Исследование динамики пространственных характеристик Θ-диапазона ЭЭГ у испытуемых с различной результативностью когнитивной деятельности

Автор: Каратыгин Н.А., Коробейникова И.И.
Журнал: Академический журнал Западной Сибири @ajws
Рубрика: Эксперимент
Статья в выпуске: 1 (72) т.14, 2018 года.
Бесплатный доступ
У испытуемых (42 человека, мужчины 20-21 года) с различной успешностью и скоростью выполнения зрительно-моторной задачи (компьютеризированный вариант теста Горбова-Шульте) исследовали пространственную организацию и динамику потенциалов 0-диапазона (4-7 Гц) ЭЭГ. У испытуемых 1-й группы, которые быстро (среднее время клика 6318±182,5) и с малым количеством ошибок (1,6±0,43) выполняли задание, во время работы (по отношению к состояниям спокойного бодрствования, вне работы) наблюдалось существенное изменение структуры и количества высококогерентных (коэффициент когерентности>0,6) взаимосвязей в диапазоне Ɵ-ритма ЭЭГ. У индивидов 2-й группы, выполнявших задание медленно (среднее время клика 9827±309,2) и с большим количеством ошибок (11,5±1,95), изменения структуры высококогерентных связей были менее выражены, а изменения их количества не достигали уровня достоверности.
Когнитивная деятельность, результативность, когерентность потенциалов, ɵ-диапазон ээг
Короткий адрес: https://sciup.org/140225912
IDR: 140225912
The study of the dynamics of spatial characteristics in the Θ- frequency band EEG in subjects with different performance of cognitive activity
In subjects (42 people, men 20-21 years old) with different performance and speed of execution of visual-motor tasks (computerized version of the Gorbov-Shultetest) the dynamics and spatial characteristics of potentials in the Ɵ-frequency band (4-7 Hz) EEG was registered. Subjects of the 1st group who quickly (the average time of click 6318±182,5) and with a small number of errors (1,6±0,43) performed the task, during the work (in relation to the states of quiet wakefulness, out of work) there was a significant change in the structure and the number of highly coherent (coherence coefficient>0,6) interconnections in the Ɵ-range of EEG. In individuals of the 2nd group who performed the task slowly (the average time of click 9827±309,2) and with a large number of errors (11,5±1,95), the changes in the structure of highly coherent bonds were significantly less pronounced, and the change in their number did not reach the level of reliability.
Текст научной статьи Исследование динамики пространственных характеристик Θ-диапазона ЭЭГ у испытуемых с различной результативностью когнитивной деятельности
Одним из методов, которые в настоящее время широко применяются при анализе ЭЭГ, является когерентный анализ [10]. Синхронизация биопотенциалов ЭЭГ различных зон коры больших полушарий мозга дает богатую информацию о функциональном состоянии человека и о включении тех или иных зон коры в единую систему обеспечивающую протекание высших психических функций человека [9].
В наших предыдущих исследованиях было продемонстрировано, что картина распределения синхронизации биопотенциалов ЭЭГ в различных корковых зонах отражает скоростные характеристики и успешность выполнения зрительно - моторных заданий [2, 5], что хорошо согласуется с результатами целого ряда работ, указывающих на связь показателей когерентности со скоростью [23] и с успешностью выполнения различных задач [14].
К настоящему времени накоплен большой объем данных о связи 9-активности с процессами памяти и произвольного внимания, которые безусловно являются одним из ключевых компонентов, влияющих на успешность выполнения зрительномоторных заданий [8, 24]. Принимая во внимание вышеизложенное, в данном исследовании был сделан акцент на анализ картины распределения и динамики когерентных взаимосвязей 9-диапазона ЭЭГ.
Целью настоящего исследования явился поиск функциональной связи пространственных характеристик 9-диапазона ЭЭГ с результативностью выполнения зрительно-моторного задания.
Методика исследования.
В исследовании на основе добровольного информированного согласия участвовали 42 испытуемых (мужчины в возрасте 18-21 года). Процедура обследования соответствовала стандартам локального этического комитета Первого ММГУ им. И.М. Сеченова. В качестве зрительно-моторной задачи использовался компьютеризированный тест Горбова-Шульте (красно-черные таблицы). Во время обследования испытуемый находился в удобном кресле перед экраном монитора (17 дюймов). На мониторе отображалась таблица (7х7 квадратов), содержащая 24 красных и 25 черных квадратов, на каждом из которых имелся собственный номер. Обследование включало 2 серии, каждая из которых состояла из двух заданий. Паттерны расположения квадратов для каждого задания были изначально заданы методом случайной генерации. Для всех испытуемых использовался одинаковый набор паттернов. В первой серии обследования, на основе предварительной инструкции испытуемый должен был в первом задании (КЧ) выбрать (указать курсором мышки) черные квадраты в порядке возрастания их номеров от (1 до 25), а затем - красные квадраты в порядке убывания номеров (от 24 до 1). Во втором задании (ЧЕРЕД) испытуемый должен был выбрать по очереди черные квадраты в порядке возрастания, а красные в порядке убывания: 1 черный, 24 красный, 2 черный, 23 красный и т.д. Во второй серии обследования испытуемому предлагали выполнить те же задания в сопровождении голосовой помехи: чтение диктором цифр от 1 до 25 в случайном порядке, не совпадающим с порядком указания испытуемым номеров квадратов и с частотой одна цифра в две секунды.
По результатам выполнения компьютерной задачи для каждого испытуемого вычисляли следующие показатели:
-
- время между последовательными кликами по квадратам, величина которого усреднялась по каждому заданию (среднее время клика, мс);
-
- число ошибок в каждом задании - включая ошибки последовательности и неверное указание цвета квадрата. Если испытуемый сбивался и отказывался от дальнейшего выполнения задания, количество оставшихся неотмеченными квадратов расценивалось как ошибки.
ЭЭГ регистрировали в исходном состоянии при открытых (Ф1ОГ) и закрытых (Ф1ЗГ) глазах, во время выполнения заданий первой серии обследования (КЧ, ЧЕРЕД), в состоянии спокойного бодрствования в перерыве между первой и второй сериями (Ф2ОГ), во время выполнения заданий второй серии (КЧ голос, ЧЕРЕД голос) и в состоянии спокойного бодрствования после выполнения заданий (Ф3ОГ). ЭЭГ регистрировали с помощью электроэнцефалографа «Нейрон-спектр» (г. Иваново) монополярно по схеме “10–20” в затылочных (О2, О1), теменных (Р4, Р3), центральных (С4, С3), лобных (F4, F3) и височных (Т4, Т3) отведениях. Объединённые референтные электроды располагались на мочках ушей. Полоса фильтрации составляла 0,5-35,0 Гц, постоянная времени - 0,32 с, режек-тивный фильтр - 50 Гц. После регистрации все записи ЭЭГ были переведены в компьютерную систему анализа и топографического картирования электрической активности мозга «BRAINSYS» для Windows и обработаны с помощью аппаратно - программного комплекса «НЕЙРО-КМ» (ООО «Стато-кин», г. Москва). Артефакты исключали из анализируемой записи с использованием возможностей программного комплекса BRAINSYS. Спектральнокогерентный анализ ЭЭГ проводили на основе быстрого преобразования Фурье (пакет программ BRAINSYS). Эпоха анализа составляла 4 сек при длительности каждого фрагмента в 1 мин, частота оцифровки - 200 Гц. Для всех 45 пар отведений вычисляли средние значения функции когерентности (Кког) в 9-диапазоне (4-7 Гц). При анализе учитывались только связи Кког которых превышал 0,6.
Для статистической обработки использовался программный пакет STATISTICA v.8.0. При нормальном распределении анализируемых показате- лей вычисляли среднее значение (M) и стандартную ошибку среднего (m).
Достоверность различий анализируемых показателей у испытуемых выделенных групп оценивали с помощью t-критерия для независимых выборок, а достоверность изменения значений показателей в разных заданиях у одной группы испытуемых оценивали с использованием t-критерия для связанных выборок.
Результаты исследования.
По соотношению скорости и успешности выполнения задания было выделено две группы испытуемых. В качестве критерия выделения групп использовали ( М±т ), при этом в качестве показателя скорости выполнения задания использовали сумму средних времен клика на этапах ЧЕРЕД и ЧЕРЕД голос, а в качестве показателя успешности использовали сумму ошибок по ЧЕРЕД и ЧЕРЕД голос. По группе испытуемых в целом сумма средних времен клика на этапах ЧЕРЕД и ЧЕРЕД голос составила 7961,2±241,3, а сумма ошибок по ЧЕРЕД и ЧЕРЕД голос - 5,81±0,758. В 1-ю группу ( n=10 ) вошли лица быстро (среднее время клика 6318±182,5) и успешно (количество ошибок составило 1,6±0,43) выполнившие задания, во 2-ю группу ( n=8 ) - испытуемые выполнившие задания медленно (среднее время клика 9827±309,2) и с большим количеством ошибок (11,5±1,95).
У испытуемых 1-й группы в состоянии спокойного бодрствования как при открытых (Ф1ОГ), так и при закрытых (Ф1ЗГ) глазах в 9-диапазоне ЭЭГ наблюдался комплекс внутриполушарных когерентных взаимосвязей между соседними областями коры левого (О1-Р3, Р3-С3, C3-F3) и правого (О2-Р4, Р4-С4, C4-F4) полушарий и между отведениями P4-F4 правого полушария (рис. 1-А1). Кроме этого в исходном состоянии (Ф1ОГ, Ф1ЗГ) имела место межполушарная когерентность потенциалов в отведениях F3-F4, С3-С4 , а при закрытых глазах в отведениях C3-F4 , C4-F3 . Паттерн когерентных взаимосвязей на этапах Ф2ОГ и Ф3ОГ был практически идентичен наблюдавшемуся в состоянии Ф1ОГ. Во всех ситуациях спокойного бодрствования при открытых глазах наблюдалась когерентность между затылочными ( О2 ) и центральными (С4) зонами правого полушария.
У испытуемых 1-й группы во время выполнения заданий рисунок когерентных взаимосвязей существенно изменялся (рис. 1-Б1). Все когерентные взаимосвязи наблюдавшиеся в исходном состоянии и на этапах спокойного бодрствования между и после заданий сохранялись и при работе (за исключением когерентности между О2 и С4 ). В дополнение к этому во время выполнения заданий появлялся новый набор когерентных взаимосвязей. Увеличивалось количество межцентральных связей, Кког О1-О2 (кроме этапа ЧЕРЕД голос) и Р3-Р4 превысили 0,6.
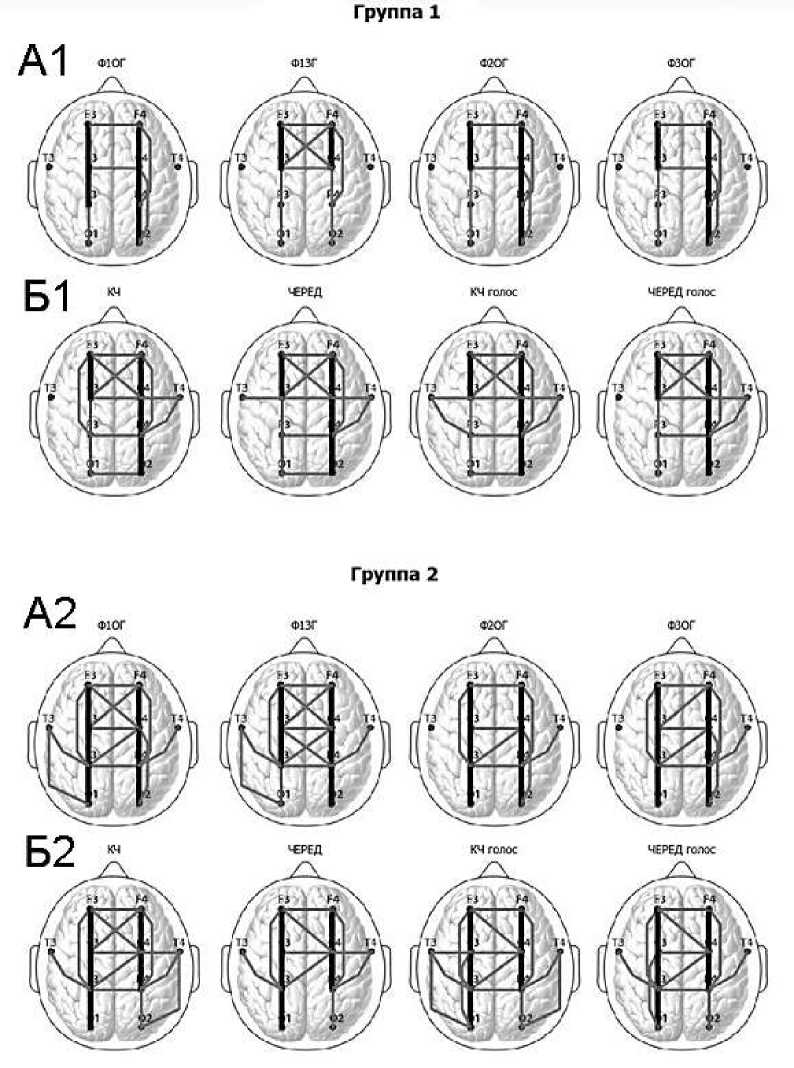
Рис. 1. Топографическое распределение когерентных связей 9-диапазона ЭЭГ испытуемых 1-й и 2-й групп на этапах обследования: А1- этапы фона и спокойного бодрствования у испытуемых 1-й группы; Б1- этапы выполнения задания у испытуемых 1-й группы; А2- этапы фона и спокойного бодрствования у испытуемых 2-й группы; Б2- этапы выполнения задания у испытуемых 2-й группы. Тонкие серые линии — 0,6< Кког <0,8, толстые черные линии — Кког >0,8.
Наряду с этим у испытуемых 1-й группы во время выполнения всех заданий выделилась система когерентных связей с фокусом в правой височной зоне ( Т4-Р4 и Т4-С4 ). На этапах КЧ голос, ЧЕРЕД появлялись когерентные взаимосвязи в отведениях Т3-С3 ; на этапе КЧ голос - в отведениях Т3-Р3 ; на всех этапах выполнения заданий - в отведениях C3-F4 и C4-F3.
У испытуемых 2-й группы (рис. 1-А2) на этапах Ф1ЗГ и Ф1ОГ имело место высокое взаимосодей-ствие биопотенциалов 9-диапазона (Кког>0,8) соседних корковых зон левого (Р3-С3, C3-F3) и правого (О2-Р4, Р4-С4, C4-F4) полушарий. Наблюдалась межполушарная когерентность между гомологичными (F3-F4, С3-С4, Р3-Р4) и негомологичными (C3-F4, C4-F3, Р3-С4; Р4-С3 в случае Ф1ЗГ) отве- дениями. Присутствовал ряд внутриполушарных длиннодистантных когерентных взаимосвязей (Р3-F3; О2-С4, Р4-F4 в случае Ф1ОГ). Особое внимание обращает на себя факт наличия в фоне когерентности потенциалов ЭЭГ в отведениях Т3-Р3, Т4-Р4, и Т3-О1.
На этапах Ф2ОГ и Ф3ОГ картина когерентных связей в целом оставалась идентичной фоновой (Ф1ОГ), за исключением уменьшения когерентности (Кког ниже 0,6) комплекса связей левой височной зоны ( Т3-Р3, Т3-О1 ) и у фронтальноцентральных межполушарных связей ( C3-F4, C4-F3 в случае Ф2ОГ; C4-F3 в случае Ф3ОГ).
Группа 1

Ф1ОГ Ф1ЗГ
КЧ
Ф1ОГ Ф1ЗГ
КЧ
........
ЧЕРЕД Ф2ОГ КЧ гол ЧЕРЕД Ф3ОГ гол
Этапы обследования
Группа 2

ilium
ЧЕРЕД Ф2ОГ КЧ гол ЧЕРЕД Ф3ОГ гол
Этапы обследования
Рис. 2. Количество когерентных связей (Кког>0,6) 9-диапазона ЭЭГ у испытуемых 1-й и 2-й групп на этапах обследования: синие столбики - этапы фона и спокойного бодрствования, красные - выполнение заданий. Звездочками отмечены достоверные различия между количеством когерентных связей на этапах обследования. Достоверность различий приведена в тексте.
У испытуемых 2-й группы во время выполнения заданий (Рис. 1-Б2) картина когерентных взаимосвязей в целом оставалась похожей на таковую в исходном состоянии. Сохранялась внутриполушар-ная когерентность потенциалов между соседними корковыми зонами левого (О1-Р3, Р3-С3, C3-F3) и правого (О2-Р4, Р4-С4, C4-F4) полушарий, межпо- лушарная когерентность между гомологичными (F3-F4, С3-С4, Р3-Р4 кроме этапа ЧЕРЕД) и негомологичными (C4-F3, Р3-С4; C3-F4 в случае КЧ) областями. На всех этапах работы также сохранялись, присутствовавшие в фоне при открытых глазах, длиннодистантные когерентные связи между теменными и фронтальными зонами правого и левого полушарий (Р3-F3 и Р4-F4). При выполнении заданий 2-й серии, когда испытуемым приходилось работать в условиях голосовой помехи, в левом полушарии усиливалась синхронизация затылочных и центральных областей (О1-С3).
У испытуемых 2-й группы, как и у испытуемых 1-й группы, во время выполнения почти всех заданий (кроме ЧЕРЕД голос) наблюдалось увеличение активности системы правополушарных височных связей ( Т4-Р4 сохранялась во всех ситуациях; Т4-С4 достигала уровня 0,6 при выполнении КЧ и ЧЕРЕД; Т4-О2 при КЧ и КЧ голос). Также на всех этапах работы сохранялась, присутствовавшая в фоне (Ф1ЗГ и Ф1ОГ) когерентность в отведениях Т3-Р3 , кроме этого на этапе КЧ голос когерентность в отведениях С3-Т3 и О1-Т3 достигала уровня 0,6.
При последующей обработке для каждого испытуемого просчитывалось количество когерентных связей (Кког>0,6) на каждом этапе обследования, после чего определялось среднее количество таких связей для испытуемых 1-й и 2-й групп.
Было выявлено, что у испытуемых 1-й группы при выполнении всех заданий количество когерентных взаимосвязей было достоверно выше относительно предшествующих или последующих этапов спокойного бодрствования (Ф1ЗГ-КЧ р =0,0021; ЧЕРЕД-Ф2ОГ р =0,0065; Ф2ОГ-КЧ голос р =0,0013; ЧЕРЕД голос-ФзОГ р =0,0251) (рис. 2). У испытуемых 2-й группы, изменение количества когерентных связей было менее выражено и не достигало достоверного уровня значимости.
Обсуждение результатов.
На основании полученных данных у испытуемых 1-й группы можно выделить три основных феномена, отражающих изменение картины синхронизации 9-активности разных зон коры при работе по отношению к этапам спокойного бодрствования: усиление когерентных взаимосвязей между негомологичными передне-центральными областями; увеличение активности системы когерентных связей с фокусом в правой височной зоне; усиление межполушарных взаимосвязей между теменными, а также между затылочными областями. Усиление когерентности между негомологичными передне - центральными областями (C3-F4 и C4-F3), наблюдавшееся при выполнении всех заданий испытуемыми 1-й группы, может свидетельствовать о более полном включении в синхронизацию областей префронтальной коры. Связь 9-активности префронтальной коры с памятью и процессами произвольного внимания продемонстрирована во многих ра- ботах [3, 8, 17]. Косвенным подтверждением важности θ-синхронизации в префронтальной и медиа-фронтальной коре является изменение исполнительных функций человека при внешнем синхронизирующем или десинхронизирующем влиянии на фоне транскраниальной электрической (tACS) или магнитной (TBS) стимуляции на частотах θ-ритма [7, 19].
Усиление θ-синхронизации во время работы у испытуемых 1-й группы в системе когерентных связей с фокусом в правой височной зоне неудивительно, так как во многих исследованиях было показано, что именно правое полушарие отвечает за наглядно-образное мышление, симультанную обработку зрительной информации и оперирование целостными образами [6, 13]. Особенно ярко это было продемонстрировано на пациентах с расщепленным мозгом [15, 16, 20]. Также установлена связь изменения когерентности в правом полушарии с успешностью выполнения эвристических задач [4].
Выявленное в настоящем исследовании усиление межполушарных взаимосвязей между теменными и между затылочными областями во время работы у испытуемых 1-й группы согласуется с результатами исследования по выявлению электрофизиологических коррелятов разной когнитивной нагрузки, где было показано увеличение θ-активности в затылочных, теменных и центральных корковых зонах при увеличении сложности задания [11].
Как уже отмечалось выше, увеличение когерентности в θ-диапазоне наблюдалось при выполнении заданий, требующих участия рабочей памяти [12, 21]. Наряду с этим имеется ряд работ, в которых рассматривалась связь θ-активности с поддержанием произвольного внимания [3, 22]. Возникает вопрос: почему несмотря на то, что во всех ситуациях фона и спокойного бодрствования у испытуемых 2-й группы было больше когерентных связей θ-диапазона, а на этапах выполнения заданий количество связей было сравнимо с количеством связей у испытуемых 1-й группы, они допускали большее количество ошибок и работали с меньшей скоростью, чем испытуемые 1-й группы? Логично предположить, что θ-активность связана с успешностью и скоростью выполнения зрительно-моторных задач опосредованно, через задействование процессов рабочей памяти и произвольного внимания. Так как зрительно-моторная задача, которую мы использовали в исследовании, требует не только запоминания, удержания и воспроизведения информации, но и построения определенной стратегии решения задачи, синхронизация на частоте θ-ритма может напрямую не отражать успешность выполнения задания. Изначально большее количество когерентных связей в фоновых состояниях и невозможность значительного увеличения их количества при работе может быть интерпретировано как снижение адаптационных возможностей у испытуемых 2-й группы [1], что в результате проявилось в более низкой скорости и меньшей успешности выполнения задания.
Таким образом, высокая синхронизация потенциалов θ-диапазона может являться коррелятом нормального функционирования систем памяти и произвольного внимания, и, наряду с этим, некоторого снижения адаптационных возможностей человека, если такая синхронизация наблюдается в фоне, но не меняется при работе. В этих условиях анализ именно динамики когерентности биопотенциалов θ-диапазона представляется наиболее эффективным инструментом, позволяющим сопоставить параметры электрической активности мозга с результативностью когнитивной деятельности человека.
Выводы.
-
1. У испытуемых, которые выполняли задание быстро и с малым количеством ошибок, наблюдалась выраженная динамика числа высоких когерентных взаимосвязей θ-диапазона ЭЭГ. Число когерентных связей в фоне и в состоянии спокойного бодрствования было достоверно ниже, чем при выполнении задания. При выполнении заданий относительно фоновых состояний наблюдалось: усиление когерентных взаимосвязей между негомологичными передне-центральными областями; увеличение активности системы когерентных связей с фокусом в правой височной зоне; усиление межполушарных взаимосвязей между теменными и между затылочными областями коры.
-
2. Для испытуемых, выполнявших задание медленно и с большим количеством ошибок, выраженные изменения числа высоких когерентных взаимосвязей биапотенциалов θ-диапазона ЭЭГ при выполнении заданий относительно фоновых состояний не выявлены.
Список литературы Исследование динамики пространственных характеристик Θ-диапазона ЭЭГ у испытуемых с различной результативностью когнитивной деятельности
- Баевский Р.М. Проблема оценки и прогнозирования функционального состояния организма и ее развитие в космической медицине//Успехи физиологических наук. -2006. -Т. 37, № 3. -С. 42-57.
- Джебраилова Т.Д., Коробейникова И.И., Каратыгин Н.А. Лабильность структуры когерентных взаимосвязей биопотенциалов в диапазонах основных ритмов ЭЭГ и эффективность интеллектуальной деятельности человека//Академический журнал Западной Сибири. -2014. -Т. 10, № 3 (52). -С. 58-60.
- Мачинская Р.И., Курганский А.В. Сравнительное электрофизиологическое исследование регуляторных компонентов рабочей памяти у взрослых и детей 7-8 лет. Анализ когерентности ритмов ЭЭГ//Физиология человека. -2012. -Т. 38, № 1. -С. 5-19.
- Разумникова О.М. Частотно-пространственная организация активности коры мозга при конвергентном и дивергентном мышлении в зависимости от фактора пола. Сообщение П. Анализ когерентности ЭЭГ//Физиология человека. -2005. -Т. 31, № 3. -С. 39-49.
- Судаков К.В., Джебраилова Т.Д., Коробейникова И.И., Каратыгин Н.А. Геометрические образы (обзоры) когерентных взаимоотношений биопотенциалов различных частотных диапазонов ЭЭГ в динамике целенаправленной деятельности человека//Российский физиологический журнал. -2013. -Т. 99, В. 6. -С. 706-718.
- Хрестоматия по нейропсихологии. под редакцией Хомской Е.Д. М.: Институт общегуманитарных исследований Московский психолого-социальный институт, 2004.
- Alekseichuk I., Pabel S.C., Antal A., Paulus W. Intrahemispheric theta rhythm desynchronization impairs working memory//Restorative neurology and neuroscience. -2017. -V. 35, № 2. -P. 147-158.
- Backus A.R., Schoffelen J.M., Szebenyi S., Hanslmayr S., Doeller C.F. Hippocampal-prefrontal theta oscillations support memory integration//Current Biology. -2016. -V. 26, № 4. -P. 450-457.
- Bressler S.L. and Kelso J.A. Coordination dynamics in cognitive neuroscience.//Frontiers in neuroscience. -2016. -V. 10. -P. 397.
- Fingelkurts A., Fingelkurts A., Kahkonen S. Functional connectivity in the brain-is it an elusive concept?//Neurosci. Biobehav. Rev. -2005. -V. 28, № 8. -P. 827.
- Fournier L.R., Wilson G.F., Swain C.R. Electrophysiological, behavioral, and subjective indexes of workload when performing multiple tasks: manipulations of task difficulty and training//Int. J. of Psychophysiology. -1999. -V. 31, № 2. -P. 129-145.
- Hanslmayr S., Pastotter B., Bauml K.H., Gruber S., Wimber M., Klimesch W. The electrophysiological dynamics of interference during the Stroop task//J. of Cognitive Neuroscience. -2008. -V. 20, № 2. -P. 215-225.
- Herve P.Y., Zago L., Petit L., Mazoyer B., Tzourio-Mazoyer N. Revisiting human hemispheric specialization with neuroimaging//Trends in cognitive sciences. -2013. -V. 17, № 2. -P. 69-80.
- Hummel F., Gerloff H. Larger interregional synchrony is associated with greater behavioral success in a complex sensory integration task in humans//Cerebral Cortex. -2005. -V. 15, № 5. -P. 670-678.
- Kingstone A. Covert orienting in the split brain: Right hemisphere specialization for object-based attention//Laterality: Asymmetries of Body, Brain and Cognition. -2016. -V. 21, № 4-6. -P. 732-744.
- LeDoux J.E., Wilson D.H., Gazzaniga M.S. Manipulo-spatial aspects of cerebral lateralization: Clues to the origin of lateralization//Neuropsychologia. -1977. -V. 15, № 6. -P. 743-750.
- Lesting J., Daldrup T., Narayanan V., Himpe C., Seidenbecher T., Pape H.C. Directional theta coherence in prefrontal cortical to amygdalo-hippocampal pathways signals fear extinction//PloS one. -2013. -V. 8, № 10:e77707.
- Pinto N., Duarte M., Gonjalves H., Silva R., Gama J., Pato M.V. Bilateral theta-burst magnetic stimulation influence on event-related brain potentials//PloS one. -2018. -V. 13, № 1.:e0190693.
- Reinhart R.M. Disruption and rescue of interareal theta phase coupling and adaptive behavior//Proceedings of the National Academy of Sciences. 2017.:10257.
- Roser M.E., Fiser J., Aslin R.N., Gazzaniga M.S. Right hemisphere dominance in visual statistical learning//J. of cognitive neuroscience. -2011. -V. 23, № 5. -P. 1088-1099.
- Roux F., Uhlhaas P.J. Working memory and neural oscillations: alpha-gamma versus theta-gamma codes for distinct WM information?//Trends in cognitive sciences. -2014. -V. 18, № 1. -P. 16-25.
- Sauseng P., Hoppe J., Klimesch W. et al. Dissociation of sustained attention from central executive functions: local activity and interregional connectivity in the theta range//Eur. J. Neurosci. -2007. -V. 25, № 2. -P. 587.
- Silberstein R.B., Song J, Nunez P.L., Park W. Dynamic sculpting of brain connectivity is correlated with performance//Brain Topogr. -2004. -V. 16, № 4. -P. 249 -254.
- Thézé R., Guggisberg A.G., Nahum L., Schnider A. Rapid memory stabilization by transient theta coherence in the human medial temporal lobe//Hippocampus. -2016. -V. 26, № 4. -P. 445-454.
