Использование органических остатков-индикаторов для уточнения палеогеографического положения двух террейнов Аляски в пределах Уральского морского пути в позднем силуре

Автор: Антошкина Анна Ивановна, Соджа К.М.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 2 (254), 2016 года.
Бесплатный доступ
Мы представляем обновленную модель для палеогеографии террейнов Аляски - Александра и Фаревел, показывая, как многочисленные палеонтологические и геологические данные ограничивают их местоположение в позднем силуре северным полушарием, а именно в пределах Уральского морского пути. Чрезвычайно выразительный микробиально-губково-гидроидный консорциум формировал довольно мощные рифы на окраинах платформ террейнов в позднем силуре (лудлове). Рифы и связанные с ними придонные сообщества показывают самое тесное палеобиогеографическое сходство с морской биотой западного и восточного склонов Урала и Салаира (юго-западная Сибирь). Изобилующие микробиальными сообществами рифы являются своего рода позднесилурийской «дактилоскопией» аномальных условий Уральского морского пути, которые были вызваны сокращающимся океанским бассейном, каледонской тектоникой и связанными с ними факторами в позднем силуре.
Уникальная рифовая биота, террейны александра и фаревел, урал, сибирь, позднесилурийская палеогеография
Короткий адрес: https://sciup.org/149129192
IDR: 149129192 | УДК: 551.83 | DOI: 10.19110/2221-1381-2016-2-14-23
Using fossil "fingerprints" to circumscribe the paleogeography of two Alaskan terranes along the Uralian seaway in the late Silurian
We present an updated model for the paleogeography of Alaska's Alexander and Farewell terranes, showing how a wealth of paleontologic and geologic data allow their locations to be circumscribed to the Northern Hemisphere-specifically the Uralian Seaway-in the Late Silurian. An unusually distinctive microbial-sponge-hydroid consortium constructed substantial platform-margin reefs in the terranes during the Late Silurian (Ludlow). The reefs and associated level-bottom communities exhibit strongest paleobiogeographic affinities with marine biotas found along the western and eastern slopes of the Urals and in Salair (southwestern Siberia). The microbe-dominated reefs are a Late Silurian «fingerprint» of abnormal Uralian Seaway conditions that were induced by a shrinking ocean basin, Caledonide tectonics, and related factors in the Late Silurian.
Текст научной статьи Использование органических остатков-индикаторов для уточнения палеогеографического положения двух террейнов Аляски в пределах Уральского морского пути в позднем силуре
Diagnostic Silurian species found at disparate sites in the Northern Hemisphere can be used to constrain the paleogeographic setting of «exotic» terranes now located in western North America (Alaska), southwestern Siberia, and the Urals. Together with other geologic evidence, these data circumscribe terrane locations to the Uralian Seaway. Specifically, extensive microbial-sponge reef communities of Late Silurian (Ludlow) age occur in the Alexander terrane (AT) and Farewell terrane (FT) of Alaska, in Salair in southwestern Siberia, and along the western and eastern slopes of the Urals. These reefs were built by an unusually distinctive consortium of microbial organisms (calcified cyanobacteria, calcareous algae, and microproblematica), sphinctozoan (aphrosalpingid) sponges, and Fistulella , a problematic hydroid. They are found nowhere else in the world during that restricted time interval.
We present below a brief overview of Northern Hemisphere paleogeography, the geologic setting of the 14
reef biotas, and their paleobiogeographic affinities. These data show that the microbe-dominated biotas record enormous ecological changes and unusual seaway conditions that prevailed across a broad region, bordered by Siberia to the north and by Laurentia and Baltica to the south [3, 53]. We hypothesize that narrowing of the Uralian Seaway and late-stage Caledonide (Scandian) tectonism generated significant terrestrial influx, reduced habitats for normalmarine metazoans, elevated nutrient loads, and, ultimately, fueled extensive microbial «blooms» that promoted stromatolite reef growth in the proto-Arctic basin in the Late Silurian.
Northern Hemisphere PaleogeographyContinents
In the Early–Mid Paleozoic, Laurentia and Baltica were clustered near the equator separated from Siberia to the north by the Uralian Seaway (Figure 1). During the
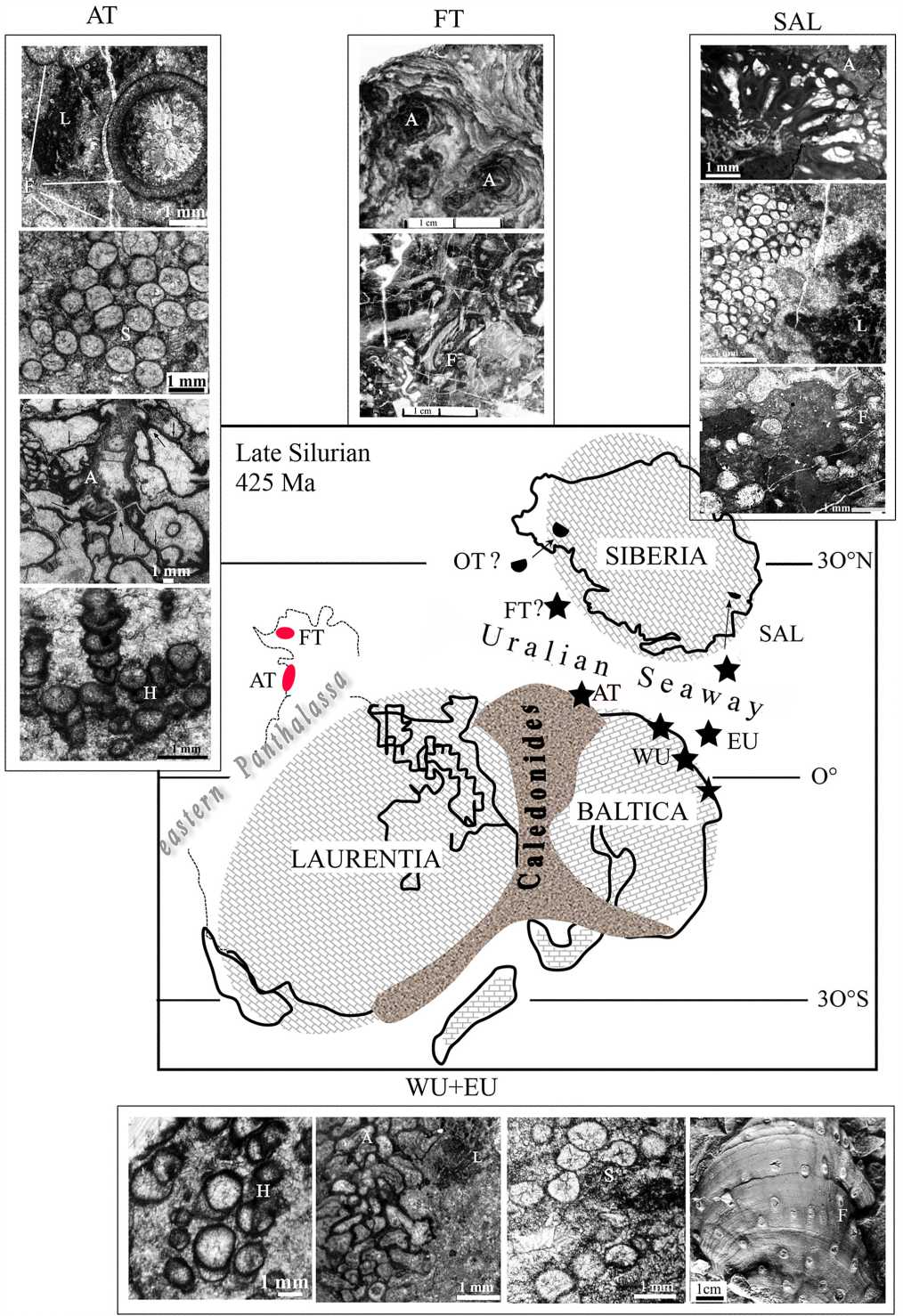
Figure 1. Our updated model for the paleogeography of Alaska’s Alexander (AT) and Farewell (FT) terranes along the Uralian Seaway, which also shows current locations of the AT and FT in Alaska and photomicrographs of representative reef biotas (stars indicate sites of co-occuring microbial-sponge-hydroid reef associations in Alaska, Salair, and the Urals). Modified from [3, 20, 30, 50]. Abbreviations: EU—Eastern Urals; OT — Omulevka terrane, SAL — Salair, WU — Western Urals. Photomicrograph abbreviations: A — aphrosalpingids, F — Fistulella hydroid, H — Hecetaphyton , L — Ludlovia , S — Sphaerina
Silurian, that seaway straddled the equator and was at its narrowest [30] with Siberia in close proximity to Baltica [19]. In the Late Silurian, Laurentia and Baltica ñollided during culminating phases of the Caledonide Orogeny. As Euramerica (Laurussia) was being consolidated, flyschmolasse sequences of the Old Red Sandstone (ORS) developed across 40° of latitude, extending 4500 km from eastern North America to Ireland, the UK, Norway, East Greenland, and Spitsbergen. The extraordinary thickness (4—8 km) of ORS sequences reflects high depositional rates in rapidly subsiding, intra-montane basins [9, 10, 19, 21, 25].
Until the onset of Uralide orogenesis, eastern Baltica (western slope of the present-day Ural Mountains) was a passive continental margin and unaffected by Caledonide orogenesis, as indicated by the absence of red bed molasse in the Late Silurian–Early Devonian. The Tagil and other volcanic arcs were accreted as terranes to Baltica, forming the eastern slope of the Urals beginning in the Late Devonian [36]. However, the pre-Carboniferous location of specific arc terranes that underwent accretion during the Uralide orogen is still debated.
Terranes
The Alexander terrane (AT) is one of the largest and best studied of the many terranes that form the North American Cordillera (all references in [50]). It originated in the Neoproterozoic–Early Paleozoic as an island arc and comprises early Precambrian detrital zircons that match the age of magmatic rocks found only in Baltica [26, 32]. Moreover, the Wales pre-Ordovi-cian orogenic event in the AT bears similarities with the Neoproterozoic–early Paleozoic Timanide orogen of eastern–northeastern Baltica [20]. Significantly, the AT is the only Cordilleran terrane that experienced a Late Silurian–Early Devonian orogeny (Klakas event); detrital zircons extracted from orogenic molasse (Upper Silurian– Lower Devonian Karheen Formation) best match those from western Baltica and the northern Caledonides [12, 13, 20, 26]. Finally, Silurian-Lower Devonian facies in the AT are very similar lithologically to coeval ORS deposits, which suggest that the arc-continent collision experienced by the AT during the Klakas event was related to Caledonide orogenesis [11—13, 55, 58]. Because its geologic history and fossil affinities are well established, the AT’s location along the mid-southern portion of the Uralian Seaway (Figure 1) serves as a linchpin for unraveling Northern Hemisphere paleogeography in the midPaleozoic, as described below.
The Farewell terrane (FT) of southwestern Alaska is a continental margin fragment, as indicated by Lower Paleozoic carbonate and siliciclastic shelf and slope deposits [20]. Early Paleozoic faunas share affinities with biotas in Siberia as well as Laurentia, Arctic Alaska–Chukotka terrane, and, to a lesser extent, the Omulevka terrane. This suggests that the terrane was derived from the Siberian (or Laurentian?) margin [14, 57]. Even though its mid-Paleozoic paleolatitude has yet to be established from paleomag-netic data, the FT was most likely located within 30° of the equator, given the thick carbonate deposits and abundance of warm-water indicators.
Debates persist about whether Salair, a Neoproterozoic–Early Paleozoic island arc, accreted to southwestern Siberia (present-day coordinates) in the early 16
Ordovician, subsequently forming part of a passive continental margin sequence, or if it remained an independent tectonic entity (micro-continent) before becoming welded to Siberia in the post-Silurian [23, 31, 60]. Thus, Salair’s paleogeography in the Late Silurian–Early Devonian remains open for discussion, although paleomagnetic data locate it within 30° of the equator in the mid-Paleozoic [59], as does paleontologic and sedimentologic evidence (discussed below).
Late Silurian Microbial Reefs
The AT is one of two Cordilleran terranes where an unusually distinctive consortium of microbial organisms (calcified cyanobacteria, calcareous algae, and micropro-blematica), sphinctozoan (aphrosalpingid) sponges, and Fistulella (a problematic hydroid) built enormous reefs in the Late Silurian (Ludlow) (Table 1). These microbe-dominated deposits do not represent level-bottom mat communities that grew in a marginal-marine or intertidal setting, as suggested by Cocks and Torsvik [19]. Rather, associated metazoans, the massiveness of the limestones, and the downslope deposition of megabreccias — some in the AT comprise house-sized blocks lithified by coarse rinds of marine carbonate cements — clearly indicate that the cal-ci-microbes constructed substantial stromatolite reefs at a platform margin [7, 45, 47, 56].
The FT is the other Cordilleran terrane where an extensive microbial reef complex developed in the Late Silurian-Early Devonian [18]. Diverse microbial taxa (e. g., Solenopora, Epiphyton, Renalcis, and Sphaerocodium ) constructed the buildups associated with dasycladaceans, gyp-idulinid brachiopods, sphinctozoan sponges (aphrosalpin-gids), problematic hydroids (e. g., Fistulella ), and rare corals and stromatoporoids. These deposits are approximately 500 m thick, 1 km wide, and several 100 km long. They interfinger with limestone debris flows and graptolitic shales of basinal origin.
The Ludlow reefs of the Urals formed in two different environmental settings: A) the West-Uralian reefs exposed for 2500 km were rigid organic structures that grew a passive platform at the eastern margin of Baltica; B) reefal limestones from the Eastern Urals originated in the Tagil island arc during a phase of waning volcanism. The Ludlow reefs are characterized by a similar, distinctive stroma-tolitic framework, encrustations of aphrosalpingid sponges on Fistulella [6, 7, 53], and relatively rare, fragmented remains of shelly marine benthos (brachiopods, coral, etc.) (Figure 1). In the Upper Silurian reefs of both regions, similar groups of organisms comprise the major reef-builders and reef-dwellers (Table 1). The co-occurrence of Aphrosalpinx textilis , Fistulella undosa , and Ludlovia multi-spora establishes a paleobiogeographic link between West-Uralian and East-Uralian environmental settings during the Late Silurian.
In northwestern Salair, Ludlow reefs described from the Potapovka Formation also exhibit a relatively low abundance of metazoans in a reef framework dominated by a similar consortium of microbial-stromatolitic taxa [3, 4].
Paleoenvironmentaland Paleogeographic Implications
Because the AT’s location is circumscribed to the Caledonide region in the Late Silurian-Early Devonian, the AT’s distinctive biotas shed insights into paleoen-vironmental conditions and paleogeographic relations that once existed in the proto-Arctic basin. In the Late Silurian, the Uralian Seaway was a narrow, equatorial– subequatorial marine corridor affected by Caledonide orogenesis along its southern margin [19, 50]. As the orogenic event reached its peak in the Late Silurian (Scandian phase), vast accumulations of Caledonide-derived sediment poured into the Uralian Seaway. Terrigenous sediment filled deep-marine troughs before flooding across shallow-marine platforms, terminating carbonate deposition to the south [19, 25]. We hypothesize that excessive nutrients, transported via terrigenous clastic runoff, fueled cyanobacterial «blooms» [50]. Unusual seawater chemistry and fluctuations in temperature, salinity, and oxygen concentrations could have also induced stressed environmental conditions [49].
The AT’s microbial reefs, which existed for a short time during culminating Caledonide (Scandian) phases in the Ludlow, clearly serve as a «fingerprint» of abnormal marine conditions that developed in the Uralian Seaway in the Late Silurian. Thus, we propose that similar, coeval biotas, such as those from the FT, Urals, and Salair, must have evolved at sites within that broad but contiguous marine basin, where Uralian Seaway conditions were conducive to the expansion of microbe-dominated communities. Percentages of taxa shared in common among coeval biotas are used to help locate more precisely each reef site and thus refine Northern Hemisphere paleogeography in the mid–Paleozoic.
Although not identical in taxonomic composition, the Alaskan reefs are most similar to coeval stromatolitesponge-hydroid reefs that developed in eastern Baltica [2, 7, 8, 47, 48]. The Uralian microbial reefs share >80 % of key taxa in common with the AT reefs (Table 1). Ludlow reefs in Salair also exhibit greatest similarities with the Uralian and AT reefs, sharing 61 % of taxa in common; far fewer of the reef taxa (22 %) co-occur in the FT [3, 4, 7] (Table 1). Similarly, the Ludlow microbial reefs in the AT and the Urals share only 25—30 % of taxa in common with the FT [48] (Table 1).
The paleontologic data indicate that Salair must have existed within the Uralian Seaway at a site between Baltica and Siberia that favored biotic exchange with the Tagil (EU) and Salair island arcs, AT, and, to a lesser extent, the FT in the Late Silurian (Figure 1). The greater similarities shared with the Uralian and AT biotas suggest that Salair was most likely located in the eastern portion of the seaway, either as an independent micro-continent or as part of southwestern Siberia (Table 1, Figure 1).
Because the FT shares some Ludlow reef biotas in common with Salair, the AT, and Urals, it too must have been located along the Uralian Seaway somewhere between Siberia, Baltica, Laurentia, and the AT [20, 50]. However, the absence of post-orogenic, flysch-molasse sequences in the FT comparable to the ORS indicates that the FT was not affected by the Caledonide orogeny and so must have resided at a significant distance from the orogenic front. Also, given the lower percentage of Silurian biotas in the FT that co-occur in the AT and Urals, the FT was most likely located along the northern margins of the Uralian Seaway at a site that lay beyond the effects of Caledonide tectonism and molasse deposition in the Late Silurian [49] (Table 1, Figure 1).
Additional support for our paleogeographic model is derived from a comprehensive compilation of all Silurian taxa described to date from the AT—from microbes to metazoans associated with reefs and level-bottom habitats (Table 1). Those data reveal that Silurian biotas in the AT do indeed exhibit strongest provincial affinities with Baltica, sharing 61 % in common. Significantly fewer AT taxa co-occur elsewhere, as indicated by the 20 % of taxa shared with Laurentia, 16 % with Salair, 12 % with the eastern Urals, and 11 % with the FT and also Siberia (Table 1). Thus, the paleontologic evidence is in full agreement with paleogeographic data derived from paleomagnetic and detrital zircon studies, which position the AT close to the Caledonide front [11—13, 20].
This multi-disciplinary perspective helps explain how similar Late Silurian marine communities could evolve in disparate tectonic settings: island arcs (AT, EU), passive continental margin (WU), and micro-continents (Salair, FT). Clearly, organisms that colonized those areas were adapted to the special conditions that arose during culminating phases of the Caledonide orogeny, as marine habitats were shrinking and receiving considerable sediment influx. Each biota acquired its own regional signature but was able to exchange larvae and undergo transmigration with other sites within the Uralian Seaway. Those in closer physical proximity or linked via effective ocean currents are characterized by a higher percentage of taxa shared in common (Figure 1).
During culminating orogenic phases in the Late Silurian-Early Devonian, the AT reefs collapsed as the shelf margin foundered, and AT biotas experienced regional extinction during marine regression and molasse progradation [55]. Elsewhere, however, abnormal conditions appear to have prevailed for millions of years at sites along the Uralian Seaway not overwhelmed by Caledonide-derived clastic influx. In particular, microbial-dominated reefs, associated in many areas with Fistulella , persisted into the Early Devonian along the western slope of the Urals, in the Tagil volcanic arc that would be accreted later to form the eastern Urals, and in the FT [5, 18]. The prolonged existence of massive reefs with a stromatolite-like, calcified framework, atypical for most Paleozoic reefs, is testament to the prevalence of unusual environmental conditions within the Uralian Seaway over a considerable interval of time [1, 2].
Finally, to test the hypothesis that the microbial-aphrosalpingid- Fistulella reefs are a Late Silurian «finger-print» for abnormal Uralian Seaway conditions in the Late Silurian, research was undertaken on coeval limestone deposits that formed in the Gobi-Altaj and Mandalovoo terranes in Mongolia. Those terranes were situated in the Northern Hemisphere within 35° of the paleoequator but beyond the eastern margin of the Uralian Seaway in the Paleozoic [35, 52]. The coeval rocks examined in Mongolia formed under environmental conditions broadly similar to those in the AT (i. e., in a spectrum of shallow- to deep-marine, limestone-dominated environments in a tectonically active setting). Yet calci-microbes do not predominate in the reefs or level-bottom communities there, and none of the distinctive microbial, aphrosalpingid, or hydroid taxa exist in the Gobi-Altai region [35, 52]. This helps confirm our hypothesis that unusual conditions along the Uralian Seaway—and nowhere else—favored the development of microbial reefs in the Late Silurian.
Table 1
Compilation of all key Silurian taxa described from the AT, their distribution in other sites, and their paleobiogeographic affinities, as indicated by the authors of the references shown
|
FORMATION |
SILURIAN TAXA (in AT) |
Authors (see references) interpret to be most like species from -> |
BAL |
LAU |
SAL |
EAS |
FAR |
SIB |
OT |
WNA |
NER |
LIV AT COS |
IND |
Affinities interpreted as |
As reported in |
||
|
authors' additional notes |
(endemic) |
||||||||||||||||
|
Level-bottom taxa are in white boxes Microbial reef taxa are in orange boxes |
|||||||||||||||||
|
Heceta |
KEY BRACHIOPODS (N=13) |
||||||||||||||||
|
Brooksina alaskensis |
X |
X |
X |
X |
Uralian-Cordilleran subprovince |
[14, 27, 42, 43] |
|||||||||||
|
Kirkidium vogulicum |
possible AT occurrence of morph. variable species* |
X |
X |
X |
(of Old World province) |
[*14, pg. 327] |
|||||||||||
|
Kirkidium cf. alaskense |
X |
| |
| |
||||||||||||||
|
Harpidium insignis |
X |
X |
X |
| |
| |
||||||||||||
|
Conchidium cf. alaskense |
X |
X |
| |
| |
|||||||||||||
|
Nanukidium cf. N. cunninghamensis |
X |
| |
| |
||||||||||||||
|
Atrypoidea borealis |
genus is widespread globally (Sil), common in Arctic Canada |
X |
X |
X |
| |
[28, 33] |
|||||||||||
|
| |
| |
||||||||||||||||
|
Gracianella |
genus is widespread globally in Silurian |
X |
X |
X |
X |
V |
[33] |
||||||||||
|
Alaskospira dunbari |
[16] |
||||||||||||||||
|
Cymbidium acutum |
X |
| |
|||||||||||||||
|
Atrypoidea tenuis |
V |
||||||||||||||||
|
Willoughby |
Sapelnikoviella santuccii |
new genus & species |
X |
[15] |
|||||||||||||
|
SIL/unknown |
Strophatrypa skaflestadi |
new genus & species |
X |
[17] |
|||||||||||||
|
(Kennel Creek |
|||||||||||||||||
|
see [29]) |
|||||||||||||||||
|
Heceta |
KEY GASTROPODS (N=22) |
||||||||||||||||
|
(& Willoughby) |
Bathmopterus liratus |
endemic genus & species |
X |
[14, 24, 40] |
|||||||||||||
|
Beraunia bohemica |
species occurs in Late Silurian of Bohemia |
X |
| |
||||||||||||||
|
Beraunia bifrons |
appears identical to species from Late Silurian, Bohemia |
X |
| |
||||||||||||||
|
Hecetastoma gehrelsi |
new genus & species |
X |
| |
||||||||||||||
|
Coelocaulus karlae |
resembles some Silurian species of Coelocaulus from |
X |
| |
||||||||||||||
|
Prague Basin; genus is known from eastern NA (mid-late Sil) |
| |
||||||||||||||||
|
Baichtalia tongassensis |
new genus & species |
X |
| |
||||||||||||||
|
Kirkospira glacialis |
new genus & species; similar to “ Pleurotomaria” |
X |
| |
||||||||||||||
|
lindströmi from Urals; genus is known from eastern NA |
| |
||||||||||||||||
|
Goniostropha |
X |
[14] |
|||||||||||||||
|
Medfracaulus turriformis |
appears identical to M. turriformis from the Urals |
X |
X |
X |
| |
||||||||||||
|
(+ genus is known from SIB) |
X |
| |
|||||||||||||||
|
Pachystrophia gotlandica |
Euomphalus gotlandicus from Wenlock of Gotland is |
X |
(X) |
X |
| |
||||||||||||
|
very similar in size and shape |
| |
||||||||||||||||
|
Retispira aff. R. volgulica |
very similar in size, shape, ornament to Ludlow |
X |
| |
||||||||||||||
|
“Bellerophon” volgulicus from Urals |
| |
||||||||||||||||
|
Pachystrophia cf . gotlandica |
(+ genus is known from Gondwana terrane) |
X |
close relationship with gastropod |
[41] |
|||||||||||||
|
Spinicharybdis krizi |
genus is known from the E Sil (Gotland) & Late Silurian |
X |
X |
faunas of Bohemia & FAR |
[33] |
||||||||||||
|
Spinicharybdis boucoti |
(Ludlow) of Canadian Arctic (+ Kazakhstan) |
X |
| |
| |
|||||||||||||
|
Haplospira craigi |
H. spinosum from the Silurian (Ludlovian) Kopanina Fm |
X |
| |
| |
|||||||||||||
|
of Czech Republic |
| |
| |
|||||||||||||||
|
Australonema sp. |
Gotland; Siberia (Sil); Bohemia (Dev); Australia (Dev) |
X |
X |
| |
| |
||||||||||||
|
Morania wagneri |
Bohemia, Scotland, eastern Canada |
X |
X |
| |
| |
||||||||||||
|
Morania nixonforkensis |
(genus is known from SIB + Gondwana terrane) |
X |
X |
| |
| |
||||||||||||
|
Medfrazyga gilmulli |
Medfrazyga clauticae from Dev of southwestern AK |
X |
V |
V |
|||||||||||||
|
FORMATION SILURIAN TAXA (in AT) Authors (see references) interpret to be most like species from -> BAL LAU SAL EAS FAR SIB OT WNA NER LIV AT COS IND Affinities interpreted as |
As reported in |
||||||||||||||||
|
Bucanopsis |
(also known from eastern NA) |
X |
X |
closest Silurian gastropod faunas |
[24, pg. 214] |
||||||||||||
|
Oriostoma |
(also known from eastern NA + Gondwana terrane) |
X |
X |
to those from the AT are known |
| |
||||||||||||
|
Euomphalopterus |
(also known from eastern NA + western Avalonia) |
X |
X |
X |
from the Urals, Bohemia, & FAR |
V |
|||||||||||
|
Heceta |
KEY BIVALVES (N=11) |
||||||||||||||||
|
& Willoughby |
Pycinodesma benjamini |
similar, coeval megalodontids known |
(X) |
(X) |
X |
[14, 22] |
|||||||||||
|
Pycinodesma giganteum |
mainly from Canadian Arctic + Urals |
X |
|||||||||||||||
|
SIL/Kennel |
? Colpomya aff. hugini |
Gotland, Sweden |
X |
Janicula potens community |
[29] |
||||||||||||
|
Creek |
? Colpomya aff. audae |
Prague Basin, Bohemia |
X |
Gotland & Prague basin, |
| |
||||||||||||
|
Modiolopsis aff. modiolaris |
Prague Basin, Bohemia (Ordovician) |
X |
Bohemia |
| |
|||||||||||||
|
Mytilarca boucoti |
Prague Basin, Bohemia (Ordovician) |
X |
| |
| |
|||||||||||||
|
? Schizodus sp . A |
China (Devonian) |
| |
| |
||||||||||||||
|
? Schizodus sp. B |
central and eastern US, Laurentia (Devonian) |
X |
| |
| |
|||||||||||||
|
Saguinolites sp . |
Prague Basin, Bohemia; China (Devonian) |
X |
| |
| |
|||||||||||||
|
Modiomorpha sp. |
X |
| |
| |
||||||||||||||
|
Goniophora thula |
Prague Basin, Bohemia |
X |
V |
V |
|||||||||||||
|
Heceta |
SCAPHOPODS (N=2) |
||||||||||||||||
|
Dentalium hecetaensis |
X |
[41] |
|||||||||||||||
|
Rhytiodentalium cf. kentuckyensis |
[41] |
||||||||||||||||
|
Heceta |
TRILOBITES (N=1) |
||||||||||||||||
|
Richterarges n. sp. |
R. aquilonius, R. bigener from Canadian Arctic |
X |
[14] |
||||||||||||||
|
Heceta |
KEY SPONGES (N=15) Aphrosalpinx textilis Aphrosalpinx nana Nematosalpinx dichotomica Nematosalpinx hormathodes Cystothalamiella alaskensis Cystothalamiella irregularis Palaeoschada crassimularlis Girtyocoeliana epiporata |
X |
X |
X |
X |
X |
strongest affinities with |
[ 7, 14, 38, 39, 50] |
|||||||||
|
X? XXX X X X |
BAL, SAL, & FAR |
| |
|||||||||||||||
|
| |
|||||||||||||||||
|
| |
|||||||||||||||||
|
| |
|||||||||||||||||
|
| |
|||||||||||||||||
|
— |
X |
X |
X |
unstated, but highly endemic |
V [39] |
||||||||||||
|
Alaskaspongiella laminosa |
new genus & species |
X |
fauna comprises many new |
| |
|||||||||||||
|
Polyplacospongia nodosa |
new genus & species |
X |
genera and species |
| |
|||||||||||||
|
Monolaminospongia gigantia |
new genus & species |
X |
| |
| |
|||||||||||||
|
Cladospongia alaskensis |
new genus & species |
X |
| |
| |
|||||||||||||
|
Virgulaspongia uniforma |
new genus & species |
X |
| |
| |
|||||||||||||
|
Stipespongia laminata |
new genus & species |
X |
| |
| |
|||||||||||||
|
Turbospongia biperforata |
new genus & species |
X |
V |
V |
|||||||||||||
|
SIL/Descon |
GRAPTOLITES |
X |
[14] |
||||||||||||||
|
Heceta |
CONODONTS |
X |
[14] |
||||||||||||||
|
Heceta |
RUGOSE CORALS (N=9) |
||||||||||||||||
|
Tryplasma spp. |
U Sil-E Dev rugosids |
[34] |
|||||||||||||||
|
Cystiphyllum spp. |
(along with detrital zircons) |
| |
|||||||||||||||
|
FORMATION |
SILURIAN TAXA (in AT) |
Authors (see references) interpret to be most like species from -> |
BAL LAU SAL |
EAS FAR |
SIB |
OT |
WNA |
NER |
LIV |
AT |
COS |
IND |
Affinities interpreted as |
As reported in |
||||
|
new cystiphyllid |
indicate the AT was in |
| |
||||||||||||||||
|
Microplasma sp. |
the northern hemisphere at |
| |
||||||||||||||||
|
Rhizophyllum sp. A |
longitude near BAL and in Late |
| |
||||||||||||||||
|
Zelophyllum sp . |
Devonian-Early Carboniferous |
| |
||||||||||||||||
|
Plasmophyllum labrum n. sp. |
X |
was close to w. Canada |
| |
|||||||||||||||
|
Plasmophyllum sp. |
X |
X |
| |
| |
||||||||||||||
|
new kyphophyllids |
V |
V |
||||||||||||||||
|
Heceta |
?HYDROID (N=1) |
|||||||||||||||||
|
& Willoughby |
Fistulella undosa |
X |
X |
X |
X |
[2, 3, 7, 53] |
||||||||||||
|
Heceta |
MICROBIOTAS (N=14) |
|||||||||||||||||
|
& Willoughby |
Girvanella problematica |
X |
X |
strongest affinities with |
[37, 54] |
|||||||||||||
|
Girvanella sp. |
BAL, SAL, & FAR |
[3, 7, 50] |
||||||||||||||||
|
Hedstroemi a sp. |
X |
X |
| |
| |
||||||||||||||
|
Epiphyto n sp. |
X |
X |
X |
X |
| |
| |
||||||||||||
|
Hecetaphyton alaskense |
new genus & species |
X |
X |
| |
| |
|||||||||||||
|
Ludlovia multispora |
X |
X |
X |
| |
[42] |
|||||||||||||
|
Renalcis cf. R. tuberculatus |
X |
X |
X |
| |
| |
|||||||||||||
|
"Solenopora" compacta |
X |
| |
| |
|||||||||||||||
|
"Solenopora" filiformis |
X |
| |
| |
|||||||||||||||
|
"Solenopora" gotlandica |
X |
| |
| |
|||||||||||||||
|
Rothpletzella gotlandica |
X |
| |
| |
|||||||||||||||
|
Tuxekanella simplex |
new genus & species |
X |
| |
[44] |
||||||||||||||
|
Wetheredella silurica |
X |
X |
| |
| |
||||||||||||||
|
Sphaerina congregata |
new genus & species |
X |
X |
V |
V |
|||||||||||||
|
Heceta |
ICHNOGENERA (N=5) |
|||||||||||||||||
|
Chondrites ichnosp. |
X |
none suggested |
[46] |
|||||||||||||||
|
?Chondrites ichnosp. |
X |
| |
| |
|||||||||||||||
|
Palaeophycus ichnosp. |
X |
| |
| |
|||||||||||||||
|
Planolites ichnosp. cf. P. beverleyensis |
X |
| |
| |
|||||||||||||||
|
?Planolites ichnosp. |
X |
V |
V |
|||||||||||||||
|
QUALITATIVE TALLY |
42 |
15 |
12 |
9 |
8 |
8 |
5 |
3 |
0 |
0 |
19 |
4 |
6 |
|||||
|
note: after completing compilation, columns |
were re-arranged to show strongest to weakest faunal affinities from left to right |
|||||||||||||||||
|
TOTAL # AT TA |
93 |
|||||||||||||||||
|
Total # AT ende |
19 |
|||||||||||||||||
|
Total # AT non- |
74 |
endemics= |
||||||||||||||||
|
% of 74 non-endemic AT taxa found in other geographic sites -> |
61 |
20 |
16 |
12 |
11 |
11 |
7 |
4 |
0 |
0 |
20% |
5 |
8 |
|||||
|
BAL LAU SAL |
EAS |
FAR |
SIB |
OT |
WNA NER |
LIV |
AT |
COS |
IND |
|||||||||
|
Abbreviations: AT (Alexander terrane); BAL (Baltica); COS (cosmopolitan); EAS (eastern Urals=accreted island-arc terranes; Antoshkina & Königshof 2008); FAR (Farewell terrane of southwestern Alaska); |
||||||||||||||||||
|
IND (indeterminate); LAU (Laurentia); LIV (Livengood terrane of east-central Alaska); NER (northeast Russia); OT (Omulevka terrane); SAL (Salair); SIB (Siberia); WNA (Cordilleran terranes in western North America) |
||||||||||||||||||
Conclusions
Our updated paleogeographic reconstruction of the Northern Hemisphere in the Late Silurian is based on an analysis of unique reef biotas and associated communities described from the AT, FT, Salair, and Urals. Those reefs, dominated by microbial rather than metazoan taxa, are diagnostic indicators of the stressed environmental conditions that developed along the Uralian Seaway in the Late Silurian during culminating phases in the Caledonide orogeny. An integration of paleogeographic (paleomagnet-ic, detrital zircon) and paleobiogeographic (paleontologic) evidence allows the AT to serve as a linchpin for identifying the Northern Hemisphere position of some accreted terranes and micro-continents in the mid-Paleozoic.
Acknowledgments
We are grateful to the National Science Foundation of USA and to many colleagues at Colgate University, Smith College, Institute of Geology (Syktyvkar), Institute of Geology and Geophysics (Ekaterinburg), Kuzbass State Pedagogical Academy (Novokuznetsk), and Mongolian University of Science and Technology (Ulaanbaatar) who provided support, help, guidance, and advice in the field. Additional thanks go to Viktor N. Puchkov for constructive comments on a final version of this paper.
Список литературы Использование органических остатков-индикаторов для уточнения палеогеографического положения двух террейнов Аляски в пределах Уральского морского пути в позднем силуре
- Antoshkina A. I. 1998. Organic buildups and reefs on the Palaeozoic carbonate platform margin, Pechora Urals, Russia. Sedimentary Geology, V. 108 (1-4), pp. 187-210.
- Antoshkina A. I. 2003. Rifoobrazovanie v Paleozoe (Sever Urala i sopredelnye oblasti) (Reef formation in Paleozoic (Northern Urals and adjacent areas)) UrO RAN, Ekaterinburg. 303 pp.
- Antoshkina A. I. 2005. Rifovaya biota Urala i Salaira: Paleobiogeograficheskoe znachenie dlya srednego Paleozoya (Reef biota of Urals and Salair: Paleobiogeographical value for Middle Paleozoic). Komi Nauchnyj Tsentr, Uralskoe Otdelenie, Rossijskaya akademiya nauk. Nauchnye doklady. V. 469, pp. 1-26.
- Antoshkina A. I., Gutak Y. M., Bagmet G. N., and Savistkij V. R. 2006. Novye dannye po silurijskim rifam Salaira, Zapadnaya Sibir (New data on Silurian reef of Salair, Western Siberia). Novosti Paleontologii i Stratigrafii. V. 9, pp. 21-29.
- Antoshkina A. I., Königshof P. 2008. Lower Devonian reef structures in Russia: an example from the Urals. Facies. V. 54, pp. 233-251.
