И) > аланин (А) > глутаминовая кислота (К) > треонин (Г) > аспарагиновая кислота (К) > серин (Г) > тирозин (АР) > лейцин (А > лизин (О) > валин (А) > фенилаланин (АР) > изолейцин (А) > метионин (СС). > НЗ/З = 0.28.
ТИТ-7 : глицин (А) > пролин (И) > аланин (А) > гидроксипролин (И) > глутаминовая кислота (К) > лизин (О) > серин (Г) > тирозин (АР) > треонин (Г) > лейцин (А > валин (А) > аспарагиновая кислота (К) > валин (А) > фенилаланин (АР) > метионин (СС) > изолейцин (А). НЗ/З = 0.17.
Из приведенных последовательностей следует, что в костном коллагене позднеплейстоцен-голоценовых верблюдов реализуется универсальный ряд снижения содержания групп аминокислот в направлении: алифатические > имино > кислые > гидроксильные > ароматические > основные > серусодержащие.
Картины некоторой хронологической деградации коллагена в верблюжьих костях наглядно выявляются при СЭМ-исследованиях. В случае голоценовых костей в коллагене хорошо сохраняется первичная плотность и волокнистость без явных признаков деструкции (рис. 19). В более древних костях первичная волокнистость в коллагене практически отсутствует, будучи за-

Рис. 19 . СЭМ-изображения микростроения коллагена, выделенного из костей голоценовых верблюдов (Тит-4-7). СЭМ-изображения в режиме вторичных электронов
Fig. 19 . SEM images of the microstructure of collagen isolated from the bones of Holocene camels (Tit-4-7). SEM images in secondary electron mode


Рис. 20 . СЭМ-изображения микростроения коллагена, выделенного из костей раннеплейстоценовых (гелазий-ских) верблюдов, с фрагментизацией первичных волокон (a, b) и субмикронными глобулярными формами бактериального происхождения (Тит-3). СЭМ-изображения в режиме вторичных электронов
Fig. 20 . SEM images of the microstructure of collagen isolated from the bones of Early Pleistocene (Gelasian) camels, with fragmentation of primary fibers (a, b) and submicron globular forms of bacterial origin (Tit-3). SEM images in secondary electron mode
мещенной разориентированными фрагментами волокон, иногда до порошковидности и часто в комбинации с микроглобулярными бактериальными формами (рис. 20). Последнее вообще характерно для интенсивно фоссилизированных ископаемых костей (Майдль, Филиппов, 2018). Понятно, что именно бактериальная пиритизация и объясняет отмеченное выше двукратное увеличение содержания серы в коллагене древних костей.
Выявляющаяся хронологическая деградация костного коллагена хорошо демонстрируется изменением спектров комбинационного рассеяния. В спектрах коллагена голоценовых костей регистрируется сильная люминесценция с весьма незначительным проявлением рамановских линий (рис. 21, a). Это свидетельствует о хорошей сохранности органического вещества. В случае древних костей в спектрах КР доминируют ра-
мановские линии G и D, указывающие на присутствие сильно преобразованного (углеродизированного) органического вещества, в сочетании со множеством линий от минеральных примесей (рис. 21, b).
Палеоэкологические изотопныеиндикаторы
Осуществлялись измерения изотопного состава углерода и кислорода в биоапатите, углерода и азота в выделенном из костей коллагене. Анализ коллагена производился в режиме непрерывного потока гелия (CF-IRMS) на аналитическом комплексе, включающем в себя элементный анализатор Flash EA 1112, соединенный через газовый коммутатор Conflo IV с масс-спектрометром Delta V Advantage. В процессе работы были использованы международные стандарты V-PDB, USGS-40 (L-Glutamic acid) и лабораторный стандарт Acetanilide (C8H9NO). Кроме того, определялся изотопный состав стронция, присутствующего как примесь в биоапатите.
В ходе анализа были получены полные изотопные данные по всем исследованным образцам биоапатита (табл. 8). В части коллагена изотопные данные по азоту были получены только для верблюдов моложе гелазия. Это обусловлено тем, что измененный коллаген в костях более древних верблюдов содержит недостаточное количество азота.
Согласно полученным данным, исследованные нами кости ископаемых верблюдов на фоне других плей-стоцен-голоценовых млекопитающих, обитавших на территории Европы и севере Евразии, характеризуются аномальными изотопными свойствами как по биоапатиту (рис. 22), так и костному коллагену (рис. 23). Это выражается в том, что биоапатит в костных остатках верблюдов демонстрирует сочетание относительно изотопно-тяжелого углерода (–11…–4 ‰) и изотопно-тяжелого кислорода (20—26 ‰). По коллагену верблюды отличаются аномально-тяжелым азотом (7— 14 ‰) и большей частью изотопно-утяжеленным углеродом (–22…–14 ‰). Такие особенности могут указывать на обитание верблюдов, в отличие других плей-стоцен-голоценовых животных, в гораздо более теплых климатических условиях.
Для позднемиоценовых верблюдов Paracamelus cf . aguirrei (Ростовская область) по изотопным данным
Рис. 21 . Спектры комбинационного рассеяния, полученные от костного коллагена: a — голоценовых верблюдов (обр. Тит-4-7); b — плиоцен-плейстоценовых верблюдов (обр. Тит-1-3)
Fig. 21 . Raman spectra obtained from bone collagen: (a) Holocene camels (sample Tit-4—7); b — Pliocene-Pleistocene camels (sample Tit-1-3)
Таблица 8. Изотопный состав вещества костных остатков ископаемых верблюдов
Table 8. Isotopic composition of the substance of bone remains of fossil camels
|
№ п/п No. |
№ обр. Sample No. |
Биоапатит / Bioapatite, ‰ |
Коллаген / Collagen, ‰ |
||
|
δ13С PDB |
δ18О SMOW |
δ13C PDB |
15 Air |
||
|
1 |
Врб-1 |
–5.01 |
25.85 |
–32.24 |
н. о. |
|
поздний миоцен / Late Miocene |
–5.01 |
25.85 |
–32.24 |
н. о. |
|
|
2 |
Врб-2 |
–7.21 |
18.14 |
–28.24 |
н. о. |
|
3 |
Врб-3 |
–6.99 |
23.94 |
н. о. |
|
|
4 |
Врб-4 |
–6.75 |
23.64 |
–27.93 |
н. о. |
|
Плиоцен / Pliocene |
–6.19 ± 1.09 |
23.48 ± 3.16 |
30.16 ± 2.4 |
||
|
5 |
Врб-5 |
–7.79 |
23.15 |
–27.62 |
н. о. |
|
6 |
Врб-6 |
–10.45 |
22.72 |
–27.09 |
н. о. |
|
7 |
Врб-7 |
–10.94 |
24.56 |
–26.8 |
н. о. |
|
8 |
Тит-1 |
–8.75 |
22.6 |
–30.71 |
н. о. |
|
9 |
Тит-2 |
–9.76 |
21.94 |
–27.41 |
н. о. |
|
10 |
Тит-3 |
–9.46 |
21.73 |
–26.47 |
н. о. |
|
плейстоцен (гелазийский век) Pleistocene (Gelasian) |
–9.49 ± 1.15 |
22.78 ± 1.01 |
–27.68 ± 1.54 |
н. о. |
|
|
11 |
Врб-8 |
–10.18 |
22.97 |
–18.68 |
8.24 |
|
12 |
Врб-9 |
–9.18 |
24.54 |
–20.03 |
6.53 |
|
13 |
Врб-10 |
–9.61 |
24.99 |
–24.48 |
н. о. |
|
14 |
3.4 |
–9.5 |
24.4 |
–13.75 |
12.52 |
|
15 |
Титов |
н. о. |
–18.75 |
12.42 |
|
|
плейстоцен (калабрийский, тибанийский и поздний века) Pleistocene (Calabrian, Chibanian and Late ages) |
–9.62 ± 0.42 |
24.22 ± 0.87 |
–19.14 ± 7.94 |
7.94 ± 5.15 |
|
|
16 |
Тит-4 |
–8.79 |
23.4 |
–18.69 |
8.09 |
|
17 |
Тит-5 |
–7.74 |
24.65 |
–17.61 |
9.5 |
|
18 |
Тит-6 |
–8.35 |
19.54 |
–17.49 |
11.34 |
|
19 |
Тит-3 |
–7.54 |
24.76 |
–15.46 |
10.74 |
|
Голоцен / Holocene |
–8.11 ± 0.57 |
23.09 ± 2.44 |
–17.31 ± 1.35 |
9.92 ± 1.44 |
|
Примечание . Статистические оценки: среднее ± стандартное отклонение. Н. о. — не определено Note. Statistical estimates: mean ± standard deviation. н. о. — not determined
Рис. 22 . Изотопный состав углерода и кислорода в биоапатите ископаемых костных остатков и в карбонатах эталонных геологических объектов:
1 — плейстоценовые лошади со стоянки Заозерье (Пермская область); 2, 3 — то же соответственно из Якутии и Западной Чукотки; 4, 5 — мамонтовая фауна соответственно с территорий Печорского Приуралья и Западной Сибири; 6 — мамонтовая фауна с Западной Европы; 7 — плейстоценовые лошади, Нидерланды; 8, 9 — люди эпох мезолита и неолита; 10, 11 — соответственно гигантопитек и орангутан, Южный Китай; 12 — карбонатные отложения в современных речных осадках, Пермский край; 13 — морские карбона-толиты; М, ПЛ, ПГ, ПК, ПКТ, Г — верблюды Северной Евразии с геологическим возрастом соответственно позднемиоценовым, плиоценовым, плейстоцен-гелазийским, плейстоцен-калабрийским, плейстоцен-калабрийско-чибанийским, голоценовым. США — данные по североамериканским плейстоценовым верблюдам
Fig. 22 . Isotopic composition of carbon and oxygen in bioapatite of fossil bone remains and in carbonates of reference geological objects:
-
1 — Pleistocene horses from the Zaozerye site (Perm Region); 2, 3 — the same, respectively, from Yakutia and Western Chukotka; 4, 5 —
mammoth fauna, respectively, from the territories of the Pechora Urals and Western Siberia; 6 — manont fauna from Western Europe; 7 — Pleistocene horses, the Netherlands; 8, 9 — people of the Mesolithic and Neolithic eras; 10, 11 — Gigantopithecus and orangutan, respectively, South China; 12 — carbonate deposits in modern river sediments, Perm Territory; 13 — marine carbonatoliths; M, PL, PG, PK, PKT, G — camels of northern Eurasia with a geological age of the Late Miocene, Pliocene, Pleistocene-Gelasian, Pleistocene-Calabrian, Pleistocene-Calabrian-Chibanian, Holocene, respectively. United States — data on North American Pleistocene camels
Рис. 23 . Изотопный состав углерода и азота в коллагене ископаемых костных остатков:
1 — плейстоценовые лошади со стоянки Заозерье (Пермская область); 2 — мамонтовая фауна с территории Печорского Приуралья; 3—12 — плейстоценовые лошади с территорий соответственно Германии, Западной Европы, Франции, Бельгии, Украины, Якутии, Западной Чукотки, Аляски; 13 — мамонтовая фауна с территории Западной Сибири; 14 — лошади медного века, Румыния; 15 — средневековые лошади с европейской части России; 16, 17 — современные лошади с территории соответственно Польши и Румынии. ПКТ, ПН, Г — верблюды Евразии с геологическим возрастом соответственно плейстоцен (калабрий-чибаний), поздний плейстоцен, голоцен. Ландшафты приведены по Г. Бошрену (Bocherences, 2003; Bocherences et al., 2003). С 3 , САМ, С 4 — типы фотосинтеза в рамках цикла Кальвина—Бенсона—Бассама (Bassham et al., 1950) в последовательности от растений лесостепных обстановок (С 3 ) к растениям степей и саванн (САМ) и далее к растениям пустынь (С 4 )
Fig. 23 . Isotopic composition of carbon and nitrogen in the collagen of fossil bone remains:
1 — Pleistocene horses from the Zaozerye site (Perm region); 2 — mammoth fauna from the territory of the Pechora Cis-Urals; (3—12) Pleistocene horses from the territories of Germany, Western Europe, France, Belgium, Ukraine, Yakutia, Western Chukotka, and Alaska, respectively; 13 — mammoth fauna from the territory of Western Siberia; 14 — copper age horses, Romania; 15 — medieval horses from the European part of Russia; 16, 17 — modern horses from the territory of Poland and Romania, respectively. PKT, PN, G — camels of Eurasia with geological age, respectively, Pleistocene (Calabrian-Chibanian), Late Pleistocene, Holocene. Landscapes are given according to G. Bocheren (Bocherences, 2003; Bocherences et al., 2003). C 3 , CAM, C 4 — types of photosynthesis within the Calvin—Benson—Bassam cycle (Bassham et al., 1950) in sequence from plants of forest-steppe environments (C 3 ) to plants of steppes and savannahs (CAM) and further to desert plants (C 4 )
реконструируется жаркий сухой климат в условиях саванно-пустынных обстановок. Плиоценовые верблюды Paracamelus alexejevi и P . praebactrianus (Одесса, Западная Сибирь, Западная Монголия) обитали в условиях перехода от жаркого относительно сухого климата к теплому и относительно влажному климату саванн. Верблюды P. alutensis и P. trofimovi начала плейстоцена ( гелазий; MIS 103-63) (Ростовская область, Таджикистан) существовали в условиях умеренно-жаркого и умеренно-влажного климата в саванно-степной ландшафтной обстановке. В раннем плейстоцене ( калабрий; MIS 62-20) условия проживания верблюдов P. gigas из Крыма отвечали климатическому оптимуму для саванн-степей. Ранненеоплейстоценовые верблюды Camelus knoblochi возрастом 120—80 тыс. лет (Поволжье, Забайкалье) обитали в условиях теплого влажного климата предледниковья в широком диапазоне ландшафтных обстановок от саванн до лесостепей и, возможно, лесов. Верблюды Camelus cf . ferus позднего плейстоцена (Астраханская область) проживали во время ленинградского (средневалдайского) мегаинтерстадиала (40—45 тыс. л., MIS-3), когда наступило потепление и развились лесотундровые ландшафты. Как раз для этого интерстадиального периода отмечают смягчение климата и увеличение площади лесного покрова (Ташак, Антонова, 2015). Голоценовые (MIS-1) верблюды Camelus bactrianus из археологических памятников Краснодарского края и Ростовской области обитали в условиях широких климатических вариаций от прохладного предоптимума к римскому оптимуму и далее к похолоданию в раннем средневековье (IV—VII века) и к средневековому оптимуму (X— XIII века). Очевидно, что полученные экспериментальные данные и сделанные выводы подтверждают ранее 26
высказанные предположения о широком распространении древних верблюдов в диапазоне от пустынных до лесостепных ландшафтов (Титов, 2004; Titov, Logvinenko, 2006; Titov, 2008).
В связи с реконструкциями условий обитания евразийских верблюдов большой интерес представляют аналогичные данные по плейстоценовым верблюдам, проживавшим на территории современных США. Судя по изотопным данным (Yann et al., 2016), североамериканские верблюды Camelops, Hemiauchenia и Palaeolama проживали в гораздо более жарких саванно-пустынных обстановках (поле США на рис. 23), что вполне совпадает с общими ландшафтно-климатическими реконструкциями для этой территории.
Изотопный состав стронция — важнейший индикатор миграционной активности животных (Силаев и др., 2021). Суть в том, что кости животных, проживающих на предгорных территориях, содержат изначально больше стронция с изотопным составом (87Sr/86Sr) вблизи показателей морской воды и морских карбонатов — 0.706—0.709. Кости животных, проживающих в условиях континентальных низменностей, отличаются более низким содержанием стронция и его изотопным составом, сближающимся с аналогичными показателями подземных и речных вод — 0.707—0.712 (Фролова, Бурикова, 1997; Юдович, 2007; Алексеева и др., 2014; Киселева и др., 2022). Проведенные исследования показали (табл. 9), что исследованные образцы по содержанию и изотопному составу стронция могут быть подразделены на две группы (рис. 24).
В первую группу входят кости плиоценовых верблюдов из Одесских катакомб, юга Западной Сибири, Монголии и голоценовых из Краснодарского края. Для этих костей характерны относительно низкие содер-
Таблица 9 . Содержание и изотопный состав стронция в исследованных образцах ископаемых верблюдов на территории Северной Евразии
Table 9. Content and isotopic composition of strontium in the studied samples of fossil camels in the territory of Northern Eurasia
|
№ п/п No. |
№ обр. Sample No. |
Геологический возраст Geological age |
Место нахождения Location |
Sr, г/т Sr, g/t |
87Sr/86Sr |
|
1 |
ВРБ-1 |
поздний миоцен / Late Miocene |
Ростовская область / Rostov region |
400 |
0.708628 |
|
2 |
» |
» |
» |
1200 |
0.708786 |
|
3 |
» |
» |
» |
300 |
0.708448 |
|
4 |
ТИТ-3 |
плиоцен / Pliocene |
» |
1500 |
0.709423 |
|
5 |
ВРБ-3 |
» |
Одесса / Odessa |
1400 |
0.709711 |
|
6 |
ВРБ-2 |
» |
Западная Сибирь / Western Siberia |
1400 |
0.710291 |
|
7 |
ВРБ-4 |
» |
Западная Монголия / Western Mongolia |
1200 |
0.709444 |
|
8 |
ТИТ-1 |
плейстоцен (гелазийский век) Pleistocene (Gelasian) |
Ростовская область / Rostov region |
1600 |
0.709449 |
|
9 |
ТИТ-2 |
» |
» |
1300 |
0.709011 |
|
10 |
ВРБ-10 |
» |
» |
1300 |
0.709428 |
|
11 |
ВРБ-5 |
» |
Таджикистан / Tajikistan |
2100 |
0.709329 |
|
12 |
ВРБ-7 |
плейстоцен (гелазийский век) Pleistocene (Gelasian) |
Ростовская область / Rostov region |
1700 |
0.709519 |
|
13 |
ВРБ-9 |
голоцен / Holocene |
Забайкалье / Transbaikalia |
900 |
0.707464 |
|
14 |
ВРБ-10 |
» |
РФ/Монголия / RF/Mongolia |
900 |
0.709925 |
|
15 |
ВРБ-8 |
» |
Поволжье / Volga region |
700 |
0.708462 |
|
16 |
ТИТ-4 |
» |
Ростовская область /Rostov region |
500 |
0.709158 |
|
17 |
ТИТ-5 |
» |
Краснодарский край / Krasnodar region |
1100 |
0.7096.46 |
|
18 |
ТИТ-6 |
» |
Ростовская область / Rostov region |
600 |
0.709136 |
|
19 |
ТИТ-7 |
» |
Ростовская область / Rostov region |
600 |
0.709322 |
|
Верблюды-аборигены / Aboriginal camels |
933 ± 488 (52 %) |
0.709298 ± 0.000772 (11 %) |
|||
|
Верблюды-мигранты / Migrating camels |
1167 ± 510 (44 %) |
0.709095 ± 0.00059 (8 %) |
|||
87Sr/86Sr
0.7078
0.7102-
0.7100 —
0.7098 —
0.7096 —
0.7094 —
0.7092 —
0.7090-
0.7088-
0.7086 -
0.7084 -
0.7082 -
0.7080 -

800 1200
Sr, г/т
Рис. 24. Стронций в костях ископаемых верблюдов как критерий различия животных —«аборигенов» и «мигрантов». Расшифровка номеров образцов в табл. 7
Fig. 24 . Strontium in the bones of fossil camels as a criterion for distinguishing between «aboriginal» and «migrant» animals. Explanation of sample numbers in table. 7
жания стронция (в среднем ниже 1000 г/т), но относительно высокие значения стронциевого изотопного коэффициента. Частота встречаемости таких образцов составляет около 21 %. Можно предполагать, что верблюды с такими параметрами по стронцию были в отношении соответствующих территорий животными-аборигенами (автохтонными), проживавшими в условиях саванн-степей в периоды более или менее стабильного теплого и умеренно влажного климата.
Вторую группу составляют резко преобладающие по частоте встречаемости кости с возрастом от позднего миоцена до голоцена, отобранные на территории от Краснодарского края и Ростовской области до Забайкалья и Западной Монголии. Эти образцы отличаются более высоким содержанием стронция (в среднем выше 1000 г/т), но относительно низким значением стронциевого изотопного коэффициента. Такие верблюды могли быть животными-мигрантами, проживавшими, судя по С-О-N-изотопным данным, во времена широких колебаний климата и широком диапазоне ландшафтных обстановок — от полупустынь до саванн-степей и лесостепей.
Заключение
Исследована уникальная серия проб костных остатков ископаемых верблюдов, отобранных из местонахождений на территории Евразии от Северо-Западного Причерноморья до Западного Забайкалья и Монголии, датированных в хронологическом диапазоне от позднего миоцена (6 млн л.) до XIV века н. э. В ходе иссле- дований применялся широкий комплекс современных методов: оптическая микроскопия, термический, химико-аналитический, рентгенофлюоресцентный, рентгенодифракционный и ретгеноспектральный микро-зондовый анализы, масс-спектрометрия с индуктивно связанной плазмой; инфракрасная и рамановская спектроскопия; изотопная масс-спектрометрия (С, О, N, Sr).
Согласно полученным результатам, образцы костей верблюдов голоценового возраста характеризуются хорошей сохранностью как компактной, так и губчатой типов костной ткани. В более древних — плио-цен-плейстоценовых — костях плотная и губчатая костные ткани явно деградированы с образованием каверн и трещин, часто заполненных новообразованными минералами, особенно карбонатами и оксигидроксидами железа. Биоапатит в таких костях подвержен коллофанизации с потерей прозрачности и приобретением на таких участках буроватой окраски. По состоянию нанопористости верблюжьи кости ближе всего согласуются с костями плейстоценовой мамонтовой фауны. При этом кости верблюдов поздне-миоцен-плейстоценового возраста системно отличаются от костей голоценовых верблюдов как большим совокупным объемом нанопор, так и пропорционально большим их условным числом (lgNo). Это отражает хронологическую тенденцию наноструктурной деградации костных остатков верблюдов в ходе фосси-лизации.
В химическом составе исследованных костей содержание неапатитовых примесей колеблется в довольно узких пределах — от 3.5 до 8.23 мас. %, возрастая по мере удревнения костей. В целом это в 1.2—3.0 раза уступает костям ископаемых млекопитающих с территорий Печорского Приуралья, Пермской области (стоянка Заозерье), Северо-Восточного Казахстана (стоянка Ушбулак). Атомный модуль Ca/P, характеризующий биоапатитовую компоненту, свидетельствует об аномально высокой степени кальцитизации верблюжьих костей, возрастающей от голоценовых образцов к позднемиоценовым. По данным рентгенофазового и рентгеноспектрального микрозондового анализов, в исследованных костях присутствует терригенная микропримесь кварца, альбита, ортоклаза, циркона, ильменита, гидрослюд и хлоритов. Кроме того, в верблюжьих костях выявлен ряд аутигенно-эпигенетических минералов: кальцита, поликомпонентного родохрозита, барита, барито-ангидрита, пирита, гётита и манганита. Степень обогащения эпигенетическими минералами возрастает по мере удревнения костей.
В составе исследуемых костных остатков обнаружен 51 микроэлемент, в том числе 11 элементов-эс-сенциалов, 18 физиологически активных элементов и 22 элемента-антибионта. Суммарная концентрация микроэлементов изменяется в диапазоне от 1500 до 5000 г/т, возрастая в 2—3 раза по мере удревнения костей. Отношение групповых концентраций микроэле-ментов-эссенциалов к микроэлементам-антибионтам снижается в среднем от 15.35 в голоценовых костях до 0.93 в плейстоценовых. Эту закономерность отражает и отношение содержаний эссенциального цинка и физиогенно-активной меди, которое в той же хронологической последовательности изменяется в среднем от 26.09 до 7.83. Таким образом, устанавливается сильная хронологическая тенденция замещения сингенетичных микроэлементов ксеногенными по мере фоссилизации верблюжьих костей. По суммарному содержанию микроэлементов исследованные верблюжьи кости могут быть сопоставлены только с костями млекопитающих со стоянки Ушбулак. Все другие изученные нами костные остатки характеризовались многократно большим обогащением микроэлементами.
Степень и тенденции изменения кристалличности биоапатита в верблюжьих костях определялись рентгеноструктурным и ИК-спектроскопическим методами. Из полученных данных следует, что в исследованной коллекции реализуется сильная тенденция увеличения степени кристалличности биоапатита в направлении от молодых костей к древним как следствие прогрессирующей фоссилизации. В составе биоапатита выявлены структурные примеси Sr и Mn в катионной и Si, S, C в анионной подрешетках. При этом биоапатит в голоценовых и плейстоцен-калабрийских костях практически не содержит примесей в катионной подрешетке и минимально содержит примеси кремния и серы в анионной подрешетке. В гелазийско-ми-оценовых костях биоапатит гораздо более примесный в обеих подрешетках. Таким образом, в биоапатите верблюжьих костей явно реализуется хронологическая тенденция усложнения состава на фоне сокращения степени карбонизации.
На кривых нагревания в диапазоне 20—600 °С зарегистрированы все три характерных для костей термических эффекта, степень проявления которых обратно коррелируется с геологическим возрастом образцов: у костей голоценового возраста они реализуются гораздо лучше, чем у более древних. Очевидно, что это обусловлено резким сокращением содержания коллагена по мере удревнения костей. По данным СЭМ-исследований выделенного из костей коллагена, его микростроение тоже сильно зависит от возраста костей. В коллагене костей голоценового возраста хорошо сохраняется первичная плотность и волокнистость без явных признаков деструкции. В более древних костях первичная волокнистость в коллагене замещается агрегатом разориентированных фрагментов волокон, часто в комбинации с микроглобулярными бактериальными формами пирита. Степень деградации органического матрикса в ископаемых костях хорошо отражают спектры КР. В случае голоценовых костей регистрируется сильная люминесценция с весьма незначительным проявлением рамановских линий. В спектрах КР, полученных из более древних костей, доминируют рамановские линии G и D, указывающие на присутствие сильно преобразованного (углеродизи-рованного) органического вещества.
Содержание углерода и азота в органической части коллагена колеблется в пределах соответственно 26—32 и 10.2—11.66 мас. %, снижаясь от голоценовых костей к палеоплейстоценовым. Атомное отношение С/N изменяется в рамках 3.1—3.83. Содержания неорганических примесей в костном коллагене, особенно карбонатных и гидроксижелезистых, возрастает от молодых костей к древним в 15—300 раз.
В составе костного коллагена ископаемых верблюдов обнаружены 15 аминокислот, представляющих обе серии (незаменимые и заменимые) и шесть групп — алифатические (А), основные (О), кислые (К), гидрок- сильные (Г), имино (И), серусодержащие (СС). Общее содержание аминокислот колеблется в пределах 700— 915 мг/г, не коррелируясь с геологическим возрастом костей. Пропорция между незаменимыми и заменимыми кислотами колеблется в узких пределах от 0.17 до 0.26, тоже не обнаруживая корреляции с геологическим возрастом верблюдов. В целом в исследованной коллекции ископаемых костей реализуется универсальный ряд снижения содержания групп аминокислот в направлении: алифатические > имино > кислые > гидроксильные > ароматические > основные > серу-содержащие.
Согласно полученным данным, исследованные нами ископаемые верблюды на фоне других плейстоцен-голоценовых млекопитающих, обитавших на территории Европы и севере Евразии, характеризуются аномальными изотопными свойствами как по биопати-ту, так и костному коллагену. Это выражается в том, что биоапатит в костных остатках верблюдов демонстрирует сочетание относительно изотопно-тяжелого углерода (–11…–4 ‰) и изотопно-тяжелого кислорода (20—26 ‰). По коллагену верблюды отличаются аномально тяжелым азотом (7—14 ‰) и большей частью изотопно-утяжеленным углеродом (–22…–14 ‰). Все эти особенности могут указывать на обитание верблюдов, в отличие других ранее исследованных нами плей-стоцен-голоценовых животных, в гораздо более теплых и при этом широко варьирующихся климатических условиях.
Результаты исследований содержания и изотопного состава стронция показали, что исследованные образцы по этим признакам могут быть подразделены на две группы. В первую группу входят кости верблюдов плиоценового возраста из Одесских катакомб, с юга Западной Сибири, из Монголии и голоценовых из Краснодарского края, для которых характерны относительно низкие содержания стронция (в среднем ниже 1000 г/т), но сравнительно высокие значения стронциевого изотопного коэффициента. Можно предполагать, что верблюды с такими параметрами были автохтонными животными, проживавшими в условиях саванн-степей в периоды более или менее стабильного теплого и умеренно влажного климата. Вторую группу составляют резко преобладающие по частоте встречаемости кости с возрастом от позднего миоцена до голоцена, отобранные на территории от Краснодарского края и Ростовской области до Забайкалья и Западной Монголии. Эти образцы отличаются более высоким содержанием стронция (в среднем выше 1000 г/т), но относительно низким значением стронциевого изотопного коэффициента. Соответственно, верблюды с такими показателями могли быть аллохтонными животными, обитавшими в условиях колебаний климата и мигрировавшими в широком диапазоне ландшафтных обстановок — от полупустынь до саванн-степей и лесостепей.
Все полученные результаты указывают на минералого-геохимические свойства ископаемых костей как на весьма эффективные средства эколого-климатических реконструкций и расшифровки истории эволюции млекопитающих.
Работа выполнена в рамках официальной научной темы 0333-2022-0044 «Фундаментальные проблемы минералогии и минералообразования, минералы как инди- каторы петро- и рудогенеза, минералогия рудных районов и месторождений Тимано-Североуральского региона и арктических территорий» и гранта РНФ № 22-2700450 (палеонтологическая часть, В. В. Титов).
Авторы благодарят проф. Й. Плихта (Нидерланды) за получение первых данных по изотопии ископаемых костей евразийских верблюдов, м. н. с. Е. М. Тропникова и ст. инженера-технолога С. Т. Неверова за участие в исследованиях.
Список литературы Ископаемые кости миоцен-голоценовых верблюдов Евразии (Россия): минералого-геохимические свойства и экологические реконструкции
- Алексеева Л. И. Верблюды Северного Причерноморья и Предкавказья // Млекопитающие позднего кайнозоя юго-запада СССР. Кишинев: Штиинца, 1974. C. 53—76.
- Alekseeva L. I. Camels of the Northern Black Sea region and Ciscaucasia. Mammals of the Late Cenozoic of the southwest of the USSR. Chisinau: Shtiintsa, 1974, pp. 53—76. (in Russian)
- Алексеева Л. П., Алексеев С. В., Кононов А. М. Изотопные стронциевые отношения (87Sr/86Sr) в подземных льдах и водах Оленекского артезианского бассейна (Западная Якутия) // Известия Сибирского отделения Секции наук о Земле РАЕН. 2014. № 6. С. 97—103.
- Alekseeva L. P., Alekseev S. V., Kononov A. M. Strontium isotope ratios (87Sr/86Sr) in underground ice and waters of the Olenek artesian basin (Western Yakutia). News of the Siberian Branch of the Section of Earth Sciences of the Russian Academy of Natural Sciences. 2014, No. 6, pp. 97—103. (in Russian)
- Анищенко Л. А., Шанина С. Н. Аминокислоты в недрах: результаты исследования и перспективы изучения // Вестник Института геологии Коми НЦ УрО РАН. 2010. № 9. С. 34—35.
- Anishchenko L. A., Shanina S. N. Amino acids as possible indicators of the conditions of accumulation and conservation of organic matter. Organic mineralogy: Proc. of the Russian meeting. Syktyvkar: Geoprint, 2009, pp. 71—74. (in Russian)
- Анищенко Л. А., Шанина С. Н. Аминокислоты как возможные индикаторы условий накопления и консервации органического вещества // Органическая минералогия: Материалы росс. совещания. Сыктывкар: Геопринт. 2009. С. 71—74.
- Anishchenko L. A., Shanina S. N. Sulfur in amino acids. Structure and diversity of the mineral world: Proc. of the International Mineralogical Seminar. Syktyvkar: Geoprint, 2008, pp. 302—304. (in Russian)
- Анищенко Л. А., Шанина С. Н. Сера в аминокислотах // Структура и разнообразие минерального мира: Материалы междунар. минералог. семинара. Сыктывкар: Геопринт. 2008. С. 302—304.
- Anishchenko L. A., Shanina S. N. Sulfur in amino acids. Structure and diversity of the mineral world: Proc. of the International Mineralogical Seminar. Syktyvkar: Geoprint, 2008, pp. 302—304. (in Russian)
- Аубекерова П. А. Новый гигантский верблюд из плиоценовых отложений текесской впадины // Фауна и флора из мезокайнозоя Южного Казахстана. Алма-Ата: Наука, 1974. Т. 6. C. 74—82 («Gigantocamelus» longipes Aubekerova, 1974).
- Aubekerova P. A. New giant camel from Pliocene deposits of the Tekes depression. Fauna and flora from the Meso-Cenozoic of Southern Kazakhstan. Alma-Ata: Nauka, 1974, V. 6, pp. 74—82 (“Gigantocamelus” longipes Aubekerova, 1974). (in Russian)
- Каткова В. И. Шанина С. Н. D-аминокислоты в составе биоапатита // Органическая минералогия: Материалы минералог. семинара с междунар. участием. Сыктывкар: ИГ Коми НЦ УрО РАН, 2019. С. 45—46.
- Katkova V. I. Shanina S. N. D-amino acids in the composition of bioapatite. Organic mineralogy: Proc. of a mineralogical seminar with international participation. Syktyvkar: IG Komi SC UB RAS, 2019, pp. 45—46. (in Russian)
- Киселева Д. В., Шагалов Е. С., Окунева Т. Г., Солошенко Н. Г., Рянская А. Д., Панкрушина Е. А., Карпова С. В., Уразова К. К., Сидурук А. Р. Изотопные отношения стронция 87Sr/86Sr в воде ряда рек Южного Урала // Geodynamics and Tectonophysics, 2022. V. 13. № 12. https: doi.org/10.5800/GT-2022-13-2s-0602.
- Kiseleva D. V., Shagalov E. S., Okuneva T. G., Soloshenko N. G., Ryanskaya A. D., Pankrushina E. A., Karpova S. V., Urazova K. K., Siduruk A. R. Isotopic ratios of strontium 87Sr/86Sr in the water of a number of rivers of the Southern Urals. Geodynamics and Tectonophysics, 2022, V. 13, No. 12. https: doi.org/10.5800/GT-2022-13-2s-0602. (in Russian)
- Котельникова Е. Н. Особенности проявления изомерии, хиральности, четности и упаковки молекул в кристаллической структуре природных органических веществ // Современные проблемы теоретической, экспериментальной и прикладной минералогии (Юшкинские чтения — 2014). Сыктывкар: Геопринт, 2014. С. 201— 208.
- Kotelnikova E. N. Features of the manifestation of isomerism, chirality, parity and packing of molecules in the crystal structure of natural organic substances. Modern problems of theoretical, experimental and applied mineralogy (Yushkin Readings 2014). Syktyvkar: Geoprint, 2014, pp. 201—208. (in Russian)
- Майдль Т. В., Филиппов В. Н. Фрамбоидальный пирит в фоссилизированных костных остатках позвоночных покаямской свиты девона Северного Тимана // Современные проблемы теоретической, экспериментальной и прикладной минералогии (Юшкинские чтения — 2018): Материалы минералог. Семинара с междунар. участием. Сыктывкар: ИГ Коми НЦ УрО РАН. 2018. С. 141—142.
- Maidl T.V., Filippov V.N. Framboidal pyrite in fossilized bone remains of vertebrates of the Pokayama Formation of the Devonian of Northern Timan. Modern problems of theoretical, experimental and applied mineralogy (Yushkin Readings 2018): Proceedings of a mineralogical seminar with international participation. Syktyvkar: IG Komi SC UB RAS, 2018, pp. 141—142. (in Russian)
- Свистун В. И. Новые находки остатков верблюдов (TYLOPODA CAMELIDAE) в области Понта юга европейской части СССР // Вестник зоологии. 1971. № 1. С. 64—68.
- Svistun V. I. New finds of remains of camels (TYLOPODA CAMELIDAE) in the Pontus region of the south of the European part of the USSR Bulletin of Zoology, 1971, No. 1, pp. 64—68. (in Russian)
- Силаев В. И., Паршукова М. Н., Гимранов Д. О., Филиппов В. Н., Киселёва Д. В., Смолева И. В., Тропников Е. М., Хазов А. Ф. Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал) // Вестник Пермского университета. Геология. 2020. Т. 19. № 4. С. 323—358.
- Silaev V. I., Parshukova M. N., Gimranov D. O., Filippov V. N., Kiseleva D. V., Smoleva I. V., Tropnikov E. M., Khazov A. F. Mineralogical and geochemical Features of cave fossilization of fossil bones using the example of the Imanai cave (Southern Urals). Bulletin of Perm University. Geology, 2020, V. 19, No. 4, pp. 323—358. (in Russian)
- Силаев В. И., Смолева И. В., Филиппов В. Н., Шанина М. Н., Хазов А. Ф., Макеев Б. А., Киселева Д. В., Фокина А. К., ван дер Плихт Й. Плейстоценовый морж на р. Печоре: минералого-геохимические данные и палеоэкологические реконструкции // Известия Коми НЦ УрО РАН. Серия «Науки о Земле». 2023. № 2. С. 56—66.
- Silaev V. I., Smoleva I. V., Filippov V. N., Shanina M. N., Khazov A. F., Makeev B. A., Kiseleva D. V., Fokina A. K., J van der Plicht Pleistocene walrus on the river. Pechora: mineralogical and geochemical data and paleoecological reconstructions. Proceedings of the Komi Scientific Center of the Ural Branch of the Russian Academy of Sciences. Series «Earth Sciences», 2023, No. 2, pp. 56—66. (in Russian)
- Силаев В. И., Шанина С. Н., Смолева И. В., Киселёва Д. В., Чередниченко Н. В., Хазов А. Ф., Туманова Е. А., Павлов П. Ю. Опыт использования минералого-геохимических свойств костных остатков для реконструкции среды обитания на палеолитической стоянке Заозерье (Средний Урал) // ПАЖМИ. 2019. № 2. С. 35—77.
- Silaev V. I., Shanina S. N., Smoleva I. V., Kiseleva D. V., Cherednichenko N. V., Khazov A. F., Tumanova E. A., Pavlov P. Yu. Experience in using mineral –geochemical properties of bone remains for the reconstruction of the habitat at the Paleolithic site of Zaozerye (Middle Urals), PAJIR, 2019, No. 2, pp. 35—77. (in Russian)
- Силаев В. И., Анойкин А. А., Павлёнок Г. Д., Хазов А. Ф., Киселёва Д. В., Шанина С. Н., Макеев Б. А., Уразова К. К. Костные остатки млекопитающих с палеонтологической стоянки Ушбулак (Северо-Восточный Казахстан): археологический контекст, минералого-геохимические свойства и палеоэкологические реконструкции // Вестник геонаук. 2022. № 4. С. 3—37.
- Silaev V. I., Anoykin A. A., Pavlenok G. D., Khazov A. F., Kiseleva D. V., Shanina S. N., Makeev B. A., Urazova K. K. Bone remains of mammals from the paleontological site of Ushbulak (North-Eastern Kazakhstan): archaeological context, mineralogical and geochemical properties and paleoecological reconstructions. Vestnik of Geosciences, 2022, No. 4, pp. 3—37. (in Russian)
- Силаев В. И., Косинцев П. А., Филиппов В. Н., Киселева Д. А., Смолева И. В., Солошенко Н. Г., Окунева Т. Г., Чередниченко Н. В., Тропников Е. М., Хазов А. Ф. Байгаринский человек эпохи мезолита: минералого- геохимические исследования, палеоэкологические реконструкции и история ранних миграций человека на территорию Западной Сибири // Вестник геонаук. 2021. № 5. С. 5—8.
- Silaev V. I., Kosintsev P. A., Filippov V. N., Kiseleva D. A., Smoleva I. V., Soloshenko N. G., Okuneva T. G., Cherednichenko N. V., Tropnikov E. M., Khazov A.F. Baigarinsky man of the Mesolithic era: mineralogical and geochemical studies, paleoecological reconstructions and the history of early human migrations to the territory of Western Siberia. Vestnik of Geosciences, 2021, No. 5, pp. 5—8. (in Russian)
- Силаев В. И., Пономарев Д. В., Симакова Ю. С., Шанина С. Н., Смолева И. В., Тропников Е. М., Хазов А. Ф. Современные исследования ископаемого костного детрита: пале- онтология, минералогия, геохимия // Вестник Института геологии Коми НЦ УрО РАН. 2016. № 5. С. 19—31.
- Silaev V. I., Ponomarev D. V., Simakova Yu. S., Shanina S. N., Smoleva I. V., Tropnikov E. M., Khazov A. F. Modern studies of fossil bone detritus: paleontology, mineralogy, geochemistry. Vestnik IG Komi SC UB RAS, 2016, No. 5, pp. 19—31. (in Russian)
- Смирнов В. И., Вотяков С. Л., Садыкова Н. О., Киселёва Д. В., Щапова Ю. В. Физико-химические характеристики ископаемых костных остатков млекопитающих и проблема оценки их относительного возраста // Термический и масс-спектрометрический элементный анализ. Екатеринбург: Гощицкий, 2009. 118 с.
- Smirnov V. I., Votyakov S. L., Sadykova N. O., Kiseleva D. V., Shchapova Yu. V. Physicochemical characteristics of fossil bone remains of mammals and the problem of estimating their relative age. Thermal and mass spectrometric elemental analysis. Ekaterinburg: Goschitsky, 2009, 118 p. (in Russian)
- Ташак В. И., Антонова Ю. Е. Модель поведения населения эпохи палеолита Западного Забайкалья при выборе мест стоянок (по материалам Подзвонкой) // Фундаментальные проблемы квартера, итоги изучения и основные направления исследований: Материалы IX Всеросс. совещ. по изуч. четвертичного периода. Иркутск: Изд-во географии СО РАН, 2015. С. 455—457.
- Tashak V. I., Antonova Yu. E. Model of behavior of the population of the Paleolithic era of Western Transbaikalia when choosing sites (based on materials from Podzvonka). Fundamental problems of the Quaternary, results of study and main directions of research: Proc. of the IX All-Russian meeting on the study of the Quaternary period. Irkutsk: Geography Publishing House SB RAS. 2015. pp. 455—457. (in Russian)
- Титов В. В. Остатки верблюдов из золотоордынских археологических памятников Приазовья // Диалог городской и степной культур на евразийском пространстве. Донские древности. Вып. 10. Азов, 2009. С. 472—475.
- Titov V.V. Remains of camels from the Golden Horde archaeological monuments from the Azov region. Dialogue of urban and steppe cultures in the Eurasian space. Don antiquities. V. 10, Azov, 2009, pp. 472—475. (in Russian)
- Титов В. В., Головачёв М. В. Остатки скелета ископаемого верблюда Camelus cf. bactrianus из верхненеоплейсто-ценовых отложений Нижней Волги // Биогеография и эволюционные процессы: Материалы LXVI сессии Палеонтологического общества при РАН. СПб.: Картфабрика ВСЕГЕИ, 2020. С. 276—278.
- Titov V.V., Golovachev M.V. Skeletal remains of the fossil camel Camelus cf. bactrianus from the Upper Neopleistocene deposits of the Lower Volga. Biogeography and evolutionary processes. Proc. of the LXVI session of the Paleontological Society of the Russian Academy of Sciences. St. Petersburg: VSEGEI Map Factory. 2020. pp. 276—278. (in Russian)
- Фролова Т. И., Бурикова И. А. Магматические формации современных океанических обстановок. М.: МГУ, 1997. 320 с.
- Frolova T. I., Burikova I. A. Magmatic formations of modern oceanic environments. M.: Moscow State University, 1997, 320 p. (in Russian)
- Хавесон Я. И. Третичные верблюды восточного полушария (род Paracamelus) // Тр. ПИН АН СССР. 1954. Т. 47. C. 100—161.
- Haveson Ya. I. Tertiary camels of the eastern hemisphere (genus Paracamelus) // Tr. PIN AS USSR, 1954, V. 47, pp. 100—161. (in Russian)
- Шанина С. Н., Голубев Е. А. Аминокислоты в шунгитах Карелии // Геохимия. 2010. № 9. С. 972—987.
- Shanina S.N., Golubev E.A. Amino acids in shungites of Karelia. Geochemistry, 2010, No. 9, pp. 972—987. (in Russian)
- Юдович Я. Э. Нормировка изотопного состава Sr в морской воде: удобный методический прием? // Вестник ИГ Коми НЦ УрО РАН. 2007. № 1. С. 9—10.
- Yudovich Ya. E. Normalization of the isotopic composition of Sr in sea water: a convenient methodological technique? Vestnik IG Komi SC UB RAS, 2007, No. 1, pp. 9—10. (in Russian)
- Bassham J., Benson A., Calvin M. The path of carbon in Photosynthesis // J. Biol. Chem., 1950. № 196 (2). Р. 703—716.
- Bocherences H. Isotopic Biogeochemistry and the paleoecology of the mammoth Steppe Fauna // Advances in Mammoth Research, 2003. № 9. P. 57—76.
- Bocherences H., Drucker D. Trophic Lovel Isotopic Enrichment of Carbon and Nitrogen in Bone Collagen: Gase Studies from Recent an Ancient Terrestrial Ecosystems // Intern. J. Osteoarchaelogy, 2003. V. 13. P. 46—53.
- Connin S. L., Betancourt J., Quade J. Late Pleistocene C4 plant dominance and summer rainfall in the southwestern United States from isotopic study of herbivore teeth // Quat. Res., 1998. V. 50. P. 179—193.
- Domingo L., Prado J. L., Alberdi M. T. The effect of paleoecology and paleobiogeography on stable isotopes of Quaternary mammals from South America // Quat. Sci. Rev., 2012. V.55. P. 103—113.
- Feranec R. S. Stable isotopes, hypsodonty, and the paleodiet of Hemiauchenia (Mammalia: Camelidae): a morphological specialization creating ecological generalization // Paleobiology, 2003. V. 29. № 2. P. 230—242.
- Feranec R. S., MacFadden B. J. Evolution of the grazing niche in Pleistocene mammals from Florida: evidence from stable isotopes // Palaeogeography, Palaeoclimatology, Palaeoecology, 2000. V. 162. P. 155—169.
- Feranec R. S., MacFadden B. J. Evolution of the grazing niche in Pleistocene mammals from Florida: evidence from stable isotopes // Palaeogeogr. Palaeoclimatol. Palaeoecol, 2000. V. 162, № 1—2. P. 155—169.
- Feranec R.S., MacFadden B.J. Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California // Paleobiology. 2006. Vol. 32. P. 191—205.
- Geraads D. Camelus grattardi, sp. nov., a new camel from the Shungura Formation, Omo Valley, Ethiopia, and the relationships of African fossil Camelidae (Mammalia) // Journal of Vertebrate Paleontology, 2014. V. 34. № 6. P. 1481—1485. DOI: 10.1080/02724634.2014.880447.
- Geraads D., Barr W.A., Reed D., Laurin M., Alemseged Z. New Remains of Camelus grattardi (Mammalia, Camelidae) from the Plio-Pleistocene of Ethiopia and the Phylogeny of the Genus // Journal of Mammalian Evolution. 2021. Vol. 28. P. 359—370. DOI: 10.1007/s10914-019-09489-2.
- Geraads D., Didier G., Barr, W. A. Reed, D., and Laurin M. The fossil record of camelids demonstrates a late divergence between Bactrian camel and dromedary // Acta Palaeontologica Polonica, 2020. V. 65, № 2. P. 251—260. DOI: 10.4202/app.00727.2020.
- Harris J. M., Geraads D., Solounias N. Camelidae. In: Cenozoic mammals of Africa. L. Werdelin & W.J. Sanders (Eds.). University of California Press, 2010. P. 815—820. DOI: 10.1525/california/9780520257214.003.0041.
- Higgins P., MacFadden B. J. Seasonal and geographic climate variabilities during the Last Glacial Maximum in North America: applying isotopic analysis and macrophysical climate models // Palaeogeogr. Palaeoclimatol. Palaeoecol, 2009. V. 283, № 1—2. P. 15—27.
- Hoppe K. A., Koch P. L. The biogeochemistry of the Aucilla River fauna. First Floridians and Last Mastodons: The Page-Ladson Site in the Aucilla River. Springer, 2006. P. 379—401.
- Kita Z. A., Secord R., Boardman G. S. A new stable isotope record of Neogene paleoenvironments and mammalian paleoecologies in the western Great Plains during the expansion of C4 grasslands // Palaeogeogr. Palaeoclimatol. Palaeoecol, 2014. V. 399. P. 160—172.
- Kohn M. J., McKay M. P. Paleoecology of late Pleistocene–Holocene faunas of eastern and central Wyoming, USA, with implications for LGM climate models // Palaeogeogr. Palaeoclimatol. Palaeoecol, 2012. V. 326. P. 42—53.
- Kohn M. J., McKay M. P., Knight J. L. Dining in the Pleistocene— who's on the menu? // Geology, 2005. V. 33, № 8. P. 649—652.
- Likius A., Brunet M., Geraads D.,Vignaud P. Le plus vieux Camelidae (Mammalia, Artiodactyla) d'Afrique limite Mio-Pliocène, Tchad // Bulletin de la Société géologique de France, 2003. T. 174. № 2. P. 187—193.
- Liu W., Hou S., Zhang X. Revision of the Late Cenozoic camelids from the Yushe Basin, Shanxi, with comments on Chinese fossil camels // Quaternary Sciences, 2023. V. 43. № 3. P. 712—751. DOI: 10.11928/j.issn.1001-7410.2023.03.05.
- MacFadden B. J., Cerling T. E. Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: a 10 million-year sequence from the Neogene of Florida // J. Vertebr. Paleontol., 1996. V.16. P. 103—115.
- MacFadden B. J., Shockey B. J. Ancient feeding ecology and niche differentiation of Pleistocene mammalian herbivores from Tarija, Bolivia: morphological and isotopic evidence // Paleobiology, 1997. V. 23, № 1. P. 77—100.
- Made van der J., Morales J. Family Camelidae. In: The Miocene land mammals of Europe / G.E. Rössner and K. Heissig (eds.). München: Pfeil, 1999. P. 221—224.
- Martini P., Costeur L., Le Tensorer J.-M., Schmid P. Pleistocene camelids from the Syrian Desert: The diversity in El Kowm L’anthropologie. 2015. Vol. 119. P. 687—693. DOI: 10.1016/j.anthro.2015.10.005.
- Martini P., Geraads D. Camelus thomasi Pomel, 1893, from the Pleistocene type-locality Tighennif (Algeria). Comparisons with modern Camelus // Geodiversitas, 2018. V. 40, № 5. P. 115—134. DOI:10.5252/geodiversitas2018v40a5.
- Nunez E. E., MacFadden B. J., Mead J. I., Baez A. Ancient forests and grasslands in the desert: diet and habitat of Late Pleistocenemammals from northcentral Sonora, Mexico // Palaeogeogr. Palaeoclimatol. Palaeoecol, 2010. V. 297. P. 391—400.
- Orlov Ju. A. Über die Reste der fossilen Camelidae aus dem Gouvernment Semipalatinsk (Westsiberien) // Ann. Soc. Pal. Russie, 1930. № 8. S. 99—116.
- Orlov Ju. A. Über die Reste der fossilen Cameliden aus dem Gouvernement Akmolinsk (Westsiberien) // Ann. Mus. Zool. Acad. Sci. URSS, 1929. № 30. S. 549—587.
- Pérez-Crespo V. A., Arroyo-Cabrales J., Alva-Valdivia L. M., Morales-Puente P., Cienfuegos-Alvarado E. Datos isotópicos (δ13C, δ18O) de la fauna pleistocenica de la Laguna de las Cruces, San Luis Potosí, México // Rev. Mex. Cienc. Geol, 2012. V. 29. P. 299—307.
- Rowan J., Martini P., Likius A., Merceron G., Boisserie J.-R. New Pliocene remains of Camelus grattardi (Mammalia, Camelidae) from the Shungura Formation, Lower Omo Valley, Ethiopia, and the evolution of African camels // Historical Biology, 2019. V. 31, № 9. P. 1123—1134. DOI: 10.1080/08912963.2017.1423485.
- Ruez D. Diet of Pleistocene Paramylodon harlani (Xenarthra: Mylodontidae): review of methods and preliminary use of carbon isotopes // Tex. J. Sci., 2005. V. 57. P. 329.
- Silaev V. I., Ponomarev D. V., Kiseleva D. V., Smoleva I. V., Simakova Yu. S., Martirosyan O. V., Vasil'ev E.A., Khazov A.F., Tropnikov E.M. Mineralogical-Geochemical Characteristics of the Bone Detritus of Pleistocene Mammals as a Surce of Paleontological Information // Paleontological Journal, 2017. V. 51. № 13. Р. 1395—1421.
- Titov V. Earliest Paracamelus of the Old World / The 33rd intern. geol. congr. (Oslo, 6—14 Aug. 2008). Oslo. 2008a. P. 1173.
- Titov V. V. Camels´ remains from the archaeological sites of the early Iron Age of the Lower Don Region // Die kultur- und naturraumliche Entwicklung des Dondeltas: abstr. Intern. tagung (8—10 Dezember 2011, Berlin). Berlin, Deutsches Archaologisches institute, 2011. P. 11— 12.
- Titov V. V. Habitat conditions for Camelus knoblochi and factors in its extinction // Quaternary international, 2008b. Vol. 179. P. 120—125. DOI:10.1016/j.quaint.2007.10.022.
- Titov V. V., Logvinenko V. V. Early Paracamelus (Mammalia, Tylopoda) in the Eastern Europe // Acta zoologica cracovensia, 2006. V. 49. № 1—2. P. 163—178.
- Trayler R. B., Dundas R. G., Fox-Dobbs K., Van DeWater P. K. Inland California during the Pleistocene—megafaunal stable isotope records reveal new paleoecological and paleoenvironmental insights // Palaeogeogr. Palaeoclimatol. Palaeoecol., 2015. V. 437. P. 132—140.
- Vetter L. Paleoecology of Pleistocene Megafauna in Southern Nevada, USA: Isotopic Evidence for Browsing on Halophytic Plants. University of Nevada, Las Vegas, 2007. 91 p.
- Votyakov S. L., Kiseleva D. V., Shcapova Yu. V., Smirnov N. G., Sadykova N. O. Thermal properties of Fossilized mammal bone remnants of the Urals // Journal of Thermal Analysis and Calorimentry, 2010. V. 101. P. 63—73.
- Yann L. T., DeSantis L. R. G., Koch P. L., Lundelius E. L. Dietary ecology of Pleistocene camelids: influences of climate, environment, and sympatric taxa // Palaeogeography, Palaeoclimatology, Palaeoecology, 2016. V. 461. P. 389—400 http://dx.doi.org/10.1016/j.palaeo.2016.08.036.
- Yoyce D. J. Chronology and new research on the Schaefer mammoth (?Mammuthus primigenius) site, Kenosha County, Wisconsin, USA // Quaternary International, 2006. № 142. Р. 44—57.
