Использование мужской стерильности типа OGURA в селекции редиса на гетерозис

Автор: Федорова М.И., Заячковская Т.В.
Журнал: Овощи России @vegetables
Рубрика: Современные направления селекции овощных культур
Статья в выпуске: 4 (13), 2011 года.
Бесплатный доступ
Для определения ЦМС-Ogura и отбора растений с этим типом стерильности в практической селекции редиса выявлены маркерные морфологические особенности цветка и цитологические особенности микрога-метогенеза и микроспорогенеза стерильных форм. В результате анализа семенных растений гибридных и инбредных потомств редиса на основе андростерильной формы дайкона по признаку ms получены и выделены потомства со 100%-ной стерильностью, которые являются исходными линиями для создания гетерозисных гибридов редиса.
Редис, цветок, цмс- ogura, гибриды f 1, инбредные линии
Короткий адрес: https://sciup.org/14024908
IDR: 14024908 | УДК: 635.152:631.527.52/527.56
The use of male sterility of OGURA type in radish breeding for heterosis
The morphological marker characteristics of the flower and cytological features of microgametogenesis and microsporo-genesis in sterile plants have been studied to identify and select plants bearing Ogura male sterility type. As result of analysis of hybrid plants and inbreeding progenies based on male sterile form of Japanese radish the three groups of progenies were observed with 100 % of sterile plants, all of them are to be the initial breeding lines for production of radish hybrids F 1.
Текст научной статьи Использование мужской стерильности типа OGURA в селекции редиса на гетерозис
В каталогах ведущих зарубежных семеноводческих фирм Японии, Франции, Нидерландов и в других странах (Beijo Zaden, Rick Zwaan, Roijl Sluis) предлагаются F1 гибриды редиса, производство которых занимает 90100% площадей. В нашей стране зарубежными фирмами предлагаются семена F1 гибридов редиса, созданных на основе мужской стерильности (Донар, Тарзан, Ровер и др.). По данным на 2011 год в Государственный реестр селекционных достижений включено 24 гибрида F1 редиса, в основном иностранной селекции (Нидерланды, Франция и др.).
Использование самонесовместимости в селекции культурных растений семейства капустных было начато в США (Pearson O.H., 1932), затем в Японии, Болгарии, Великобритании, Нидерландах и в нашей стране. В 70-е годы во ВНИИССОК В.Я. Кравчуком выделены самонесовмести-мые растения среди сортов редиса Розово-красный с белым кончиком и Тепличный Грибовский путем анализа за- вязываемости семян от автогамного опыления цветков. Выделены и отобраны гомозиготные растения по аллелям самонесовместимости, в результате оценки их на комбинационную способность отобрано 6 самонесовместимых линий, гибриды с которыми превосходили исходные сорта по массе корнеплода на 22-65% (Кононков П.Ф., Кравчук В.Я., Мохов А.И., Рабунец Н.А., 1977).
Спорофитная система самонесовместимости для достижения высокой гомозиготности и получения изогенных линий требует ручного опыления бутонов. Это трудоемкий процесс, что и приводит к сокращению числа родительских компонентов (Монахос, Барашева, 1999).
Наиболее полно преимущества генетического подхода проявляются при использовании цитоплазматической мужской стерильности. Среди растительного разнообразия видов Brassica L. форма с ядерно-цитоплазматической мужской стерильностью впервые обнаружена Ogura (1968) у неидентифицированного сорта японского подвида редьки – дайкона. В семидесятых годах Боннет (Bonnet A., 1975) интродуцировал форму мужскостерильного дайкона во Францию и передал мужскую стерильность редису. Это позволило ему создать линии редиса – закрепители стерильности и уже в 1977 году в официальном каталоге Франции был зарегистрирован первый F1 гибрид редиса, созданный на основе этого типа стерильности.
Во ВНИИССОК ведутся исследования по созданию гибридов с 1990 года. Среди обширной коллекции сортоо-бразцов дайкона, интродуцированнной в 1988 году в Россию Буниным М.С., выделена форма мужскистерильного дайкона. Гибридологический анализ показал, что андро-стерильность у него контролируется взаимодействием между рецессивным ядерным геном в гомозиготном состоянии msms и стерильной цитоплазмой (S), то есть соответствует Ogu-ЦМС (Бунин М.С., 1992). Большой научный и практический интерес для гетерозисной селекции различных видов растений семейства Brassicaceae L. имеет использование этой формы дайкона в качестве донора Ogu-ЦМС.
В лаборатории селекции и семеноводства столовых корнеплодов ВНИИССОК проводятся исследования по передаче ЦМС-ogu от дайкона редису. Путем скрещивания фертильных линий с линией ЦМС-ogu установлено, что во многих гибридных поколениях наблюдается 100% фертильность. Предположительно, большинство сортов и селекционных форм редиса, участвовавших в скрещиваниях, имеют доминантный по фертильности генотип и использованные опылители подходят для выделения из них закрепителя стерильности. Выявлены особенности наследования и характер проявления основных морфобиологических признаков в гибридном потомстве F1 при скрещивании андростерильной формы дайкона с сорто-популяциями редиса и дайкона (Бунин М.С., Кириллова А.Б., 1997; Даньков А.М., 2001; Заячковская Т.В., 2005; Заячковская Т.В., Федорова М.И. и др., 2005).
На Селекционной станции им. Н.Н. Тимофеева созданы пары МС-линия-закрепитель ядерно-цитоплазматической стерильности, впоследствии созданы и внесены в Госре-естр первые отечественные гибриды дайкона Император и Фламинго, созданные на основе ogu-ЦМС. Также созданы стерильные линии лобы и линии закрепителя стерильности на основе гибридизации выделенного из коллекции селекционной станции стерильного образца дайкона с отборами из лобы. Во ВНИИ овощеводства также начата работа по оценке и созданию исходного материала для гетерозисной селекции редиса (Зубик И.Н., 2005; Миронов А.А., 2010; Янаева Д.А., 2011).
Материалы и методы
Исследования проводили в условиях защищенного грунта. Материал для исследований – образцы отечественной и зарубежной селекции: гибридные популяции и сортопопуляции редиса Королева Марго, Фея, Дуро, Красный великан, Французский завтрак, Родос, Вариант, Моховский; перспективные сортообразцы редиса – 42/03 (сортотип розово красный с белым кончиком), 43/03 (сортотип фиолетовый), 45/03 (сортотип красный), F1 Тарзан; сортопопуляции дайкона.
Маточные корнеплоды выращивали по общепринятой методике. Для скрещиваний отбирали хорошо развитые семенные растения, которые изолировали в фазу бутонизации индивидуальными изоляторами. Опыление проводили вручную. Гибридные потомства от скрещивания ms x mf анализировали по признаку стерильности в период массового цветения, а также по хозяйственноценным параметрам. Стерильность определяли числом стерильных растений от их общего количества в процентах. Методы работы – инбридинг, беккроссирова-ние, сибсовые скрещивания и отбор. Длину частей цветка стерильных и фертильных растений на разных стадиях развития бутонов измеряли с использованием линейки на бинокулярной лупе «OPTON M 35 W». Цитологический анализ проводился на временных ацетокар-
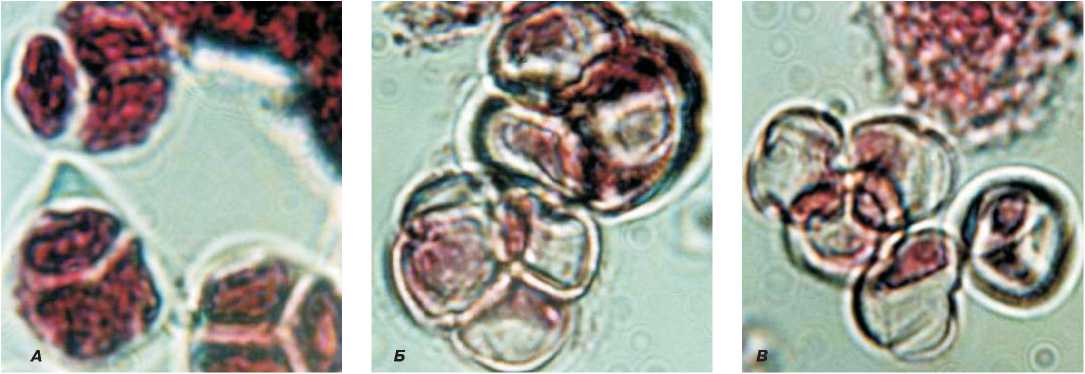
Рис. 1. Развитие микроспор цветка редиса с ЦМС- Ogura.
А – тетрады, Б – не распавшиеся тетрады, В – стерильная пыльца.
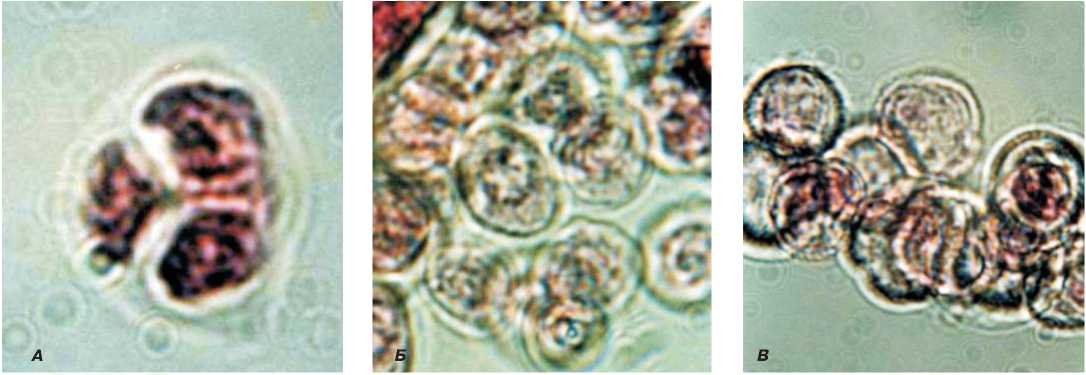
Рис. 2. Развитие микроспор цветка дайкона с ЦМС-Ogura, (х90). А – тетрада, Б – распад тетрад, В – стерильная пыльца.
миновых препаратах по методике Паушевой, а для окрашивания пыльников использовали дифференциальный краситель (Паушева, 1988; Данвелл, 1989).
Создание ms-линий редиса проводили с использованием генисточника андростерильности формой дайкона с ЦМС-Ogura – MS Gensuke. Получены беккроссиро-ванные и инбредные потомства на основе гибридов F1 мс х Моховский, мс х Французский завтрак, мс х Вариант, мс х 43/03, мс х Красный Великан – 203/07-209/07, 216/07-223/07, 213/07-215/07, 314/07, 200/07-212/07 и 313/07.
Всего изучено 98 беккроссированных потомств (ВС1 – ВС3), полученных на основе исходных частично стерильных 14-ти популяций F1, 334 инбредных потомств (J1 – J5), 69 гибридных комбинаций.
Результаты и обсуждение
Несомненную актуальность представляет собой иссле- дование Ogu-ЦМС с точки зрения развития пыльцы цветков у фертильных растений и ЦМС-форм, что имеет научный приоритет. В связи с этим для более полного представления о данном типе стерильности совместно с лабораторией биотехнологии ВНИИССОК (Шмыковой Н.А.) проведено сравнительное изучение морфологических особенностей цветка и развития пыльника стерильных и фертильных цветков растений.
Путем цитологического анализа установлены особенности нарушения развития пыльника стерильного дайкона и редиса.
Они становятся заметными, как правило, на стадии тетрад или ранних одноядерных микроспор (рис.1, 2). В пыльниках бутонов длиной 2,5 мм в одноядерных микроспорах не происходит утолщение оболочки. Микроспоры слипаются, образуя конгломераты, которые не контактируют со стенками пыльника в отличие от микроспор фертильных растений, которые не слипаются, а равномерно распреде- ляются по периметру стенки пыльцевого гнезда (рис. 3, 4). В итоге в бутонах дайкона размером 3 мм пыльца уже стерильная. Следствием нарушения процессов микроспорогенеза и развития соматических тканей пыльника у андростерильной формы дайкона является уменьшение размера пыльника, которое начинается после распада тетрад в бутонах длиной 2 мм.
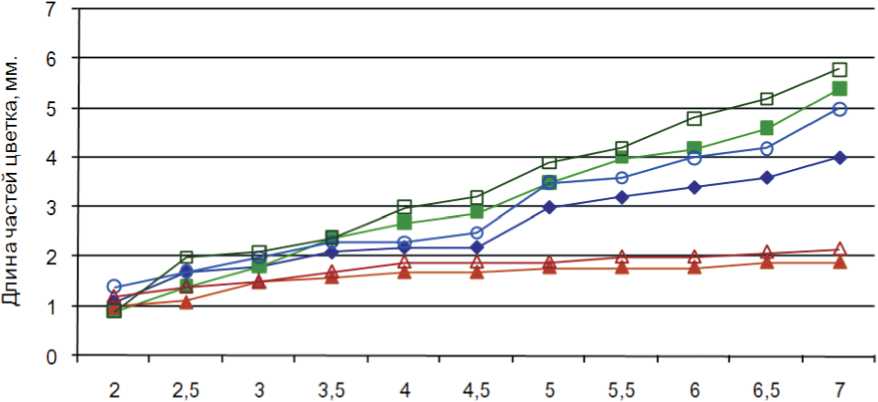
На графике представлена динамика развития основных частей цветка дай-
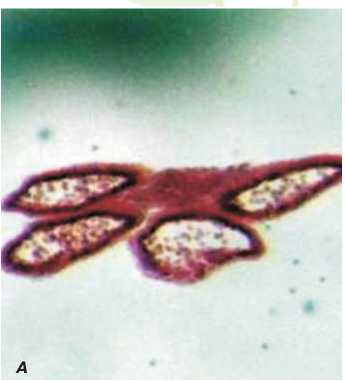
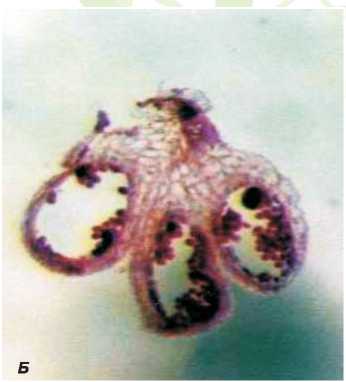
кона в процессе онтогенеза. Анализ данных показал, что уже в бутонах длиной 2 мм пыльник фертильных растений дайкона длиннее пыльника стерильных растений на 0,2 мм (рис. 5). Превышение в длине пестика над тычинкой наблюдается при всех исследуемых размерах бутона как фертильных, так и стерильных цветков.
В этот период растения с ЦМС можно определить визуально: внутри бутона длиной 5мм хорошо видны темножелтые или коричневые, слипшиеся пыльники дайкона и редиса с ЦМС-Ogura (рис 6).
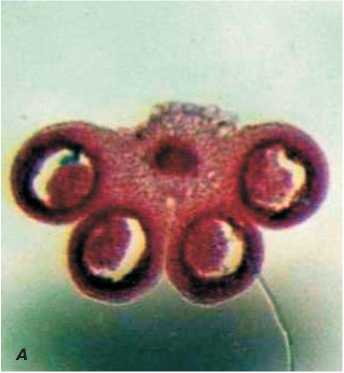
Рис. 4. Поперечный срез пыльников стерильного цветка дайкона (увеличение 6,4).
А - на стадии развития одноядерных микроспор, Б - на стадии развития зрелой пыльцы.
Рис.3. Поперечный срез пыльников фертильного цветка дайкона (увеличение 6,4).
А - на стадии развития одноядерных микроспор, Б - на стадии развития зрелой пыльцы.
Длинабутона, мм
Рис. 5. Изменение длины частей цветка стерильных и фертильных растений дайкона в онтогенезе.
-
- ♦— тычинка МС
-
- ■— пестик МС
-
- *— пыльник МС
-
■ е— тычинка МФ
в— пестик МФ
-
- 6— пыльник МФ


Рис. 6. Пыльники и пестик фертильного и стерильного цветка растений: А - редиса, Б - дайкона, длина бутона 5мм (*90).
Таким образом, на самых ранних стадиях развития цветка различия в развитии пестика и тычинки как стерильных, так и фертильных растений незначительны. Только в самых крупных бутонах длиной 6,5 мм непосредственно перед раскрытием цветка фиксируются морфологические нарушения в развитии тычинки стерильных цветков, проявляющиеся в увеличивающейся разнице ее длины по сравнению с длиной пестика.
Изучение размеров частей раскрытых фертильных и стерильных цветков показало, что внешнее и внутреннее строение обоих типов цветка одинаковое. Бледно-розовые лепестки цветка изучаемой формы дайкона с ЦМС по окраске практически не отличались от окраски лепестков цветка фертильного дайкона (рис. 7).
Наблюдения в период массового цветения растений дайкона и редиса также показали, что, цветки растений мужскистерильной формы дайкона отличаются от фертильных цветков величиной основных частей. Цветки муж-скистерильной формы дайкона меньше, чем цветки фертильных растений. Размер цветка фертильных растений дайкона и редиса варьировал в пределах 20,1-21,6 мм, 17,8-19,9 мм, соответственно. Как и размеры цветка, так и его части – тычинка, пестик, пыльник и цветоножка – у стерильных растений дайкона меньше, чем соответствующие части цветков фертильных растений. Визуально, наиболее заметен выступающий над тычинками пестик, что является маркерным признаком полностью стерильных растений дайкона (рис. 7).
Отставание пыльников в развитии в процессе онтогенеза цветка приводит к тому, что в раскрытых цветках темно-жел-
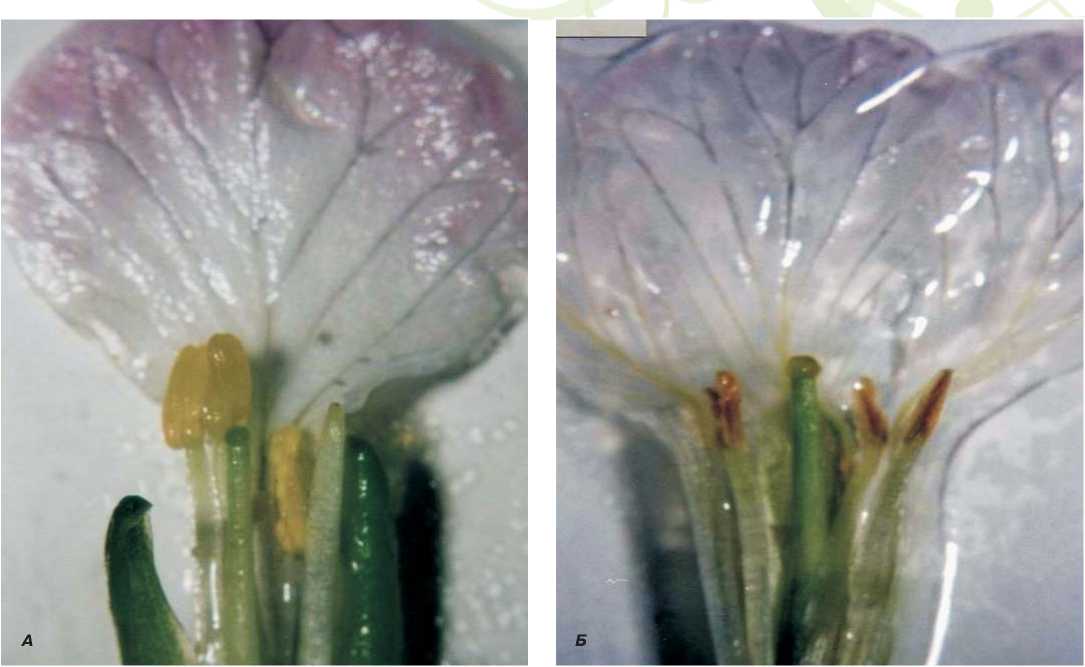
Рис. 7. Соотношение частей цветка фертильных и стерильных растений дайкона. А - цветок фертильного растения, Б - цветок стерильного растения
тые или коричневые, сморщенные, без высыпающейся пыльцы пыльники стерильного дайкона короче на 0,56мм, чем у хорошо сформированных пыльников фертильных растений дайкона, которые имеют ярко-желтую окраску и легко растрескиваются. Полученные данные являются основой для определения стерильности типа ЦМС-Ogura на морфологическом и цитологическом уровнях в селекционной работе с использованием этого типа стерильности (Бунин М.С., Шмы-кова Н.А., Заячковская Т.В., 2004).
Гибридное потомство от скрещивания стерильной формы и фертильных растений анализировали по признаку ms и группе хозяйственно ценных параметров с целью отбора перспективных гибридных комбинаций для селекции на гетерозис. Гибридные комбинации получены с участием в качестве отцовских компонентов растений сортопопуляций редиса и дайкона. В результате анализа гибридного потомства по признаку стерильности в 9 гибридных популяциях из 14 обнаружены растения с признаками мужской стерильности.
Количество растений с признаками стерильности в полученных гибридных комбинациях 10 – 71%; доля цветков с признаками стерильности – 65-100%.
Таким образом, в результате анализа признака стерильности гибридного потомства, полученного на основе сте- рильной формы дайкона, были выделены перспективные сортопопуляции редиса Дуро, Французский завтрак и №42/03, Моховский, Вариант. Они использованы как перспективные источники для будущих линий – закрепителей признака стерильности.
Гибриды F1 различались между собой по ряду морфологических признаков листовой розетки и корнеплода (рис. 8). Форма листовой розетки варьировала в наибольшей степени среди всех качественных признаков гибридов F1 (Cv=39,66%). Также как и у материнской формы дайкона, гибриды характеризовались крупной листовой розеткой (30,0-40,2 см), рассеченными узколировидными листьями, наличием опушения на черешках листьев (рис. 8). Гибриды, полученные от скрещивания с разными формами дайкона, имели больше листьев в розетке (9-11 шт.), чем гибриды, полученные от скрещивания дайкона и редиса (7-9 шт.). Рассеченность их достигала 12-13 долей, тогда как у последних – 6-9 долей. Варьирование по признакам длины, ширины и формы листовой пластинки было незначительным (Cv=13,58%, 13,29% и 10,29%, соответственно). Длина листовой пластинки колебалась от 23 до 35 см., что значительно большее листовой пластинки редиса (на 20-30%).


Рис. 8. Гибриды F1 с редисом : 2 - Фея, 7 - Королева Марго, 9 - №43/03, 11 - Моховский, полученные на основе стерильной формы дайкона.
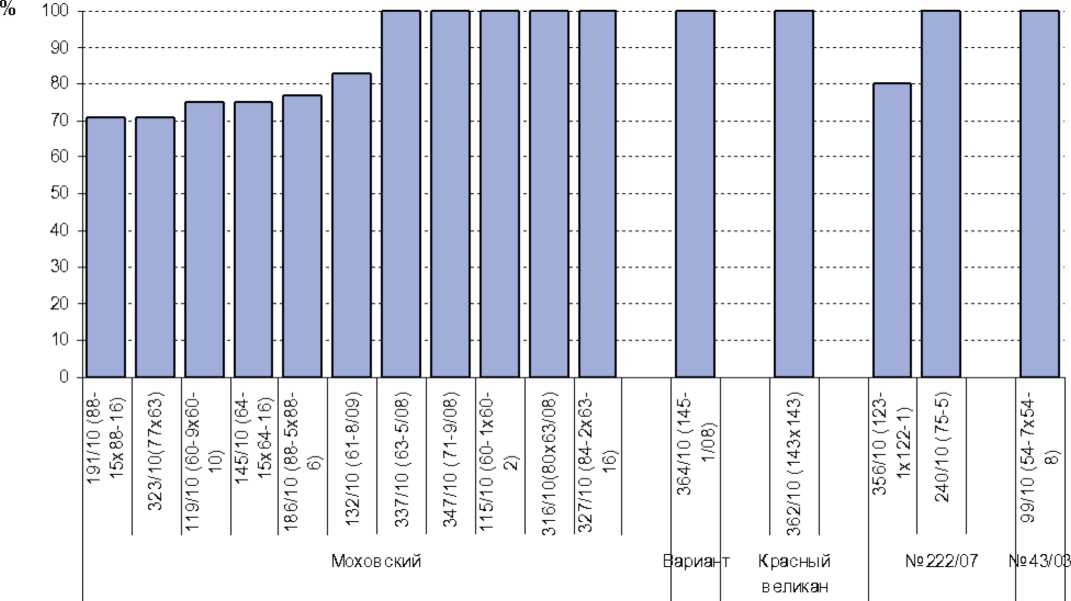
гибридное потомство, исходная популяция
Рис. 9. Проявление стерильности в инбредных потомствах и гибридных комбинациях
При скрещивании длиннокорнеплодного дайкона (масса 122 г) с корнеплодами редиса (округлая форма, масса 15-20 г) у гибридов F1 преобладали корнеплоды промежуточной формы, которые значительно превосходили по массе отцовские родительские компоненты. Отмечена значительная изменчивость формы и индекса формы корнеплодов гибридов F1 (Cv=51,32% и Cv=55,15%, соответственно). В связи с этим гибриды были сгруппированы по форме: I – округло-овальные корнеплоды с массой 130-163 г, II – овальные с массой 100-225 г, III – цилиндрические с массой 57-117 г.
Гибриды, где в качестве отцовских компонентов использовались сортопопуляции редиса Вариант, Французский Завтрак и Фея характеризовались наибольшей массой корнеплода 152-225 г. Варьирование признака длины и диаметра корнеплода гибридов было наименьшим среди количественных признаков и составляло Cv=21,56% и Cv=26,06%, соответственно.
Структура наружной окраски корнеплодов гибридов F1 в большинстве случаев неоднородная как в пределах корнеплода (Cv=25,35%), так и в пределах всего потомства (Cv=44,38%). Преобладающая на- ружная окраска изменялась от белой (у гибридов с дайконами) до фиолетовой (у гибридов с редисами); степень интенсивности окраски – от головки к основанию корнеплода.
Масса растений гибридов также варьировала (Cv=29,49%) и составляла 103-305 г, при массе отцовских растений редиса – 18-38 г. При этом доля листовой розетки большинства гибридов уменьшилась в общей массе растения по сравнению с обеими родительскими формами.
Гибридные комбинации с округло-овальной и овальной формой корнеплода МС х Фея, МС х Мохов-ский, МС х Французский Завтрак и МС х Вариант характеризуются преобладающей долей корнеплода в массе растения и высокой продуктивностью по сравнению с родительскими компонентами и являются наиболее перспективными для создания закрепителя стерильности, а также исходных линий с высокими хозяйственно-ценными показателями, обеспечивающими проявление гетерозисного эффекта в гибридных комбинациях с андростерильной формой дайкона.
Выделенные исходные гибридные популяции F1 характеризовались степенью стерильности в пределах 25-
1. Перспективный исходный линейный материал для гетерозисной селекции редиса (ms-100%)
|
Исходная популяция |
Гибридное потомство |
|
Моховский |
457/11 (127-2х127-3) |
|
474-А/11(162-2мф) |
|
|
477/11 (163-2мф) |
|
|
481/11 (163-6мф) |
|
|
508/11 (320-1мсх320-2мф) |
|
|
512/11 (332-1мсх331-1мф) |
|
|
Красный великан |
569/11 (362-3х363-4) |
|
№222/07 |
560/11 (356/11мсх360-1мф) |
|
631/11 (238-1мс х 238-2мф) |
|
|
Вариант |
619/11 364/10 (145-1/08) |
|
619-А /11 364/10 (364-2мсх366-2мф) |
|
|
620/11 (366-2мф) |
|
|
623/11 (366-3мсх366-4мф) |
|
|
№43/03 |
434/11 (92-1мс х 92-2мф) |
50%. В первом поколении беккросса ВС1 выделено шесть потомств со степенью стерильности 33-100%; в 26 изученных потомствах ВС2 стерильность варьировала от 10 до 100%, в трех потомствах все растения оказались стерильными. Однако степень наследования стерильности в результате третьего беккроссирования в этих трех потомствах составила 30-63%. В результате дальнейшего беккроссирования получено 52 потомства ВС3 со степенью стерильности 19-100%, два – со степенью стерильности 3-6%, а также одно полностью фертильное потомство. Анализ скрещиваний, проведенных на основе сортопопуляций редиса Моховский, Вариант, 43/03, Красный великан, позволил выделить пять ин-бредных потомств со степенью стерильности 83-100 % и 9 гибридных комбинаций ms x mf – со стерильностью 71-100% (рис. 9). Выделено 14 потомств, гибридных комбинаций ms x mf и линий со 100%-ной стерильностью. В трех из них №№ 619, 619А, 569 стерильность сохраняется в последующих потомствах и закреплена №№ 363-4, 366-2 (табл. 1). Таким образом, созданы стерильные линии и закрепители как перспективные для создания гетерозисных гибридов редиса F1.
Заключение
Сравнительное изучение развития пыльников цветков стерильных и фертильных растений, особенно процесса развития микроспор in vivo, необходимо для раскрытия причин нарушения процессов микроспорогенеза в пыльниках растений с ЦМС – Ogura. В пыльниках андростерильной формы дайкона нарушения в развитии микроспор проявляются на стадии тетрад или ранних одноядерных микроспор. Следствием нарушения является уменьшение размера пыльника цветков стерильных растений, которые начинаются после распада тетрад в бутонах длиной 2 мм. В самых крупных бутонах длиной 6,5 мм непосредственно перед раскрытием цветка фиксируется отставание в развитии тычинки от пестика. Полностью раскрытые цветки стерильных растений имеют меньший размер основных частей цветков, деформированные пыльники и отлича- ются от фертильных наличием выступающего над тычинками пестика, что является маркерным признаком для данного типа стерильности.
Изучение изменчивости основных морфобиологических признаков гибридов F1 на основе ЦМС-Ogura позволило провести оценку используемых в качестве опылителей сортопопуляций редиса и дайкона различных сортотипов, как потенциальных родоначальников линий с высокими показателями, обеспечивающи- ми проявление гетерозисного эффекта по большинству морфобиологических признаков. Гибриды F1 характеризовались крупной листовой розеткой, рассеченными узколировидными листьями, наличием опушения на черешках листьев, как у материнской формы дайкона. Наследование массы корнеплода происходит по промежуточному типу. ЦМС-Ogu передавалась большинству гибридных потомств со степенью стерильности 3-70%.
Список литературы Использование мужской стерильности типа OGURA в селекции редиса на гетерозис
- Бунин М.С. Создание и использование андростерильной линии дайкона в селекции гибридов F1 редьки и редиса. Материалы VI съезда Всероссийского общества генетиков и селекционеров им. Н.И Вавилова. -М., 1992. Ч. 11. -С. 23.
- Бунин М.С., Кириллова А.Б. Использование мужской стерильности в селекции гетерозисных гибридов корнеплодных овощных культур вида Raphanus sativus L. Гетерозис сельскохозяйственных растений. Международный симпозиум, Т. 5, XII, 1997. -М. -С. 9496.
- Бунин М.С., Шмыкова Н.А., Иванушкина Т.В. Методические рекомендации по определению жизнеспособности пыльцы культур вида Raphanus sativus. -М. -2003. -34с.
- Бунин М.С., Шмыкова Н.А., Иванушкина Т.В. Цитологическое изучение особенностей развития пыльника растений вида Raphanus sativus с ЦМС -Ogura.//Доклады ТСХА. -М.: Изд-во МСХА. -2004. -В. 276. -С.461-465.
- Данвелл Д.М. Культура гаплоидных клеток//Биотехнология растений: культура клеток/Пер. с анг. Под ред. Р. Г. Бутенко. -М.: Агропромиздат, 1989. -С. 3351.
- Даньков А.М. Исходный материал для селекции редиса по основным хозяйственно ценным признакам//Канд.дисс. -М. -2001.
- Заячковская Т.В. Оценка исходного материала культур вида Raphanus sativus с использованием методов репродуктивной биологии. Канд. Дисс. -М. -2005.
- Заячковская Т. В., Федорова М. И., Бунин М.С., Степанов В. А., Заячковский В. А. Исходный материал редиса для селекции на гетерозис.//Современное состояние и перспективы развития селекции и семеноводства овощных культур. Международный симпозиум (9-12 августа 2005), Т. 2. -М., 2005. -С. 137-145.
- Зубик И.Н. Наследование основных хозяйственных признаков и комбинационная способность инбредных линий дайкона. Автореф. дис. канд. с.-х. наук. М. -2005. -21с.
- Кононков П.Ф., Кравчук В.Я., Мохов А.И., Рабунец Н.А. Самонесовместимость и использование ее для получения гетерозисных гибридов редиса. Труды по селекции овощных культур. -М. -1977. -с.17-21.
- Миронов А. А. Создание мужски стерильных линий лобы (Raphanus sativus L. convar. Lobo Sazon. et Stankev), оценка комбинационной способности устойчивых к киле линий. Автореф. дис. канд. с.-х. наук. М. -2010. -2 с
- Монахос Г. Ф., Барашева Г.М. Наследование массы корнеплода инбредными линиями редиса (Raphanus sativus L. var. sativus)//Известия тимирязевской сельскохозяйственной академии. -1999. Вып. 1. -С. 92100.
- Паушева З.П. Практикум по цитологии растений. -М.: Агропромиздат, 1988. -271 с.
- Янаева Д. А. Создание исходного материала для гетерозисной селекции редиса европейского (Raphanus sativus L. var. Sativus) для защищенного грунта. Автореф. дис. канд. с.-х. наук. М. -2011. -2 с.
- Bonnet A. Introduction and utilization of a cytoplasmatic male sterility in early European varieties of radish-Raphanus sativus L.//Ann. Amelior. Plantes. -1975. -V. 25. -№4. -P. 381-397.
- Ogura H. Studies on the new male sterility in Japanese radish, with special reference to the utilization of this sterility towards the practical raising of hybrid seeds. Mem. Fac. Agr. Kagoshima Univ. -1968. -№ 6. -P. 39-78.
- Pearson O.H. Breeding plants of the cabbage group. -California Exp. Sta. Bull.1932. P.532.
