Исследование электрических сигналов зерен пшеницы

Автор: Барышева Надежда Николаевна, Пронин Сергей Петрович
Журнал: Вестник аграрной науки Дона @don-agrarian-science
Рубрика: Технологии, средства механизации и энергетическое оборудование
Статья в выпуске: 2 (46), 2019 года.
Бесплатный доступ
Генерация электрических сигналов играет важную роль в жизни растений. Практическое применение исследований генерации электрических сигналов у высших растений носит широкий характер, одна из основных функций - это оценка возможности адаптации к изменениям условий внешней среды, оценка физиологического состояния. Основываясь на экспериментальные исследования, установлено, что электрические потенциалы зерен пшеницы могут быть использованы в качестве показателей их качества. Установлено, что при проращивании семян вдистиллированной водеэлектрический потенциал покоя равен -64 мВ для семян с высокой всхожестью, для низкой - составил -187 мВ. Опираясь на уравнение Гольдмана-Ходжкина-Катца, где значение потенциала характеризуется проницаемостью мембраны и концентрацией ионов, была выдвинута гипотеза о проницаемости оболочки семян пшеницы разной всхожести, которая подтвердилась в результате экспериментальных исследований и моделирования данных в среде MathCad. Чем выше всхожесть семян, тем выше проницаемость оболочки, а, следовательно, и ниже значение потенциала...
Зерна пшеницы, электрический потенциал, экспериментальное исследование, гипотеза, теоретическое исследование, раствор kcl, электрохимический процесс, проницаемость оболочки, экспресс-диагностика, всхожесть
Короткий адрес: https://sciup.org/140243663
IDR: 140243663 | УДК: 58.084.1
Текст научной статьи Исследование электрических сигналов зерен пшеницы
Введение. Генерация электрических сигналов играет важную роль в жизни растений. Генерация электрических сигналов характеризует физиологическое состояние растений [1]. Поэтому анализ природы и закономерностей электрических потенциалов у растений представляется весьма актуальным.
Впервые электрический сигнал был зафиксирован в нервных и мышечных клетках животных [2], далее были проведены исследования на высших растениях, которые подтвердили генерацию электрического импульса у высших и низших растений [3, 5].
Генерация электрических потенциалов у высших растений характеризуется и рядом специфических особенностей [3–5]. На сегодняшний день результаты исследований подтверждают тот факт, что процесс генерации биопотенциалов у животных и у растений не только сопровождает те или иные жизненно важные процессы, но и представляет основную роль в их протекании [6, 7].
Причина, по которой в растениях происходит генерация электрических сигналов, скорее всего, является необходимостью быстро реагировать на внешние раздражители, например, факторы стресса окружающей среды, в частности, свет, холод, ожог, давление, прикосновение, ранение и др. Было установлено, что различные экологические стимулы вызывают специфические реакции в живых клетках, которые способны передавать электрический сигнал в область воздействия [11, 14, 15]. В отличие от химических сигналов, электрические сигналы способны быстро передавать информацию на большие расстояния, поскольку за последние два десятилетия было обнаружено множество физиологических эффектов электрической сигнализации [11–13].
Практическое применение исследований генерации электрических сигналов у высших растений носит широкий характер. Одна из основных функций – это оценка возможности адаптации к изменениям условий внешней среды и оценка физиологического состояния.
Исследования генерации электрических сигналов семян пшеницы показали, что результаты могут быть использованы для диагностики качества посевного материала [17]. Например, значения электрического потенциала для зерен сорта «Алтайский янтарь» (мягкий сорт) с высокой всхожестью 97% составили -64 мВ, для семян с низкой всхожестью 87% составили -187 мВ. Чем ниже всхожесть семян, тем выше значение потенциала [18]. Установлено, что значение потенциала определяется электрохимическими процессами внутри зерен и проницаемостью их оболочек (мембран).
Понятие о том, что растительные мембраны проявляют проницаемость для конкретных ионов, было разработано в начале XX века Остерхаутом и другими учеными [18]. Эти идеи вызвали интенсивные исследования катионных проводимостей в растительных мембранах, что привело к обнаружению и детальной характеристике данной области [18–20].
Как правило, основной поток ионов представляет собой перемещение ионов Na+ , K+ и сг [7]. Поскольку диффузионные потенциалы, основанные на пассивном потоке ионов, зачастую хорошо согласуются с экспериментально измеряемой величиной потенциала [7, 8], для описания мембранного потенциала может быть использовано уравнение Гольдмана– Ходжкина–Катца.
Теоретические исследования показали, что мембраны растений более проницаемы для K+ [21] и за исключением в эпидермисе листьев гороха [22], K+ обычно является наиболее проницаемым ионом. В большинстве случаев K+ доминируют в пассивном притоке в корневые клетки [22]. Нарушение физиоло- гических свойств влечет за собой значительное уменьшение поглощения K+.
Использование солевых растворов показало резкое увеличение оттока ионов K+ при высокой солености, что подтверждено исследованиями мембраны хлопковых корневых волосков [23]. С увеличением концентрации NaCl наблюдалось смещение ионов Na+. В результате исследований появилось предположение, что Ca2+ защищает мембраны от неблагоприятного воздействия Na+, тем самым обеспечивает целостность мембраны и минимизирует утечку K+[23].
Исследование воздействия солевых растворов на изменение концентрации ионов зерен пшеницы вследствие генерации электрических сигналов является целью данной работы. Для решения поставленной задачи было проведено экспериментальное исследование.
Методика исследования. Исследование заключается в экспериментальном определении электрических потенциалов семян пшеницы и оценке их коррелируемости со всхожестью исследуемых семян.
Для измерения электрических потенциалов в растениях используются два разных метода, а именно: внеклеточной и внутриклеточной записи [1]. Наиболее популярными методами в настоящее время являются методы внутриклеточной записи. Однако при измерении электрических потенциалов зерен пшеницы многие современные методы исследования, а именно методы с использованием микроэлектродной техники, методы фиксации напряжения, пэтч-кламп-методы зачастую трудно применимы.
Для экспериментального исследования электрических сигналов зерен пшеницы были разработаны регистрирующие электроды. Электроды сделаны из стали. Один электрод выполнен в качестве зажима с углублением для более плотного контакта с поверхностью зерна, второй электрод представляет собой тонкую иглу длиной 4 мм [16].
Исследования выполнены с использованием пшеницы «Алтайский янтарь». Семена были предоставлены Россельхозцентром Алтайского края, всхожесть определена по ГОСТ 12038-84.
В результате исследований экспериментально установлено, что для зерен с высокой всхожестью 97% значение потенциала покоя составило -64 мВ, для семян со всхожестью 95% -76 мВ, со всхожестью 92% -105 мВ, 90% -125 мВ, 87% -187 мВ. Опираясь на уравнение Гольдмана–Ходжкина–Катца, такая разница значений может быть объяснена различной проницаемостью оболочек и концентрацией ионов Na+, K+ и Cl– у зерен с разной всхожестью.
Поскольку на этапе проращивания семян использовалась дистиллированная вода, теоретические значения потенциалов по уравнению Гольдмана– Ходжкина–Катца противоречили экспериментальным данным [18]. В дистиллированной воде внешняя концентрация ионов равна нолю. Внешние концентрации ионов образуются за счет внутренних ионов и зависят от проницаемости оболочки, поэтому в уравнение были внесены поправки:
RT , PK[K]in+(l-PK)[K]jout+PNa[Na]in+(l-PNa)[Na]out+(l-Pci)[Cl]in+Pci[cl]iout
F (l-PK)[K]in+PK[K]jout+(1-PNa)[Nahn+PNa[Na]out+Pci[ci]in+(1_Pci)[cl]jciut’ где R – универсальная газовая постоянная; T – абсолютная температура; F – постоянная Фарадея; Pk, PNa, PCl – коэффициенты проницаемости для ионов K+, Na+, Cl–; [K+]out, [Na+]out, [Cl–]out – внешние концентрации ионов K+, Na+, Cl–; [K+]in, [Na+]in, [Cl–]in – внутренние концентрации ионов K+, Na+, Cl–; [K+]jout, [Na+]jout, [Cl–]jout – концентрации ионов во внешних солевых растворах, используемых при проращивании зерен пшеницы, j – номер солевого раствора с заданной концентрацией.
В процессе исследования воздействия солевых растворов на изменение электрических потенциалов семян использовали растворы KCl, NaCl.
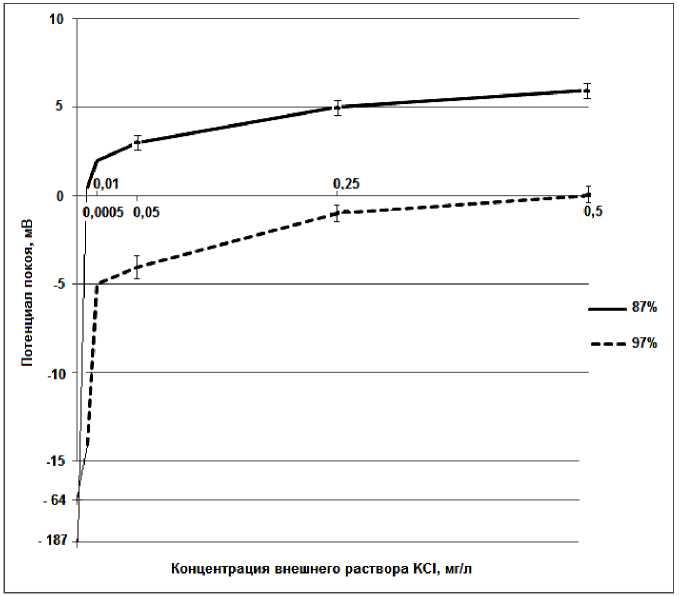
Рисунок 1 – Результаты экспериментальных исследований зерен пшеницы с разной всхожестью с использованием солевых растворов KCl c различной концентрацией
Методика исследования представляла собой проращивание семян в термокамере с использованием солевого раствора при температуре 20 °С в течение 12 часов, затем измерение электрических сигналов с помощью разработанных электродов [17].
Результаты исследований и их обсуждение. В результате исследований было выявлено, что использование солевых растворов существенно влияет на генерацию сигналов. Для анализа изменений сигналов задавали массовые концентрации растворов и измеряли потенциал покоя (ПП).
На рисунке 1 представлены графики экспериментальных исследований семян пшеницы с разной всхожестью при их замачивании в солевых растворах KCl с различными концентрациями.
Для сравнения приведены значения ПП по оси ординат при нулевой концентрации KCl. С целью уменьшения линейного размера оси ординат после значения -15 мВ она выполнена не в масштабе, но с указанием значений ПП. Поскольку в диапазоне концентраций KCl от 0 мг/л до 0,0005 мг/л измерения ПП не проводили, то экспериментальные точки ПП соеди- нены тонкими прямыми линиями. Вполне вероятно, что, согласно представленному уравнению, закон изменения ПП носит логарифмический характер.
При использовании раствора KCl потенциал покоя меняется не однозначно для зерен с разной всхожестью. При массовой концентрации ионов калия 0,0005 мг/л ПП для зерен с высокой всхожестью уменьшается с -64 мВ до -14 мВ, а для зерен с низкой всхожестью ПП падает до 0 мВ. Дальнейшее увеличение массовой концентрации ионов калия до 0,5 мг/л приводит к тому, что ПП для зерен с низкой всхожестью меняет знак на положительный и достигает +7 мВ, а ПП для зерен с высокой всхожестью монотонно уменьшается и приобретает значение 0 мВ.
Исходя из приведенного уравнения, подобный результат имеет место, если проницаемость оболочки для ионов калия и общая концентрация ионов у зерен с высокой всхожестью гораздо выше проницаемости оболочки и концентрации ионов у зерен с низкой всхожестью.
В результате математического моделирования в среде MathCad с использованием эксперименталь- ных данных и системы уравнений были определены коэффициенты проницаемости оболочки семян пшеницы и концентрации ионов на внутренней стороне оболочки зерен [37].
Для зерен пшеницы со всхожестью 97% коэффициенты проницаемости составили: P k = 0,510±0,080, Р Na = 0,020±0,80, Р Cl = 0,450±0,070.
Относительные коэффициенты проницаемости составляют: Р к :Р Na :Р Сl = 1:0,04:0,88. При этом концентрация ионов калия равна 10–5 мг/л, концентрация ионов натрия -10–3 мг/л, концентрация ионов хлора -10–4 мг/л.
Для зерен пшеницы со всхожестью 87% коэффициенты проницаемости равны:
P k = 0,02600±0,00400, Р Na = 0,00010±0,00002, Р Cl = 0,10000±0,02000.
Относительные коэффициенты проницаемости составляют: Р к :Р Na :Р Сl = 0,26:0,001:1. При этом концентрация ионов калия составляет -10–7 мг/л, концентрация ионов натрия -10–5 мг/л, концентрация ионов хлора -10–9 мг/л.
Коэффициенты проницаемости у оболочек зерен пшеницы со всхожестью 87% значительно ниже, чем для зерен с высокой всхожестью 97%, что, в свою очередь, и дало подтверждение предложенной гипотезе.
На рисунке 2 представлены результаты применения раствора NaCl при проращивании зерен с различной всхожестью.
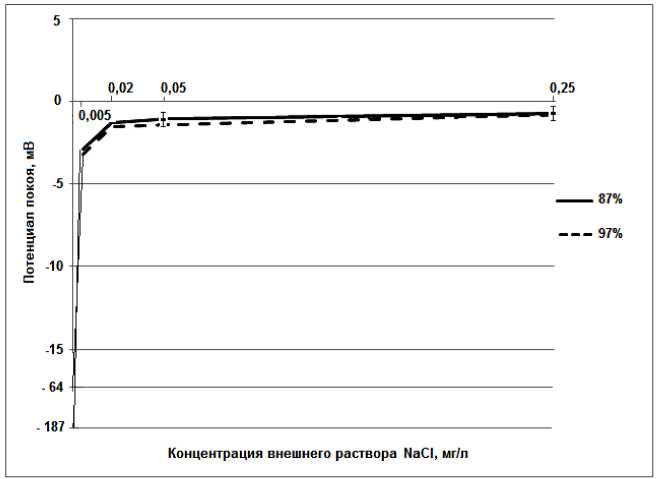
Рисунок 2 – Результаты экспериментальных исследований зерен пшеницы с разной всхожестью с использованием солевых растворов NaCl c различной концентрацией
Как видно из графиков, у зерен с обеими всхо-жестями наблюдаются существенные изменения значений ПП от -3 мВ до -1 мВ при изменении концентрации от 0,005 мг/л до 0,25 мг/л. При этом графики не имеют достоверного отличия.
Как следует из представленного уравнения, значение потенциала снижается при условии роста концентраций ионов Na+ и Cl , что инаблюдается на рисунке 2.
С применением раствора NaCl изменилась проницаемость оболочки зерен пшеницы, со всхожестью 97% коэффициенты проницаемости составили:
Р к = 0,0100±0,0035, Р Na = 0,0010±0,0004,
Р Cl = 0,4900±0,0600. При этом концентрация ионов калия составила -10-5 мг/л, концентрация ионов натрия -10-4 мг/л, концентрация ионов хлора -10-4 мг/л.
Для зерен пшеницы со всхожестью 87% коэффициенты проницаемости равны: Р к =0,00500±0,00100, Р Na =0,00010±0,00005, Р Cl =0,49000±0,00500. При этом концентрация ионов калия составила -10-7 мг/л, концентрация ионов натрия -10-5 мг/л, концентрация ионов хлора -10-9 мг/л.
Зерна пшеницы можно сравнить с клеткой живого организма, которая обладает мембранным потенциалом. Мембранный потенциал любой клетки используется для электрического транспорта извне веществ, необходимых клетке для ее жизнедеятельности [23]. В зерне роль мембраны выполняет оболочка. В зависимости от ее проницаемости, концентрации внутренних и внешних ионов на оболочке возникает ПП, который влияет на развитие зерна.
Как видно из приведенных данных, при замачивании зерен дистиллированной водой наибольший ПП возникает у зерен с низкой всхожестью. Низкая проницаемость оболочки и низкая концентрация внутренних ионов даже при малых внешних концентрациях растворов солей KCl и NaCl ведет к резкому уменьшению ПП, стремящегося к нулю. А при концентрации раствора KCl более 5·10-4 мг/л ПП приобретает положительные значения. Очевидно, что нулевые, а тем более положительные значения ПП не способны обеспечить электрический транспорт веществ через оболочку зерна. Электрический транспорт молекул необходимых веществ осуществляется посредством белка и присое- диненного иона натрия [23]. Если не будет ПП, то не будет движения ионов натрия через оболочку вовнутрь, а если ПП к тому же приобретает положительные значения, то возникающее поле вообще будет отталкивать ионы от оболочки.
У зерен с высокой всхожестью, при аналогичных внешних концентрациях раствора KCl сохраняется небольшой ПП. Однако и он монотонно стремится к нулю и при концентрации 0,5 мг/л приобретает значение нуля.
Воздействие внешнего раствора NaCl на зерна с различной всхожестью практически одинаково. Графики идут почти параллельно друг другу. Потенциалы покоя при концентрации 5·10-3 мг/л составляют -2 мВ, -3 мВ и практически монотонно уменьшаются до нуля при концентрации 0,25 мг/л.
Выводы. Основываясь на экспериментальных исследованиях, установлено, что электрические потенциалы зерен пшеницы могут быть использованы как показатели их качества. Установлено, что при проращивании семян в дистиллированной воде электрический потенциал покоя равен -64 мВ для семян с высокой всхожестью, для низкой – составил -187 мВ. Такая разница значений объясняется разной проницаемостью оболочки и концентрацией ионов Na+ , K+ и сг у зерен с высокой и низкой всхожестью.
Опираясь на уравнение Гольдмана–Ходжкина– Катца, где значение потенциала характеризуется проницаемостью мембраны и концентрацией ионов, была выдвинута гипотеза о разной проницаемости оболочки семян пшеницы разной всхожести. Чем выше всхожесть семян, тем выше проницаемость оболочки, а следовательно, и ниже значение потенциала.
Применение солевых растворов KCl, NaCL вызвало изменение потенциалов у зерен как с низкой, так и высокой всхожестью. При этом значения потенциалов у семян с низкой всхожестью ниже. При использовании раствора KCl зерна пшеницы со всхожестью 87% отличаются положительными значениями потенциалов в отличие от значений потенциалов семян со всхожестью 97%, разница значений объясняется разной проницаемостью оболочки у семян (у семян с низкой всхожестью проницаемость ниже). При использовании раствора NaCL потенциал у зерен со всхожестью 87% и 97% с увеличением концентрации раствора приближается к нулю. В сельскохозяйственной практике почвы, содержащие NaCl, называют засоленными. Эти соли вредят нормальному развитию растений.
Полученные результаты исследований свидетельствуют о целесообразности применения электрических потенциалов семян пшеницы для экспресс-диагностики их всхожести.
Список литературы Исследование электрических сигналов зерен пшеницы
- Fromm J., Lautner S. Electrical signals and their physiological significance in plants. Plant Cell Environ. 2007. -30, P. 249-257.
- Martonosi, A.N. Animal electricity, Ca2+ and muscle contraction. A brief history of muscle research. ActaBiochim. 2000. -Pol. 47, P. 493-516.
- О природе потенциала действия высших растений/С.С. Пятыгин, В.А. Опритов, А.В. Половинкин, В.А. Воденеев//ДАН. -1999. -C. 404-407.
- Opritov, V.A. Electrical component of general adaptation syndrome of higher plants/V.A. Opritov, S.S. Pyatygin, V.A. Vodeneev//Plants under environmental stress: Abstr. Int. Symp. Moscow, 2001. -P. 215.
- The role of bioelectric potentials in reception of cooling in higher plants/V.A. Opritov, S.S. Pyatygin, V.A. Vodeneev, S.A. Mysyagin//Signaling Systems of Plants Cells: Abstr. In-tern. Symp. Pushino: ONTI, 2001. P. 95-96.
- Opritov V.A., Pyatygin S.S., Retivin VG (1991) Bioelectrogenesis in higher plants. Nauka, Moscow.
- Медведев, С.С. Физиологические основы полярности растений. Труды СПб о-ва естествоиспытателей/С.С. Медведев. -СПб.: Кольна, 1995. -159 с. 81.
- Медведев, С.С. Электрофизиология растений/С.С Медведев. -СПб.: Изд-во С.-Петербургского университета, 1997. -122 с. 82.
- Медведев, С.С. Электрические поля и рост растений/С.С. Медведев//Электронная обработка материалов, Кишинев. -1990. -№ 3. -С. 68-74.
- Крутецкая, З.И. Биофизика мембран/З.И. Крутецкая, А.В. Лонский. -СПб.: Изд-во СПбГУ, 1994. -287 с.
- Lautner S., Grams T.E.E., Matyssek R. & Fromm J. Characteristics of electrical signals in poplar and responses in photosynthesis. Plant Physiology 138. -2005. -P. 2200-2209.
- Fromm J. Long-distance electrical signalling and its physiological functions in higher plants. In Plant Electrophysiology (ed. A.G. Volkov), Springer-Verlag, Berlin and Heidelberg, Germany, 2006. -pp. 269-285.
- Fromm J. &Lautner S. Characteristics and functions of phloem-transmitted electrical signals in higher plants. In Communication in Plants -Neuronal Aspects of Plant Life (eds F. Baluska, S. Mancuso & D. Volkmann). Springer Verlag, Berlin and Heidelberg, Germany. -2006. -pp. 321-332.
- Trebacz K., Dziubinska H. &Krol E. Electrical signals in long-distance communication in plants. In Communication in Plants -Neuronal Aspects of Plant Life (eds F. Baluska, S. Mancuso & D. Volkmann), Springer-Verlag, Berlin and Heidelberg, Germany. -2006. -pp. 277-290.
- Surova L., Sherstneva O., Vodeneev V., Katicheva L., Semina M., Sukhov V. Variation potential-induced photosynthetic and respiratory changes increase ATP content in pea leaves. J. Plant Physiol. 202, 2016. -pp. 57-64.
- Экспериментальная установка для исследования потенциала действия зерен пшеницы/М.В. Шереметьев, А.А. Зырянов, Н.Н. Мерченко, А.Г. Зрюмова, С.П. Пронин//Ползуновский альманах. -2011. -№ 1. -С. 177-178.
- Барышева, Н.Н. Метод контроля всхожести семян пшеницы по изменению мембранного потенциала/Н.Н. Барышева, С.П. Пронин//Ползуновский вестник. -2015.
- Мерченко, Н.Н. Зависимость мембранного потенциала зерен пшеницы от концентрации ионов на внутренней стороне оболочки и ее проницаемости/Н.Н. Мерченко, С.П. Пронин//Фундаментальные исследования. -2014. -№ 8. -С. 1539-1544. 88 с.
- Véry A-A, Sentenac H. Molecular mechanisms and regulation of K+transport in higher plants. Annual Reviews of Plant Biology 54. -2003. -P. 575-603.
- Demidchik V, Tester MA. Sodium fluxes through nonselective cationchannels in the plant plasma membrane of protoplasts from Arabidopsis roots. Plant Physiology 128, 2002. -pp. 379-387.
- Elzenga JTM, van Volkenburgh E. Characterization of ion channelsin the plasma membrane of epidermal cells of expanding pea (Pisumsativumarg) leaves. Journal of Membrane Biology 137, 1994. -P. 227-235.
- Maathuis FJM, Sanders D. Energization of potassium uptake inArabidopsis thaliana. Planta 191, 1993. -P. 302-307.
- Беркинблит, М.Б. Электричество в живых организмах/М.Б. Беркинблит, Е.Г. Глаголева. -М.: Наука, 1988. -2
