Исследование действия ионизирующего излучения на радиационно- индуцируемые изменения клеток системы крови мышей на уровне организма

Автор: Когарко И.Н., Петушкова В.В., Когарко Б.С., Пряхин Е.А., Нейфах Е.А., Ктиторова О.В., Андреев С.С., Ганеев И.И., Кузьмина Н.С., Селиванова Е.И., Пелевина И.И.
Рубрика: Научные статьи
Статья в выпуске: 1 т.32, 2023 года.
Бесплатный доступ
В работе была поставлена задача изучения радиационных «эффектов свидетеля» на уровне организма. В эксперименте были использованы облучённые и необлучённые мыши, которые содержались совместно. Мыши были облучены в дозе 3 Гр на исследовательской радиобиологической гамма-установке с источниками 137Cs. Оценивали количество лейкоцитов и относительное количество лимфоцитов в периферической крови у мышей через 3, 7, 14, 30, 60 и 90 сут после начала эксперимента. У необлучённых мышей-«свидетелей», содержавшихся с облучёнными мышами как в клетках без перегородки, так и с перегородкой, прослеживается возможная тенденция к уменьшению числа лейкоцитов по сравнению с показателями в биоконтроле. При анализе относительного количества лимфоцитов показана тенденция к снижению показателя у мышей-«свидетелей» как при содержании без перегородки, так и с перегородкой. У необлучённых животных-«свидетелей» при содержании с облучёнными животными в клетке с перегородкой было зарегистрировано статистически значимое снижение относительного количества лимфоцитов в периферической крови: на 3-и сут (t=2,13; p=0,047) и 30-е сут (t=2,94; p=0,01) и на 90-е сут (t=3,07; p=0,01) после облучения, а также при содержании в одной клетке с облучёнными мышами без перегородки на 60-е сут (t=2,24; p=0,038) после начала эксперимента. Одним из возможных объяснений выявленных изменений может быть «эффект свидетеля» у необлучённых животных. У облучённых животных, которых содержали в одной клетке с перегородкой вместе с необлучёнными животными, было зарегистрировано статистически значимое повышение относительного количества лимфоцитов на 3-и (t=2,6; p=0,02), 14-е (t=2,61; p=0,018) и 60-е сут (t=2,38; p=0,03) - «эффект спасения». На основании полученных в настоящем эксперименте данных делается предположение, что радиационный «эффект свидетеля» возможно имеет обратный характер, то есть необлучённые организмы способны снижать радиационные эффекты у облучённых особей - «эффект спасения».
Ионизирующее излучение, «немишенные эффекты», структурно-функциональные нарушения генома, функциональные изменения системы крови, радиационно-индуцированный «эффект свидетеля», радиационно-индуцированный «эффект спасения», лимфоциты, лейкоциты, беспородные мыши-самки, межорганизменные взаимодействия, многофакторный анализ полученных данных.
Короткий адрес: https://sciup.org/170198088
IDR: 170198088 | УДК: 577.346:591.11:599.323.4 | DOI: 10.21870/0131-3878-2023-32-1-48-60
Investigation of the effects of ionizing radiation on radiation-induced bystander changes in the cells of the blood system of mice at the inter-organism level
The study was aimed at the task to determine the radiation-induced “bystander effects” at the inter-organism level. In the experiment, irradiated and non-irradiated mice kept together were used. The mice were irradiated at a dose of 3 Gy at a research radiobiological gamma-ray facility with sources of 137Cs. The number of leukocytes and the relative number of lymphocytes in peripheral blood of mice were evaluated at 3, 7, 14, 30, 60 and 90 days after the irradiation. In unirradiated “bystander” mice kept with irradiated animals, both in cages without a septum and with a septum, a possible tendency to decrease the number of leukocytes compared to the same indicators in biological control was found. When analyzing the relative number of lymphocytes, a tendency to decrease in this indicator in “bystander” mice both when kept without a partition and with a partition was shown. In unirradiated bystander animals when kept with irradiated animals in a cage with a partition, a statistically significant decrease in the relative number of lymphocytes in peripheral blood was recorded: at the 3-rd day (t=2.13; p=0.047), at the 30-th day (t=2.94; p=0.01) and at the 90-th day (t=3.07; p=0.01) after irradiation, as well as when kept in the same cage with irradiated mice without a partition at the 60-th day (t=2.24; p=0.038) after the start of the experiment. This result can be considered as a radiation-induced “bystander effect” in unirradiated animals. In irradiated animals, which were kept together with unirradiated animals in the cage with a partition, a statistically significant increase in the relative number of lymphocytes was recorded at the 3-rd day (t=2.6; p=0.02), 14-th day (t=2.61; p=0.018) and at the 60-th day (t=2.38; p=0.03) after irradiation, so the radiation-induced “rescue effect” was obtained. Based on the data obtained in this experiment, it is assumed that the radiation-induced “bystander effect” may be reversed, that is, unirradiated organisms are able to reduce radiation-induced effects in irradiated individuals - the “rescue effect”.
Текст научной статьи Исследование действия ионизирующего излучения на радиационно- индуцируемые изменения клеток системы крови мышей на уровне организма
В настоящее время в ряде работ [1-3] показано, что ионизирующая радиация вызывает широкий спектр изменений в клеточном составе периферической крови животных (мышей). Показано, что биологические эффекты в клетках после действия ионизирующего излучения (ИИ) обусловлены повреждением ядерной ДНК, нарушениями в системе репарации ДНК, что реализуется в повышении хромосомных аберраций и индукции других неблагоприятных эффектов. У клеток,
Когарко И.Н. – вед. науч. сотр., д.м.н.; Петушкова В.В .* – вед. спец., к.э.н.; Когарко Б.С. – ст. науч. сотр., к.ф.-м.н.; Нейфах Е.А. – ст. науч. сотр., к.б.н.; Ктиторова О.В. – науч. сотр., к.б.н.; Ганеев И.И. – инж.-исслед.; Пелевина И.И. – д.б.н. ФГБУН ФИЦ ХФ РАН. Пряхин Е.А. – зав. отд., д.б.н.; Андреев С.С. – ст. науч. сотр., к.б.н. ФГБУН УНПЦ РМ ФМБА России. Кузьмина Н.С. – ст. науч. сотр., д.б.н. ФГБУН ФИЦ ХФ РАН, ФГБУН ИОГен РАН. Селиванова Е.И. – ст. науч. сотр., к.б.н. ФГБУН ФИЦ ХФ РАН, МРНЦ им. А.Ф. Цыба – филиал «НМИЦ радиологии» Минздрава России.
которые находятся рядом с облучёнными клетками или в культуральной среде от облучённых клеток, также могут наблюдаться изменения, схожие с обнаруженными в облучённых клетках.
Так называемые «немишенные эффекты» ИИ, включая «эффекты свидетеля», изучены in vitro и in vivo [4, 5]. Показано, что «эффекты свидетеля» характеризуются не только переносом повреждений от облучённых клеток к необлучённым в виде хромосомных аберраций, генных мутаций, изменения экспрессии генов, но и апоптозом, неопластической трансформацией клеток-«свидетелей».
Существование радиационно-индуцированного «эффекта свидетеля», при котором необлу-чённые клетки проявляют реакции, сходные с характерными для облучённых при воздействии облучённых клеток или кондиционированной среды от облучённых клеток, было рассмотрено в тех же работах Coates et al. (2004) [4], Matsumoto et al. (2004) [5] и другими. Ранее считалось, что «эффект свидетеля» может реализовываться только в отдельных клетках в пределах одного органа или системы. Недавно было показано, что «эффект свидетеля» может быть зарегистрирован не только в клеточных культурах или in vivo в одном организме, но и на межорганизменном уровне, облучённый организм может индуцировать изменения у необлучённого организма [6-9].
Дистанционный «эффект свидетеля» - наименее изученный с точки зрения проявлений и механизмов феномен, относящийся к «немишенным эффектам». В рамках комплексного проекта по оценке радиационно-индуцированных «немишенных эффектов» в работах [10-15] приведены результаты наших исследований. В нашем предыдущем эксперименте [10, 11] у необлучённых мышей, находившихся на протяжении 3 мес. в контакте с облучёнными (3 Гр) особями, выявлены признаки изменений, сходных с радиационно-индуцированными. Отмечались изменение поведенческих реакций, статистически значимая тенденция к уменьшению веса селезёнки (r=-0,416; p=0,048), повышению площади алопеции (r=-0,631; p=0,001) и иные аномалии.
В другой работе [13] было показано, что у необлучённых мышей при их совместном содержании с облучёнными животными вне зависимости от наличия или отсутствия перегородки повышение частоты нормальных хроматофильных (оксифильных) эритроцитов с микроядрами по сравнению с показателем в биологическом контроле, которые можно было интерпретировать как «эффект свидетеля». У облучённых животных получен так называемый «эффект спасения» на тех же эритроцитах разной степени зрелости в периферической крови у мышей.
В работе [14] был проведён анализ состояния кроветворных органов (костный мозг, селезёнка и тимус), включивший в себя исследование количества ядерных клеток в них, а также оценку массы селезёнки и тимуса. Полученные результаты говорят о наличии «эффекта свидетеля» в ядросодержащих клетках костного мозга мышей.
Целью настоящего исследования является изучение радиационных «эффектов свидетеля» и «эффектов спасения» на уровне организма у облучённых и необлучённых животных при их совместном содержании при использовании лейкоцитов и лимфоцитов периферической крови мышей.
Материалы и методы
Исследование выполнено на 60 беспородных мышах-самках. В опыте использовали здоровых животных, не подвергавшихся ранее другим экспериментальным воздействиям. Группы формировали методом сплошной выборки. При проведении экспериментальных процедур соблюдались соответствующие международные правила работы с животными [16, 17]. Животные (n=20) были облучены в дозе 3 Гр на исследовательской радиобиологической гамма-установке
ИГУР-1М с четырьмя источниками 137Cs, мощность дозы 0,79 Гр/мин, неравномерность γ-поля в рабочем пространстве не более 5%. Поглощённая доза для мышей составила 3,0 Гр.
Мыши были рассажены в четыре клетки, в каждой из которых находилось по пять облучённых мышей и по пять необлучённых (мыши-«свидетели»). В двух из четырёх клеток облучённые и необлучённые мыши могли свободно контактировать друг с другом. В двух других клетках мыши облучённой и необлучённой групп были разделены металлической сеткой, что исключало их тактильный контакт. Для контроля были использованы: группа биоконтроля (n=10) и группа облучённых животных (n=10), каждая из которых содержалась в отдельной клетке. Было получено 6 экспериментальных групп. Эксперимент продолжался 3 мес.
Кровь из хвостовой вены мышей отбирали на 3-и, 7-е, 14-е, 30-е, 60-е и 90-е сут после начала эксперимента. Кровь в объёме 8 мкл использовали для подсчёта количества лейкоцитов с помощью гемоцитометра Пикоскель PS-4. Для анализа формулы крови подсчитывали 200 лейкоцитов.
Статистический анализ проводили с помощью t-критерия Стьюдента. Различия принимали статистически значимыми при вероятности 0-гипотезы р<0,05. Для оценки влияния различных факторов на анализируемые показатели проводили многофакторный дисперсионный анализ в обобщённой линейной модели. Оценивали влияние следующих воздействующих факторов: гамма-облучение; срок после начала эксперимента; «фактор контакта» – вид контакта между облучёнными и необлучёнными животным (0 – отсутствие контакта между облучёнными и необлу-чёнными животными; 1 – обонятельный контакт (с перегородкой); 2 – тактильный и обонятельный контакт (без перегородки).
Результаты
Известно, что острое гамма-облучение приводит к снижению количества лейкоцитов в периферической крови у млекопитающих. В представленных экспериментах после острого гамма-облучения в дозе 3 Гр уже через 3 сут в периферической крови регистрировалось статистически значимое снижение количества лейкоцитов более чем в 5 раз (рис. 1). С увеличением времени после облучения происходило восстановление количества лейкоцитов в периферической крови и на 60е сут этот показатель практически не отличался у облучённых животных по сравнению с показателем в группе биологического контроля. Следовательно, облучение приводит к снижению количества лейкоцитов в периферической крови, показатель повышается в зависимости от времени после начала эксперимента, что отражает его пострадиационное восстановление.
У необлучённых животных-«свидетелей» не было выявлено статистически значимых отклонений от группы биологического контроля, однако при совместном пребывании с облучёнными животными у необлучённых животных в клетке без перегородки проявляется тенденция к относительному уменьшению числа лейкоцитов на 3-и сут и в отдалённый период времени на 30-е и 60-е сут после облучения (рис. 2). При наличии в клетке перегородки также наблюдается относительное уменьшение числа лейкоцитов на 3-и, 14-е, 30-е, а также 90-е сут. Этот результат, по-видимому, можно считать «эффектом свидетеля» у необлучённых животных, показанном на графике (рис. 2), где линия, соответствующая результатам «эффекта свидетеля», находится ниже линии контроля.
Результат, показанный на рис. 3, по-видимому, можно считать «эффектом свидетеля» у необлучённых животных. У облучённых животных, которых содержали вместе с необлучёнными животными-«свидетелями», было зарегистрировано по сравнению с гамма-контролем снижение количества лейкоцитов при совместном нахождении с необлучёнными животными в клетке с перегородкой на 3-и, 7-е, 14-е, 60-е, а также 90-е сут, а при содержании без перегородки на 3-и, 7-е, 14-е и 60-е сут.
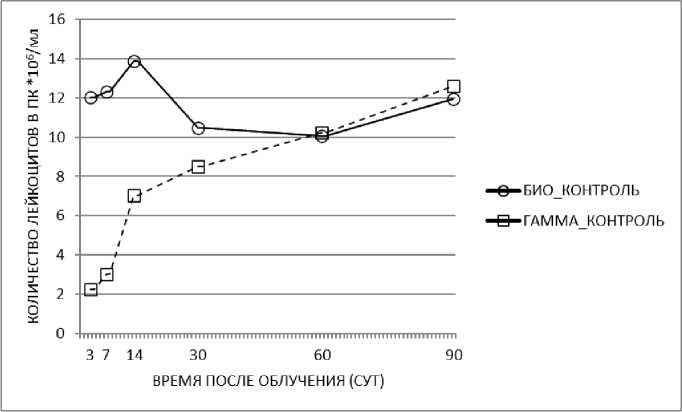
Рис. 1. Изменение количества лейкоцитов после острого гамма-облучения мышей в дозе 3 Гр через 3 сут.
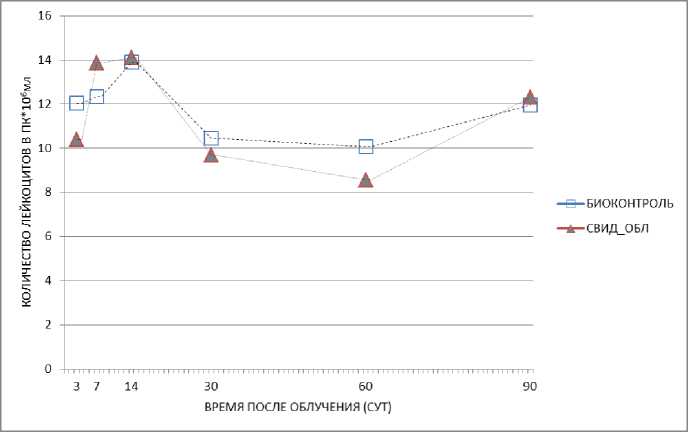
Рис. 2. Количество лимфоцитов в динамике у необлучённых мышей при совместном пребывании с облучёнными мышами.
Проведение многофакторного анализа выявило статистически значимое влияние факторов «День» (F=8,05; p<0,001) и фактора «Облучение» (F=14,10; p<0,001). Как видно из табл. 1, коэффициент для фактора «Облучение» показывает, что в среднем гамма-облучение приводит к снижению количества лейкоцитов на 4,7 млн лейкоцитов/мл. Как видно из табл. 1, при обонятельном контакте достоверно снижалось количество лейкоцитов в периферической крови в среднем на 0,89 млн лейкоцитов/мл (t=2,17; p=0,031), при этом [Фактор контакта=2], соответствующий тактильному и обонятельному взаимодействию облучённых и необлучённых животных, не оказывал статистически значимого влияния (t=1,45; p=0,15); B – свободный член.
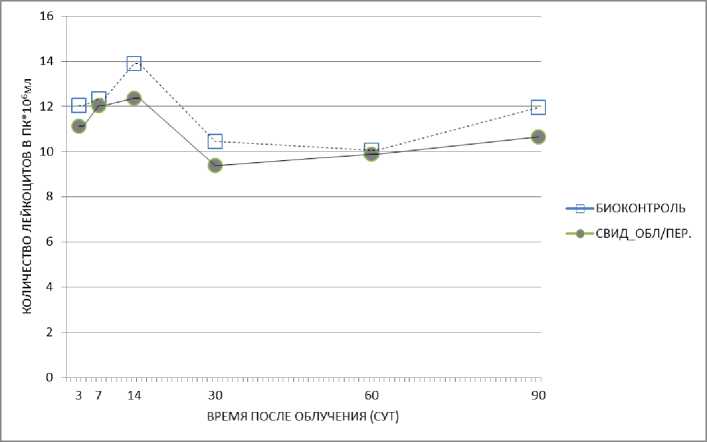
Рис. 3. Количество лимфоцитов в динамике у необлучённых мышей при наличии перегородки между ними и облучёнными мышами.
Таблица 1
Параметры обобщённой линейной модели при анализе влияния различных факторов на количество лейкоцитов в периферической крови у мышей
|
Параметр |
B |
t |
р |
|
Свободный член |
5,72 |
15,08 |
4,8x10-4° |
|
[Фактор контакта=0] |
0,00 |
||
|
[Фактор контакта=1] |
-0,89 |
-2,17 |
0,031 |
|
[Фактор контакта=2] |
-0,59 |
-1,45 |
0,150 |
|
День |
0,04 |
8,05 |
1,2x10-14 |
|
[Облучение=0] |
0 |
||
|
[Облучение=1] |
-4,71 |
-14,10 |
3,6x10-36 |
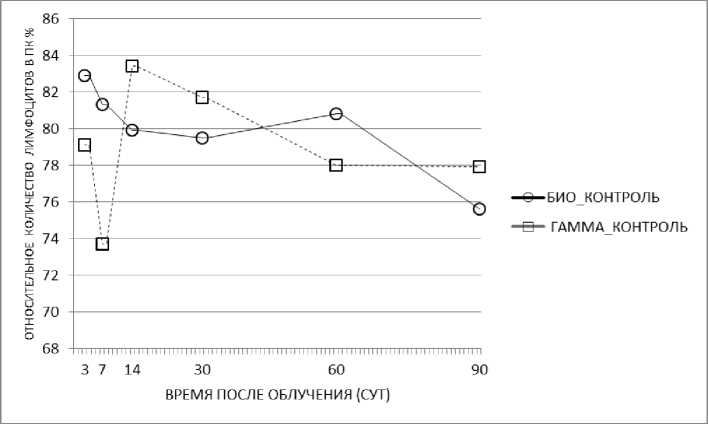
Рис. 4. Относительное количество лимфоцитов периферической крови в динамике после гамма-облучения мышей.
При анализе клеточного состава периферической крови было выявлено, что доля лимфоцитов в группе биологического контроля во время всего эксперимента изменялась незначительно и находилась в диапазоне от 76 до 83%. Гамма-облучение вызывало временное снижение количества лимфоцитов на 7-е сут после облучения (рис. 4). В дальнейшем, на 30-е, 60-е и 90-е сут после облучения не было выявлено статистически значимого снижения доли лимфоцитов в периферической крови.
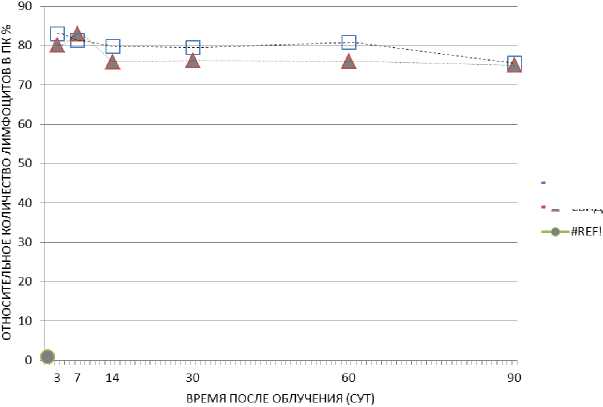
У необлучённых животных-«свидетелей» было зарегистрировано снижение относительного количества лимфоцитов в периферической крови на 3-и, 14-е, 30-е и 60-е сут после начала эксперимента при содержании в одной клетке с облучёнными мышами без перегородки (рис. 5). Такое же снижение относительного количества лимфоцитов в периферической крови мышей было отмечено на 3-и, 7-е, 30-е и 90-е сут при содержании с облучёнными животными с перегородкой (рис. 6). По-видимому, этот результат можно считать «эффектом свидетеля» у необлу-чённых животных, представленном на графике, где линия, отвечающая за «эффект свидетеля», расположена ниже контрольной линии. Необходимо отметить также, что относительное число лимфоцитов периферической крови у необлучённых животных-«свидетелей», содержавшихся с перегородкой, на 90-е сут после облучения резко понизилось с 78,2 до 67%.
У облучённых мышей, которых содержали с необлучёнными животными-«свидетелями» в одной клетке без перегородки, по сравнению с показателем в группе облучённого контроля было зарегистрировано повышение доли лимфоцитов в периферической крови на 7-е и 60-е сут - «эффект спасения», на 14-е и 30-е сут было зарегистрировано снижение этого показателя (рис. 7). У облучённых животных, которых содержали в одной клетке с перегородкой вместе с необлучёнными животными, было зарегистрировано повышение относительного количества лимфоцитов на 3-и, 14-е и 60-е сут («эффект спасения») и снижение показателя на 30-е сут (рис. 8).
—*—СВИДОБЛ
-В-БИОКОНТРОЛЬ
Рис. 5. Относительное количество лимфоцитов периферической крови у необлучённых мышей при содержании их в одной клетке без перегородки с облучёнными мышами (гамма-облучение) в течение 90 сут.
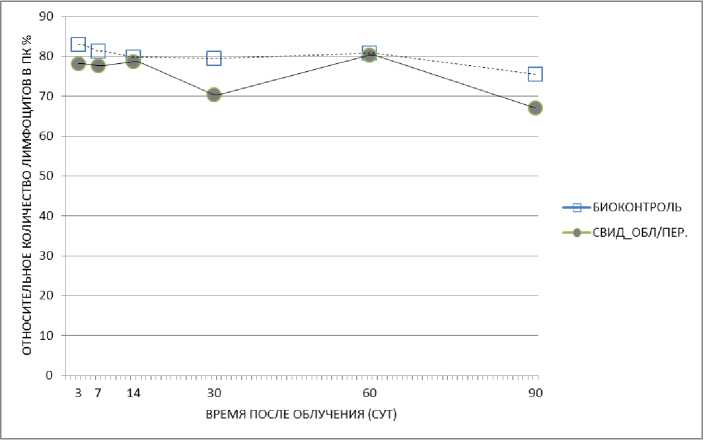
Рис. 6. Относительное количество лимфоцитов периферической крови у необлучённых мышей при содержании их в одной клетке с перегородкой с облучёнными мышами (гамма-облучение) в динамике.
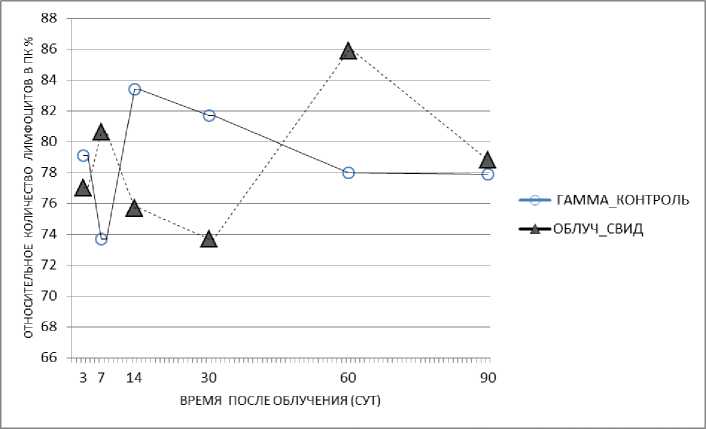
Рис. 7. Относительное количество лимфоцитов периферической крови у облучённых мышей (гамма-облучение) при содержании их в одной клетке без перегородки с необлучёнными мышами в течение 90 сут.
Таким образом, у облучённых животных, содержавшихся в клетке с перегородкой совместно с необлучёнными животными, зарегистрировано повышение относительного количества лимфоцитов, которое можно определить как «эффект спасения» в данном эксперименте.
Проведение многофакторного дисперсионного анализа показало, что фактор облучения снижает относительное количество лимфоцитов в зависимости от времени после начала эксперимента на 0,04% в день и «Фактор контакта» оказывал статистически значимое влияние на анализируемый показатель (F=6,07; p=0,0026) (табл. 2).
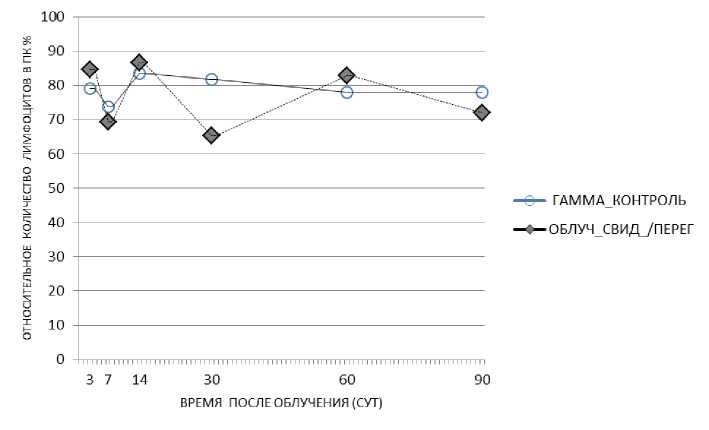
Рис. 8 . Относительное количество лимфоцитов периферической крови у облучённых мышей (гамма-облучение) при содержании их в одной клетке с перегородкой с необлучёнными мышами в течение 90 сут.
Таблица 2
Влияние различных факторов на количество лимфоцитов в периферической крови у мышей
|
Параметр |
F |
р |
|
Фактор контакта |
6,07 |
0,003 |
|
День |
8,54 |
0,004 |
|
Облучение Фактор контакта * |
0,82 |
0,440 |
|
Облучение |
0,23 |
0,640 |
Примечания: * R2=0,062.
Таблица 3
Параметры обобщённой линейной модели при анализе влияния различных факторов на количество лимфоцитов в периферической крови у мышей
|
Параметр |
B |
t |
р |
|
Свободный член |
80,73 |
101,20 |
2,3x10-255 |
|
[Фактор контакта=0] |
0,00 |
- |
- |
|
[Фактор контакта=1] |
-3,32 |
-3,45 |
6,3x10-4 |
|
[Фактор контакта=2] |
-1,28 |
-1,30 |
0,190 |
|
День |
-0,04 |
-2,92 |
0,004 |
При этом статистически значимые отличия были характерны при содержании облучённых и необлучённых мышей в одной клетке с перегородкой. Такие условия эксперимента вызывали у облучённых и необлучённых животных-«свидетелей» снижение доли лимфоцитов в периферической крови в среднем на 3,3% (табл. 3).
Обсуждение и заключение
В опубликованных данных [1, 3] было показано, что биологические эффекты в гемопоэтических клетках после действия ИИ могут быть обусловлены изменениями в геноме клеток в виде генных мутаций, изменения экспрессии генов, хромосомных аберраций, повреждений структуры
ДНК, её регенерации, изменений количества внутриклеточных активных форм кислорода, апоптоза и других явлений.
При анализе полученных в настоящем исследовании данных предполагается, что радиационно-индуцированные «эффекты свидетеля» могут быть обнаружены и на межорганизменном уровне, когда у необлучённых животных, находящихся рядом с облучёнными, также могут наблюдаться изменения, схожие с обнаруженными у облучённых животных. Для объяснения этого явления в публикациях предполагается несколько механизмов: 1) непосредственный контакт и передача повреждающих факторов; 2) выделение соединений, вызывающих изменения у необлу-чённых животных на расстоянии [13, 18]; 3) действие внеклеточной ДНК [19]; 4) действие биофотонов [8, 9] и др.
В данном исследовании при проведении многофакторного анализа было выявлено статистически значимое влияние фактора «День» и фактора «Облучение». При этом гамма-облучение приводит к снижению количества лейкоцитов в периферической крови мышей на 4,7 млн лейкоцитов в мл. Доля лимфоцитов в периферической крови мышей снижается в среднем на 3,3%. Показатель фактора времени и фактора облучения при изучении лимфоцитов также продемонстрировал снижение числа лимфоцитов.
У необлучённых животных, содержавшихся с облучёнными в одной клетке как с перегородкой, так и без перегородки, наблюдался «эффект свидетеля». У облучённых животных, содержащихся с необлучёнными в одной клетке без перегородки на 7-е и 60-е сут после облучения, и содержавшихся с необлучёнными животными в одной клетке с перегородкой на 3-и, 14-е, и 60-е сут, обнаружено повышение числа лимфоцитов по сравнению с гамма-контролем - «эффект спасения».
Мы предполагаем, что на уровне организма сочетание эффектов модификации ДНК с последующим апоптозом, другими клеточными явлениями и образованием летучих метаболитов, которые воздействуют на клетки крови индуцируют «эффект свидетеля», обратной стороной которого является «эффект спасения». Механизм происходящих процессов на уровне «клеток-свидетелей» и «клеток спасения» при межорганизменном контакте не ясен, но можно предположить, что те же вышеназванные биологические вещества, а также такие, как оксид азота, активные формы кислорода, цитокины и другие через каскад сигнальных процессов могут привести к подобным биологическим нарушениям.
Полученные результаты предполагают необходимость дальнейших исследований радиационно-индуцированных «эффекта свидетеля» и «эффекта спасения» с использованием молекулярно-клеточных методических подходов.
Работа выполнена в рамках государственного задания Минобрнауки России (Регистрационный номер № 122040400089-6 ) за счёт субсидии, выделенной ФИЦ ХФ РАН на выполнение государственного задания, тема: «Изучение принципов структурно-функциональной организации биомолекулярных систем, разработка методов дизайна их физико-химических аналогов и создание на этой основе биологически активных препаратов нового поколения».
Список литературы Исследование действия ионизирующего излучения на радиационно- индуцируемые изменения клеток системы крови мышей на уровне организма
- Lehnert B.E., Goodwin E.H., Deshpande A. Extracellular factor(s) following exposure to alpha particles can cause sister chromatid exchanges in normal human cells chromatid exchanges in normal human cells. Cancer Res., 1997, vol. 57, no. 11, pp. 2164-2171.
- Prise K.M., Belyakov O.V., Folkard M., Michael B.D. Studies of bystander effects in human fibroblasts using a charged particle microbeam. Int. J. Radiat. Biol., 1998, vol. 74, no. 6, pp. 793-798.
- Lyng F.M., Seymour C.B., Mothersill C. Initiation of apoptosis in cells exposed to medium from the progeny of irradiated cells: a possible mechanism for bystander-induced genomic instability? Radiat. Res., 2002, vol. 157, no. 4, pp. 365-370.
- Coates P.J., Lorimore S.A., Wright E.G. Damaging and protective cell signalling in the untargeted effects of ionizing radiation. Mutat. Res., 2004, vol. 568, no. 1, pp. 5-20.
- Matsumoto H., Takahashi A., Ohnishi T. Radiation-induced adaptive responses and bystander effects. Biol. Sci. Space, 2004. vol. 18, no. 4, pp. 247-254.
- Reis P., Lourenço J., Carvalho F.P., Oliveira J., Malta M., Mendo S., Pereira R. RIBE at an inter-organismic level: a study on genotoxic effects in Daphnia magna exposed to waterborne uranium and a uranium mine effluent. Aquat. Toxicol., 2018, vol. 198, pp. 206-214.
- Mothersill C., Smith R.W., Saroya R., Denbeigh J., Rowe B., Banevicius L., Timmins R., Moccia R., Seymour C.B. Irradiation of rainbow trout at early life stages results in legacy effects in adults. Int. J. Radiat. Biol., 2010, vol. 86, no. 10, pp. 817-828.
- Mothersill C., Smith R., Wang J., Rusin A., Fernandez-Palomo C., Fazzari J., Seymour C. Biological entanglement-like effect after communication of fish prior to X-ray exposure. Dose Response, 2018, vol. 16, no. 1, pp. 1559325817750067. DOI: 10.1177/1559325817750067.
- Smith R., Wang J., Seymour C., Fernandez-Palomo C., Fazzari J., Schültke E., Bräuer-Krisch E., Laissue J., Schroll C., Mothersill C. Homogenous and microbeam X-ray radiation induces proteomic changes in the brains of irradiated rats and in the brains of nonirradiated cage mate rats. Dose Response, 2018, vol. 16, no. 1, pp. 1559325817750068. DOI: 10.1177/1559325817750068.
- Petushkova V.V., Pelevina I.I., Kogarko I.N., Neyfakh E.А., Kogarko B.S., Ktitorova O.V. Radiation- induced inter-organism bystander effects. Some aspects of the transmission. Radiatsionnaya biologiya. Radioekologiya – Radiation Biology. Radioecology, 2020, vol. 60, no. 3, pp. 229-238. (In Russian).
- Petushkova V.V., Pelevina I.I., Kogarko I.N., Neyfakh E.А., Kogarko B.S., Ktitorova O.V. Radiation- induced inter-organism bystander effects. Some aspects of the transmission. Biology Bulletin, 2020, vol. 47, no. 12, pp. 1610-1617.
- Andreev S.S., Aldibekova A.E., Kogarko I.N., Petushkova V.V., Akleev A.B. New evidence of inter- organism bystander effects in mice. VIII Congress on Radiation Research Collected works, abstracts of reports. Dubna, JINR, 2021, pp. 83. (In Russian).
- Pelevina I.I., Akleev A.V., Kogarko I.N., Petushkova V.V., Kogarko B.S., Pryakhin E.A., Neyfakh Е.А., Ktitorova O.V. Radiation-chemical effects of ionizing radiation on the organism and genotoxic disorders of the blood system. Khimicheskaya fizika – Russian Journal of Physical Chemistry, 2021, vol. 40, no. 12, pp. 48-55. (In Russian).
- Pelevina I.I., Kogarko I.N., Pryakhin E.A., Petushkova V.V., Kogarko B.S., Neyfakh Е.А., Andreev S.S., Ktitorova O.V., Ganeev I.I. Study of the disorders of the blood system of irradiated and unirradiated animals kept in contact. Radiatsionnaya biologiya. Radioekologiya – Radiation Biology. Radioecology, 2022, vol. 62, no. 5, pp. 477-483. (In Russian).
- Kogarko I.N., Petushkova V.V., Kogarko B.S., Pryakhin E.A., V.V. Neyfakh Е.А., Ktitorova O.V., Andreev S.S., Ganeev I.I., Kuzmina N.S., Selivanova E.I. Investigation of the role of contact effects of irradiated animals on unirradiated animals under the influence of radiation. Annual XXVII conference of the FRCCP RAS of the department of CIMBIO: abstracts of reports. Moscow, 2022, pp. 77-86. (In Russian).
- Directive 2010/63/EU of the European Parliament and of the Council of the European Union on the protection of animals used for scientific purposes. St. Petersburg, Rus-LASA, 2012. (In Russian).
- European Convention for the protection of vertebrate animals used for experimental or other scientific pur-poses. Strasbourg, 1986. Available at: https://rm.coe.int/168007a6a8 (Accessed 05.04.2022). (In Russian).
- Surinov B.P., Isaeva V.G., Dukhova N.N., Sharetsky A.N. The changes in immunomodulatory and attractive properties of mice secretions after radiation exposure or induction of the “bystander effects”. Radiatsionnaya biologiya. Radioekologiya – Radiation Biology. Radioecology, 2021, vol. 61. no. 1, pp. 5-12. (In Russian).
- Ermakov A.V., Konkova M.S., Kostyuk S.V., Veiko N.A. DNA signaling pathway that ensures the development of the radiation effect of the witness in human cells. Radiatsionnaya biologiya. Radioekologiya – Radiation Biology. Radioecology, 2011, vol. 51, no. 6, pp. 651-659. (In Russian).
