Использование онтогенетического подхода при регистрации кривой Каутского в ходе стрессового воздействия на растения листовой капусты

Автор: Шихов В.Н., Нестеренко Т.В., Тихомиров А.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 7, 2016 года.
Бесплатный доступ
В статье рассмотрены изменения формы индукционной кривой медленной флуоресценции хлорофилла листьев листовой капусты сорта Бансэй Маруба в норме и при обратимом стрес-се в условиях светокультуры. Использование метода индукции флуоресценции хлорофилла (ИФХ) было сопряжено с применением предло-женного авторами ранее онтогенетического подхода для анализа полученных данных. Приме-нение этого подхода до сих пор ограничивалось анализом численных параметров. В данной ра-боте онтогенетический подход использован для сравнения форм индукционных кривых флуорес-ценции, которые также имеют свою зависи-мость от возраста листа. В качестве стрессо-вого воздействия для опытной части растений был создан водный дефицит сроком на 3 суток. Было произведено сравнение параметров ИФХ и форм индукционной кривой флуоресценции ли-стьев капусты в контрольных условиях, в усло-виях стресса и при последующей репарации. Анализ параметров ИФХ показал, что стрессо-вая реакция у растений листовой капусты носи-ла полностью обратимый характер. Наблюдае-мые в данном случае в стрессовый и репараци-онный период изменения формы кривой ИФХ с точки зрения формально примененного онтоге-нетического подхода можно ошибочно охарак-теризовать как «омоложение». На основе полу-ченных данных сделан вывод, что сложившееся представление о сходстве процессов, вызван-ных стрессовым воздействием на раститель-ный организм, и процессов старения нуждается в коррекции. При этом онтогенетический под-ход, как элемент комплексного интегрального анализа состояния растений, продемонстриро-вал свои возможности выявлять произошедшие в растительном организме изменения.
Индукция флуоресценции хлорофилла, стресс, листовая капуста
Короткий адрес: https://sciup.org/14084731
IDR: 14084731 | УДК: 581.1:
The use of ontogenetic approach for the Kautsky curve recording during the stress impact on colewort plants
In the study the changes of the chlorophyll induc-tion curve’s shape for colewort (Bansei Maruba) leaves in normal and under the reversible stress in the conditions of light culture were investigated. The use of the chlorophyll fluorescence induction (CFI) tech-niques was linked with applying the ontogenetic ap-proach that was earlier introduced by the authors for the data analysis. This approach was used only with numerical parameters till now. In this study the onto-genetic approach was used to compare the CFI curve shapes that also have age dependence for the leaves. The stress conditions were realized as a 3-day water deficit. Comparison of the parameters and the curve shapes of the colewort leaves’ CFI was ex-ecuted in the normal, stressed and reparation condi-tions. The analysis of CFI parameters showed that colewort plant response to stress conditions was completely reversible. It was shown that the consider-ing of the CFI curve’s shape changes for reversible stress conditions with the help of formally used on-togenetic approach can result into erratic assump-tion of “rejuvenation” effect. The data obtained has led to the conclusion that the well known thesis of similarity between the stress-induced changes and the aging processes in plants should be corrected. However, the ontogenetic approach as an element of complex integral analysis of plants’ condition has demonstrated the ability to detect the plant organ-ism’s changes.
Текст научной статьи Использование онтогенетического подхода при регистрации кривой Каутского в ходе стрессового воздействия на растения листовой капусты
Введение. Широко применяемый в мировой практике метод индукции флуоресценции хлорофилла (ИФХ) уже давно зарекомендовал себя как объективный и оперативный способ определения физиологического состояния фотосинтетического аппарата (ФСА) растений в нормальных и стрессовых условиях [1]. Одними из наиболее важных для понимания механизмов функционирования фотосинтетического аппарата растений на уровне листа в различных условиях являются характеристики медленной фазы (P-S-M-T-переход [1, 2]) ИФХ, так как они в большей степени связаны с биохимическими процессами фотосинтеза и интенсивностью СО2-газообмена в листьях [3–6]. При этом кинетика ИФХ регулируется рядом взаимосвязанных факторов, которые подробно рассматриваются в обзорах [1–4]. В том числе хорошо известна возрастная зависимость формы индукционной кривой медленной флуоресценции интактных листьев [7]. Одним из аспектов использования данной зависимости являются стрессовые для растений условия. Считается, что разные стрессоры вызывают, как правило, ускорение старения ФСА растений [8, 9]. При этом обычно происходят такие же изменения формы индукционной кривой флуоресценции, как и при старении листа [7]. Встречается и противоположный тип ответной реакции. Например, патофи- зиологические изменения при вирусной инфекции картофеля проявлялись в удлинении срока вегетации по сравнению со здоровыми растениями и в более позднем появлении «сложной» кинетики флуоресценции у вирусных растений, что свидетельствует об их замедленном развитии [9].
Сходство процессов старения растительного организма и вызванных стрессовыми факторами изменений, оцениваемых по показателям ИФХ, послужило основой предложенного ранее авторами данной работы онтогенетического подхода [10] в оценке состояния высших растений. Онтогенетический подход показал себя в качестве полезного инструмента в экспериментальных работах [11], позволяя получать иногда оценку состояния растений, дополняющую традиционные подходы. При этом обычно не возникал вопрос применения онтогенетического подхода при использовании для характеристики состояния высших растений не только численных параметров ИФХ, но и качественного вида индукционной кривой. Тем не менее, подобный вопрос, несмотря на кажущуюся архаичной саму его постановку, может представлять определенный практический интерес.
Цель исследования : Изучение особенностей онтогенетического подхода при использовании качественного вида кривой ИФХ для оценки состояния растений в условиях обратимых стрессовых воздействий.
Объекты и методика исследования . Растения листовой капусты сорта Бансэй Маруба были выращены в Институте биофизики СО РАН в условиях полной светокультуры на вегетационной установке закрытого типа. Освещение производилось комбинацией ламп ДРФ-1000 и ДРЛ-1000. Освещенность в камере составляла порядка 100 Вт/м2 ФАР. Температура воздуха на протяжении всего эксперимента автоматически поддерживалась на уровне 24±1 ºС, влажность воздуха составляла около 70 %. Растения выращивали в двух раздельных вегетационных сосудах на биологическом субстрате, подготовленном по модифицированной методике, основа которой приведена в работе [12]. Полив осуществлялся автоматически насосами из специальных емкостей внутри камеры один раз в сутки. Концентрация СО 2 была атмосферная, камеру не герметизировали.
В качестве объективного контроля физиологического состояния растений в работе использовали метод импульсно-модулированной флуоресценции хлорофилла [4–5]. Измерения проводили на интактных листьях с помощью флуориметра PAM-2100 производства фирмы Walz (Германия). Перед измерениями флуоресценции растения предварительно адаптировали в темноте в течение 30 мин [13]. Интенсивность действующего белого света от внутреннего галогенного источника составляла порядка 200 Вт/м2 ФАР. Измерения флуоресценции проводили в 4 повторностях для каждого исследуемого варианта на зрелых листьях, находящихся в верхней части растительного ценоза, где условия освещенности были достаточно однородны.
По достижению растениями возраста 21 суток один из вегетационных сосудов с растениями был отключен от системы полива с целью создать стрессовые условия водного дефицита. Стрессовые условия длились 3 суток до полной потери тургора листьев и контролировались при помощи системы видеомониторинга, описанной ранее [13], на 4-е сутки полив был возобновлен. Измерения флуоресценции проводили ежедневно, при возобновлении ирригации измерения были проведены через 2 часа после ирригации. Еще через сутки после возобновления ирригации были выполнены измерения флуоресценции для оценки репарационных процессов в растениях.
Результаты исследования и их обсуждение. Данные по импульсно-модулированной флуоресценции хлорофилла, полученные в результате эксперимента, показывают изменение в сравнении с контролем значений ряда флуоресцентных параметров в ходе стрессового воздействия (табл. 1). Двухфакторный дисперсионный анализ данных таблицы демонстрирует достоверность отличий между контрольным и опытным вариантами для всех приведенных в таблице показателей.
Эти данные говорят о том, что отмеченные изменения в состоянии растений были полностью обратимыми и повреждения ФСА не произошло.
На рисунке 1 показан типичный вид кривой ИФХ в норме по состоянию до стрессового воздействия. В ходе стрессового воздействия и последующего репарационного периода у растений контрольного и опытного вариантов тип кривой ИФХ демонстрировал различные закономерности смены своего вида. Так, у растений, подвергнутых стрессовому воздействию, за трое суток такого воздействия заметно вырос как по амплитуде, так и по ширине второй пик на кривой медленной ИФХ, что в сочетании с его близостью к первому пику дало кривой ИФХ мощное «плечо» в этой области. При этом изменения качественного вида кривой ИФХ у опытных растений под действием стресса, представленные на рисунке 2, демонстрировали закономерность, похожую на изменение параметров импульсно-модулированной флуоресценции хлорофилла. Это подтверждается и данными таблицы, где приведены значения времени полуспада этих кривых ИФХ. Но при этом после прекращения действия стрессового фактора тип кривой ИФХ вернулся к виду, более похожему на исходный тип до воздействия, чем на тип кривой ИФХ у контрольных растений одного с ними возраста.
Рис. 1. Исходная типичная кривая МИФ листьев капусты сорта Бансэй Маруба в нормальных условиях среды, полученная за 3 суток до начала эксперимента
Изменения флуоресцентных параметров листьев капусты сорта Бансэй Маруба в ходе стрессового воздействия
|
Возраст растений, сут |
ETR, мкмоль/(м2 с) (F=6.865) |
Ф, отн. ед. (F=6.902) |
T 0.5 , с (F=5.711) |
|||
|
Контроль |
Опыт |
Контроль |
Опыт |
Контроль |
Опыт |
|
|
19 |
210A ±8 |
193a ±11 |
0.58C ±0.02 |
0.54c ±0.03 |
11.5E ±1.3 |
13.0e ±0.8 |
|
22 |
193AB ±6 |
188a ±24 |
0.54C ±0.02 |
0.52c ±0.07 |
14.0E ±1.2 |
21.3fg ±1.2 |
|
23 |
222A ±5 |
181ab ±16 |
0.61D ±0.01 |
0.50cd ±0.04 |
16.0E ±4.1 |
24.5f ±1.0 |
|
24 |
219A ±6 |
172abd ±13 |
0.60CD±0.02 |
0.48cd ±0.03 |
13.8E ±0.9 |
33.8fh ±3.8 |
|
25 (2 часа репарации) |
216A ±11 |
144bd ±12 |
0.59CD±0.03 |
0.40d ±0.03 |
10.3E ±1.3 |
14.8e ±0.9 |
|
26 |
210A ±4 |
234c ±8 |
0.57C ±0.01 |
0.64e ±0.02 |
12.0E ±1.1 |
16.3e ±1.7 |
|
29 |
160AB ±19 |
219ac ±9 |
0.45C ±0.05 |
0.62ce ±0.03 |
11.0E ±1.7 |
13.5e ±0.5 |
Примечание: начало стресса – в возрасте 22 суток, окончание – 25 суток, далее репарация. Для каждого из параметров указаны значения F-критерия по ANOVA при F кр =2.324, P < 0.05. Статистически достоверные отличия внутри вариантов обозначены разными буквами.
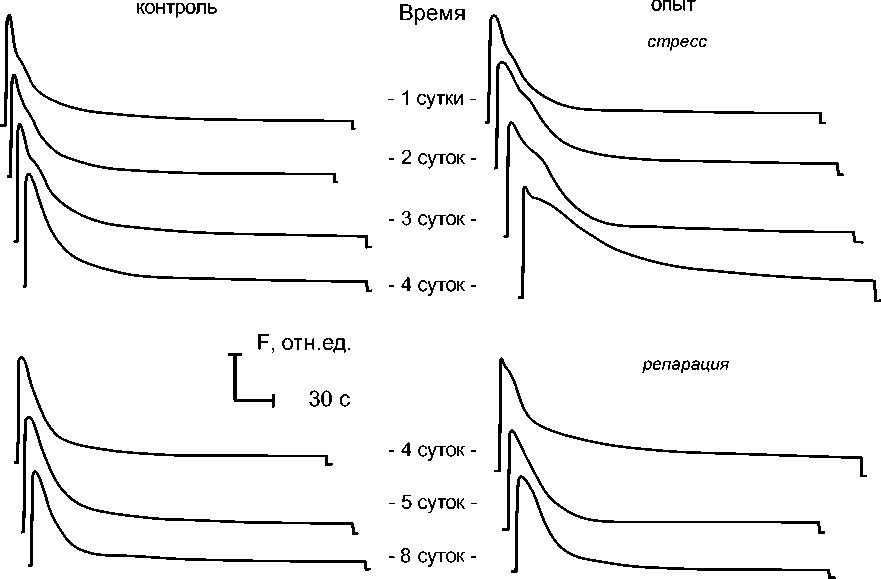
Рис. 2. Динамика типичных кривых МИФ хлорофилла листьев капусты сорта Бансэй Маруба в ходе нормального онтогенеза и при стрессе (засуха)
Анализируя полученные результаты с точки зрения онтогенетического подхода, можно увидеть одно существенное отличие между применением данного подхода к численным параметрам флуоресценции и к качественному виду кривой ИФХ. Численный параметр по своей природе формализован и обезличен. Поэтому применение онтогенетического подхода при анализе численных данных обладает достаточно высокой степенью универсальности по отношению к тем или иным ситуациям. Так, зная онтогенетическую зависимость изменения значе- ний какого-либо численного параметра, можно давать оценки состояния растительного организма и прогнозы его изменений на будущее [10]. И благодаря безличному характеру числового значения любого параметра не возникает проблем с трактовкой ситуации. Однако, когда для анализа используется качественный вид кривой (на примере ИФХ), исследователь имеет дело по сути с визуальным образом. В этом случае онтогенетическая зависимость качественного вида кривой ИФХ представляет собой серию таких визуальных образов, связанную закономерностью порядка смены одного образа другим, что хорошо было известно уже давно [1-2, 7]. Принципиальным отличием от численных параметров в данном случае является возможность существования уникальных образов кривой ИФХ, характерных для той или иной стадии онтогенеза листа или растения. Пока онтогенетический подход применяется для сравнения различных условий выращивания [11], указанный аспект не играет никакой существенной роли. Но как только этот подход начинает применяться в условиях действия какого-либо стресс-фактора, его формальное применение может привести к некорректной интерпретации полученных результатов.
В данной работе, применив онтогенетический подход к имеющимся зависимостям изменения типа кривой ИФХ листьев растений листовой капусты в норме и при обратимом стрессе, мы формально получаем «сенсационный» вывод об «омоложении» листьев капусты после прекращения действия стрессового фактора. Это следует из того факта, что на 8-е сутки эксперимента (4-е сутки после прекращения действия стресс-фактора) вид кривой ИФХ опытных растений был наиболее близок по форме к исходному типу кривой ИФХ у растений перед началом эксперимента, а не к какому-то типу кривой ИФХ листьев растений контрольного варианта, имевших место в ходе эксперимента. Для того, чтобы не получать мнимую «сенсацию», необходимо поставить под сомнение уже упоминавшийся ранее в данной работе подход, заключающийся в понимании стрессовой реакции растительного организма как процесса ускоренного (либо наоборот замедленного) онтогенеза.
Тем не менее, полученные нами результаты не означают, что онтогенетический подход в данном случае не работает. Скорее наоборот, можно сказать, что при практическом отсутствии повреждений ФСА какая-то остаточная реакция растений на пережитый стресс в виде водного дефицита все же имеет место. Это предположение хорошо согласуется с данными работы [14], где было показано, что пятидневная засуха для листа кукурузы не оказывает влияния на содержание и соотношение фотосинтетических пигментов, уровень перекисного окисления липидов и несущественно влияет на квантовый выход ФС2. Но при этом авторами были зафиксированы различия в уровне фотосинтетической ассимиляции СО2, связанные по всей видимости с изменением устьичной проводимости листа. А поскольку на форму кривой ИФХ влияют не только процессы непосредственно фотосинтетической трансформации поглощенной энергии, но и функционирование системы фиксации СО2 [15], то обнаруженный нами эффект изменения типа кривой ИФХ в результате стрессового воздействия как раз говорит в пользу аналогичного механизма. Поэтому в данном случае онтогенетический подход показывает, что, несмотря на отсутствие необратимых повреждений ФСА, растения листовой капусты в результате стресса претерпели определенные изменения своего состояния, которые мы в этой работе просто не зафиксировали явным образом.
Заключение. Таким образом, на основании полученных в работе результатов можно сделать два вывода. Первый заключается в том, что онтогенетический подход позволяет выявить произошедшие в растительном организме изменения даже в том случае, когда исследуемый показатель, например форма кривой ИФХ, лишь частично отражает такие изменения, в частности величину фотосинтетической ассимиляции СО2. Второй вывод состоит в необходимости коррекции существующего представления о протекании стрессового состояния у растительного организма как об ускоренном или замедленном онтогенезе, так как в ряде конкретных случаев реакция растений на стресс-фактор может не вписываться в представления, сформулированные подобным образом. С практической точки зрения, для исследований в области физиологии растений, экологии, растени- еводстве важно применять рассмотренный онтогенетический подход, не превращая его в формальный метод, а используя как один из элементов комплексного интегрального анализа состояния растений в различных условиях среды.
Список литературы Использование онтогенетического подхода при регистрации кривой Каутского в ходе стрессового воздействия на растения листовой капусты
- Lazar D. Chlorophyll a fluorescence induction//Biochim. Biophys. Acta 1999. -V. 1412. -P. 128.
- Корнеев Д.Ю. Информационные возможно-сти метода индукции флуоресценции хло-рофилла. -Киев: Алтерпресс, 2002. -188 с.
- Baker N.R. Chlorophyll Fluorescence a Probe of Photosynthesis in Vivo//Ann.Rev. Plant Biol. 2008. -V. 59. -pp. 89-113.
- Lichtenthaler H.K., Buschmann C., Knapp M. How to Correctly Determine the Different Chlo-rophyll Fluorescence Parameters and the Chlorophyll Fluorescence Decrease Ratio RFd of Leaves with the PAM Fluorometer//Photo-synthetica. -2005. -V. 43. -pp. 379-393.
- Roháček K. Chlorophyll fluorescence parame-ters: the definitions, photosynthetic meaning, and mutual relationships//Photosynthetica. -2002. -V. 40. -pp. 13-29.
- Карапетян Н.В., Бухов Н.Г. Переменная флуоресценция хлорофилла как показатель физиологического состояния растений//Фи-зиология растений. -1986. -Т. 33. -С. 1013-1036.
- Нестеренко Т.В., Сидько Ф.Я. Медленная индукция флуоресценции хлорофилла в онтогенезе двудольных растений (на при-мере листьев огурца)//Физиология расте-ний. -1986. -Т. 33. -С. 772783.
- Thomas H., Stoddart J.L. Leaf Senescence//Ann. Rev. Plant Physiol. -1980. -V. 31. -Р. 83-111.
- Цоглин Л.Н., Андреенко Т.И., Розанов В.В. и др. Фотосинтетический аппарат растений картофеля при длительном действии ви-русной инфекции//Физиология растений. -1987. -Т. 34, № 6. -С. 1103-1111.
- Nesterenko T.V., Tikhomirov A.A., Shikhov V.N. Ontogenetic approach to the assessment of plant resistance to prolonged stress using chlorophyll fluorescence induction method//Photosynthetica. -2006. -V.44, №. 3. -Р. 321-332.
- Шихов В.Н., Величко В.В., Нестеренко Т.В. и др. Онтогенетический подход при оценке методом индукции флуоресценции хлоро-филла реакции растений чуфы на условия культивирования//Физиология растений. -2011. -Т. 58, № 2. -С. 290-295.
- Manukovsky N.S., Kovalev V.S., Rygalov V.Ye. et al. Waste bioregeneration in life sup-port CES: development of soil organic sub-strate//Adv. Space Res. -1997. -V. 20, № 10. -Р. 1827-1832.
- Тихомиров А.А., Шихов В.Н. Зотин А.Г. и др. Обнаружение признаков деструктивного воздействия на высшие растения при по-мощи IP-камеры//Экологические системы и приборы. -2013. -№ 10 -С. 18-30.
- Воронин П.Ю., Рахманкулова З.Ф., Маев-ская С.Н. и др. Изменение фотосинтеза в результате адаптации проростков кукурузы к кратковременной засухе//Физиология растений. -2014. -Т. 61, № 1. -С. 143-147.
- Ireland C.R., Baker N.R., Long S.P. The role of carbon dioxide and oxygen in determining chlorophyll a fluorescence quenching during leaf development//Planta. -1985. -V. 165. -Р. 477-485.
