Исследование микробного профиля структурированной ассоциативной культуры микроорганизмов – кефирных грибков

Автор: Градова Нина Борисовна, Саранцева Александра Алексеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Коррекция экологического неблагополучия
Статья в выпуске: 5-3 т.14, 2012 года.
Бесплатный доступ
Исследован микробный профиль кефирных грибков из четырёх техногенных ниш. При исследовании 55 изолятов дрожжей не выявлено культур, обладающих β-галактозидазной активностью, способных использовать лактозу для спиртового брожения. При исследовании 39 изолятов бактерий выявлено присутствие двух физиологических групп молочнокислых бактерий: обладающих и не обладающих β-галактозидазной активностью. Использование метода ПЦР для идентификации выделенных молочнокислых бактерий показало, что использование методов через «выделение чистых культур» не позволяет получить объективную характеристику микробного профиля кефирных грибков. Сравнение грибков из двух техногенных ниш при использовании метода денатурирующего градиентного гель электрофореза («без выделения чистых культур») показало их идентичность по составу молочнокислых бактерий и перспективность использования метода для разработки концептуальной модели биологической системы кефирных грибков.
Микробный профиль, ассоциативная культура, кефирные грибки, концептуальная модель
Короткий адрес: https://sciup.org/148201584
IDR: 148201584 | УДК: 579.67:579.262
Research the microbic profile of the structured associative culture of microorganisms – kefiric grains
The microbic profile of kefiric grains from four technogenic niches is investigated. At research 55 isolates of yeast it is not revealed the cultures, possessing β-galactosidase activity, capable to use lactose for spirit fermentation. At research 39 isolates of bacteria presence of two physiological groups of lactic bacteria is revealed: possessing and not possessing β-galactosidase activity. PCR method used for identification of the allocated lactic bacteria showed that using the methods through «allocation of pure cultures» doesn't allow to receive the objective characteristic of microbic profile of kefiric grains. Comparison of grains from two technogenic niches when using a method of denaturing gradient gel electrophoresis («without allocation of pure cultures») showed their identity on structure of lactic bacteria and prospects of using the method for development the conceptual model of biological system of kefiric grains.
Текст научной статьи Исследование микробного профиля структурированной ассоциативной культуры микроорганизмов – кефирных грибков
Изучение закономерностей формирования структуры и функционирования ассоциативных микробных культур имеет общебиологическое значение, поскольку расширяет представления о механизмах организации природных стабильных биологических систем, а также является основой для разработки технологии создания и управления практически ценными микробными сообществами, для обеспечения их стабильности и высокой активности. Одной из таких морфологически оформленных стабильно функционирующих ассоциативных культур микроорганизмов являются кефирные грибки, период использования которых человеком для получения кефира – продукта, обладающего высокой биологической ценностью, превышает тысячелетия. Кефирные грибки как практически используемая ассоциативная культура микроорганизмов давно привлекает внимание, как отечественных исследователей, так и зарубежных, однако закономерности формирования его структуры и функционирования остаются недостаточно изученными. При изучении микробного состава кефирных грибков установлено, что основными компонентами ассоциативной культуры являются микроорганизмы четырех физиологических групп: молочнокислые бактерии, осуществляющие гомо- и гете-роферментативное брожение, дрожжи, осуществляющие спиртовое брожение и уксуснокислые бактерии. Микробный профиль кефирных грибков, описанный разными авторами, значительно различается [1, 2]. В кефирных грибках описано присутствие более 20 видов молочнокислых бактерий разных родов, более 10 родов и видов дрожжей, 2 вида уксуснокислых бактерий. Среди дрожжей описаны культуры как способные, так и не способные использовать лактозу для брожения, а также одновременное присутствие тех и других [3]. На основании литературных данных можно было бы предполагать, что кефирные грибки представляют систему, разные трофические структуры которой могут выполнять одинаковые функции, однако для такого вывода, как и для построения концептуальной модели биологической системы кефирных грибков как единой системы отсутствуют необходимые данные. В частности, нет ясности в определении продуцента данной системы, закономерностях трофических отношений между молочнокислыми бактериями разных родов и видов.
Цель работы : изучение микробного профиля кефирных грибков из разных техногенных ниш и потока веществ между микробными компонентами, как основы для построения концептуальной модели системы.
Объекты и методы. Объектами исследовании являлись нативные кефирные грибки, используемые для получения кефира на молочных предприятиях Ставрополя (КГ-1), Гагарина (КГ-2) и лиофилизированные кефирные грибки, используемые на предприятиях Владикавказа (КГ-3) и Москвы (КГ-4). В лабораторных условиях кефирные грибки поддерживали на стерильном молоке с содержанием жира 0,5-1,5%. Объем зерен определяли по объему вытесняемой жидкости при их погружении в молоко. Для определения микробного состава проводили высев из кефирных зёрен, последовательно промытых стерильным молоком, 1% раствором молочной кислоты, гомогенизированных (растирание в ступке) и суспендированных в 1% растворе триптона, на традиционно используемые твёрдые среды:
-
- дрожжевая среда(105), г/л: глюкоза (лактоза) – 100, дрожжевой экстракт - 10, мел - 20;
-
- среда Сабуро (глюкозо-пептонная), г/л: глюкоза - 40, пептон - 10, дрожжевой автолизат - 30;
-
- среда MRS, г/л: гидролизат казеина - 10, пептон - 10, ацетат натрия - 5, цитрат триаммония – 2,0, К 2 НРO 4 – 2,0, MgSO 4 – 0,2, MnSO 4 · 4Н 2 O – 0,05.
Посевы инкубировали при 27°С. Морфологические признаки колоний описывали при просмотре под микроскопом с увеличением ×400 и ×900. В качестве критерия оценки активности кефирных грибков как макросистемы и культур микроорганизмов, выделенных из них, использовали интегральные параметры их функциональной активности: изменение рН среды, титруемой кислотности при определённом времени экспозиции в молоке, скорость образования сгустка и его характер, а также наличие β-галактозидазной активности. Титруемую кислотность молока выражали в градусах Тернера (°Т). β-галактозидазная активность микроорганизмов определялась по их кислотообразующей активности [4] и при использовании хромогенного субстрата X-Gal [5]. Активность спиртового брожения определяли в сосудиках Эйнгорна-Смитта и выражали в относительных единицах 1, 1/2, 1/3 и 1/6 (по объему вытесненной жидкости). Для идентификации бактериального компонента консорциума использовали метод ПЦР, и метод ДНК-ДНК гибридизации – для дрожжей. Для сравнения микробного профиля кефирных грибков использовали метод денатурирующего градиентного гель электрофореза (DGGЕ).
Результаты и обсуждение. Исследуемые кефирные грибки представляли желеобразные гранулы неправильной формы со складчатой и бугристой поверхностью, различающиеся по размеру и упругости (табл. 1). Средний объём кефирных грибков популяции КГ-1 и КГ-2 составлял около 1,0 см3, а КГ-3 и КГ-4 около 0,5 см3, что соответствует диапазону размеров кефирных грибков, описанному в литературе. Содержание азота в кефирных грибках, косвенно указывающее на микробную их составляющую, практически было одинаковым: 4,7-5,0%, что также соответствовало количеству азота 4,54,8%, определяемого в кефирных грибках, отобранных на разных предприятиях в Аргентине [2].
Таблица 1. Характеристика кефирных грибков
|
Кефирные грибки |
КГ-1 |
КГ-2 |
КГ-3 |
КГ-4 |
|
объем, см3 |
1,1±0,03 |
0,95±0,02 |
0,52±0,02 |
0,47±0,03 |
|
содержание азота, % |
4,7 |
5,0 |
4,9 |
4,8 |
|
упругость |
+++ |
+++ |
++ |
+ |
При инкубировании в молоке единичных кефирных грибков небольшого размера (рис. 1) было показано, что в течение 1-2 суток объём грибков увеличивался до средних размеров, характерных для культивируемых популяций, после чего от материнских зёрен отделялось 1-5
мелких образований. Общий объём массы кефирных грибков в дальнейшем увеличивался за счёт роста отделившихся мелких образований, а объём материнского грибка медленно увеличивался, приближаясь к первоначальному. Наблюдаемая динамика роста кефирных грибков показывает, что их микробное сообщество способно взаимодействовать со средой обитания как некое кооперативное целое.
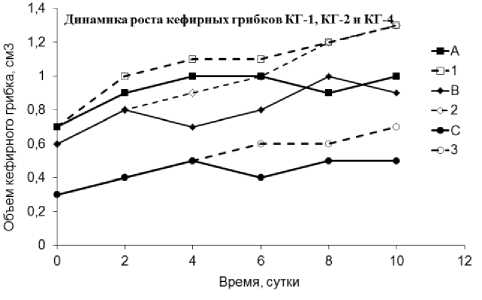
Рис. 1. Динамика роста кефирных грибков:
А – КГ-1, В - Г-2 и С – КГ-4 – объём кефирных грибов;
1, 2 и 3 – объём общей массы кефирных грибков
Исследование микробного состава изучаемых кефирных грибков классическими методами при высеве отмытых, растёртых и суспендированных в триптоне грибков на твёрдые среды показало, что количество дрожжей в ассоциативной культуре было практически в 10 раз меньше, чем бактерий (табл. 2). В процессе работы было отмечено, что при подготовке грибков к высеву, их растиранию необходимо учитывать степень их упругости. При повторном растирании наиболее упругих грибков количество высеваемых микроорганизмов увеличивалось на 15-20%, что нами учитывалось в дальнейшей работе.
Учитывая данные литературы о разной способности дрожжей, выделенных из кефирных грибков, к использованию лактозы в качестве субстрата для спиртового брожения, а также исходя из предположения о возможности дрожжей, образующих морфологически однотипные колонии, обладать разными физиологобиохимическими свойствами, были исследованы все выделенные изоляты из обособленно стоящих колоний, морфологически характерных для дрожжей. Всего было исследовано 23 изолята из КГ-1 и 32 из КГ-2. При микроскопировании первично выделенных изолятов дрожжей было выявлено их инфицирование бактериями, чаще кокковой формы. При рассеве изолятов на среду Сабуро также не были выделены чистые культуры дрожжей из многих изолятов, что может свидетельствовать о тесных симбиотических взаимоотношениях дрожжей и бактерий. При использовании метода дисков, пропитанных ампи-цилином, из колоний, выросших близ диска, были выделены чистые культуры дрожжей.
Таблица 2. Количество дрожжей и бактерий в кефирных грибках, (КОЕ/мл)
|
Кефирные грибки |
КГ-1 |
КГ-2 |
КГ-3 |
КГ-4 |
|
дрожжи |
4,1 · 104 |
2,8 · 104 |
9,4 · 103 |
9,3 · 103 |
|
бактерии |
9,8 · 104 |
1,2 · 105 |
2,1 · 105 |
3,2 · 105 |
Исследование физиолого-биохимических свойств выделенных 55 чистых культур дрожжей показало, что все дрожжи аспорогенны, при их инкубировании в молоке не наблюдалось снижения рН среды, повышения показателей титруемой кислотности, не образовывался сгусток. Изоляты дрожжей не обладали β-галактозидазной активностью. Определение функциональной активности изолятов дрожжей, инфицированных бактериями, показало, что функциональная активность изолятов выше, чем выделенных из них чистых культур бактерий (табл. 3), что свидетельствует о тесных симбиотических отношениях молочнокислых бактерий и дрожжей.
Симбиотические отношения дрожжей и бактерий были показаны также в опыте при использовании метода накопительных культур. При культивировании в микроаэрофильных условиях растёртых кефирных грибков КГ-1 и КГ-
-
2 на минеральной среде, содержащей разные источники углерода: лактозу (при разных содержаниях дрожжевого автолизата), глюкозу, галактозу и этанол.
Таблица 3. Функциональная активность изоля-тов дрожжей, инфицированных бактериями, и чистых культур дрожжей
|
Объект |
Время образ. сгустка, сут. |
Характер сгустка |
Индекс лакто-зос-бражи-вания |
|
|
изолят 1 |
5 |
+++ |
120 |
1,0 |
|
ч.к. бакт. 1 |
7 |
++ |
110 |
0,89 |
|
изолят 2 |
6 |
++ |
110 |
1,0 |
|
ч.к. бакт. 2 |
7 |
++ |
100 |
0,89 |
|
изолят 3 |
7 |
++ |
105 |
0,94 |
|
ч.к. бакт. 3 |
8 |
+ |
30 |
0,07 |
В четвёртом пассаже активность роста накопительных культур на лактозе, глюкозе и галактозе была, практически, одинакова. При этом не было выявлено конкурентного преимущества дрожжей или бактерий, компонентов кефирных грибков, при развитии на лактозе в присутствии высокой концентрации дрожжевого автолизата, глюкозе и галактозе (табл. 4). На среде с этанолом развивались только палочковидные бактерии, идентифицированные как бактерии p.Aceto-bacter. При отсутствии в среде автолизата и низком его содержании, 0,1г/л, на среде с лактозой развивались только дрожжи, что подтверждает большую роль продуктов автолиза дрожжей в трофической структуре микробного сообщества кефирных грибков и не дают оснований для определения продуцента микробной системы кефирных грибков при их развитии на молоке.
Таблица 4. Характеристика микробного состава накопительных культур
|
Лактоза Глюкоза Галактоза |
Этанол |
|
конц. дрож. автолиз., г/л 0 0,1 0,2-1,0 20 дрожжи и бактерии дрожжи дрожжи и бактерии |
бактерии |
Дрожжи, выделенные из накопительных культур на разных источниках углеводов, были исследованы на содержание ГЦ пар в ДНК. Результаты показали, что по показателю минимального содержания ГЦ пар и диапазону изменчивости дрожжи относились к роду Kluyveromyces , которые способны активно использовать лактозу в качестве источника углерода. Выделенные культуры дрожжей не использовали лактозу для спиртового брожения, а осуществляли с разной степенью активности спиртовое брожение на глюкозе. Культуры, характеризующиеся наиболее низкой активностью спиртового брожения на глюкозе, использовали галактозу для спиртового брожения (табл. 5). Таким образом, проведенные исследования показали, что дрожжи не являются продуцентом в микробной системе исследованных кефирных грибков, они не обладают β-галактозидазной активностью, не используют лактозу для спиртового брожения.
Таблица 5. Активность спиртового брожения дрожжей на разных углеводах (количество штаммов, %)
|
Акт-ть спирт. брожения, отн. ед. |
Количество штаммов, выделенных на средах с разными источниками углерода,% |
||
|
глюкоза |
галактоза |
лактоза |
|
|
1 |
37 |
0 |
0 |
|
1/2 |
27 |
0 |
0 |
|
1/3 |
18 |
18 |
0 |
|
1/6 |
9 |
9 |
0 |
Для исследования бактериального профиля ассоциативной культуры кефирных грибков из КГ-1, КГ-2 и КГ-4 из высевов на твёрдые среды были выделены чистые культуры из отдельно стоящих колоний, имеющих даже незначительные морфологические различия. Колонии молочнокислых бактерий характеризуются малым размером (диаметр большинства колоний от 1 до 2 мм) и сходной морфологией, что крайне затрудняет анализ высевов. На основании морфологии колоний и морфологии клеток было выделено 13 чистых культур из КГ-1, 12 - из КГ-2 и 14 - из КГ-4. Среди бактериальных изолятов на основании морфологии колоний, пигментации, образования зон просветления и характера роста были идентифицированы уксуснокислые бактерии. Среди молочнокислых бактерий, выделенных из кефирных грибков, выявлено несколько физиологических групп:
-
- палочковидные, Г+, каталазоотрицательные, осуществляющие гомо- и гетероферментативное молочнокислое брожение;
-
- палочковидные, каталазоположительные, Г+; - кокковидные, Г-, каталазоотрицательные.
При использовании метода ПЦР среди бактериальных культур, выделенных из разных кефирных грибков, были идентифицированы молочнокислые бактерии родов Lactococcus , Lactobacillus и Leuconostoc (табл. 6).
Наибольшее видовое разнообразие было отмечено среди бактерий родов Lactobacillus и Leuconostoc . Исследование функциональных свойств выделенных молочнокислых бактерий показало, что не все молочнокислые бактерии обладали β-галактозидазной активностью. Так, выделенные из всех изученных кефирных грибков культуры Lactobacillus plantarum и другие виды этого рода, выделенные из КГ-4, не обладали β-галактозидазной активностью. Бактерии Lactococcus lactis , Lactobacillus sakei и Leuconostoc mesenteroides , выделенные из всех трёх видов изученных кефирных грибков, характеризовались высокой β-галактозидазной активностью, малым временем образования сгустка и способностью снижать рН среды до 2-3.
Таблица 6. Бактериальный профиль кефирных грибков
|
Бактерия |
% сов-падений |
Присутствие в кефирных грибках |
Активность |
||||
|
КГ-1 |
КГ-4 |
КГ-2 |
время образов. сгустка, сут. |
рН |
β-галактоз. акт-ть |
||
|
Lactococcus lactis |
91-93 |
+ |
+ |
+ |
2 |
2-3 |
7 |
|
Lactobacillus sp |
86-89 |
+ |
- |
- |
7 |
3 |
0 |
|
Lactobacillus lactis |
- |
- |
+ |
1 |
2-3 |
7 |
|
|
Lactobacillus delbrueckii |
98 |
+ |
- |
- |
5 |
5 |
7 |
|
Lactobacillus plantarum |
93 |
+ |
+ |
+ |
1 |
2 |
0 |
|
Lactobacillus sakei |
99 |
+ |
+ |
+ |
1 – 2 |
4 |
7 |
|
Lactobacillus otakiensis |
94 |
- |
+ |
- |
0 |
0 |
0 |
|
Lactobacillus kefiri |
94 – 99 |
- |
+ |
- |
0 |
5-6 |
0 |
|
Leuconostoc gelidum |
94 |
+ |
- |
- |
1 |
4 |
7 |
|
Leuconostoc mesenteroides |
98-99 |
+ |
+ |
+ |
1-3 |
2-4 |
7 |
|
Leuconostoc lactis |
- |
- |
+ |
2 |
4 |
7 |
|
|
Streptococcus lactis |
+ |
- |
+ |
2-3 |
4-5 |
7 |
|
|
Acetobacter aceti |
95 |
+ |
+ |
+ |
0 |
0 |
0 |
|
Acetobacter sp |
96-98 |
+ |
+ |
+ |
0 |
0 |
0 |
Таким образом, полученные данные показывают присутствие в бактериальной компоненте кефирных грибков молочнокислых бактерий разных видов, способных осуществлять молочнокислое брожение при использовании лактозы (обладающие β-галактозидазной активностью), так и не использующие лактозу для молочнокислого брожения. Это свидетельствует о сложных трофических взаимоотношениях между культурами молочнокислых бактерий, которые могут быть либо конкурентными за субстрат, либо кооперативными в его использовании. Однако данные, характеризующие различия в бактериальном профиле разных кефирных грибков, являются недостаточно корректными, поскольку они были получены на основе «выделенных чистых культур», что может вносить ошибку в результаты в связи с особенностями морфологии колоний молочнокислых бактерий и способностью роста ряда видов молочнокислых бактерий на специфических средах. Так, в процессе экспериментов было показано, что культуры, выделенные из морфологически казалось однотипных колоний, по данным реакции ПЦР относились к разным видам молочнокислых бактерий и наоборот. Полученные данные подтверждают выводы ряда авторов о необходимости для изучения микробного профиля кефирных грибков использования методов «независимых от выделения чистых культур» [6, 7]. Помимо этого следует учитывать мнение отдельных авторов о большом значении в объективности получаемых данных методом ПЦР правильности подобранных праймеров.
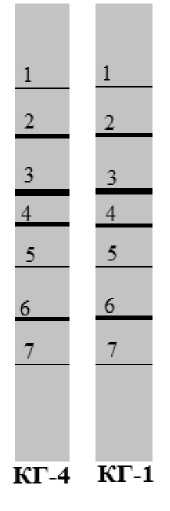
Рис. 2. Схема DGGE КГ-4, КГ-1
В связи с этим для сравнительной характеристики микробного профиля кефирных грибков КГ-1 и КГ-4 был использован метод денатурирующего градиентного гель электрофореза (DGGE), который позволяет определить микробный профиль консорциума микроорганизмов без выделения чистых культур. Результаты исследований (рис. 2, 3) показали практически идентичный микробный состав кефирных грибков, используемых для приготовления кефира на предприятиях Ставрополя и Москвы. Доминирующими по степени четкости выраженных полосок являются одни и те же 7 видов микроорганизмов. Отмечены лишь некоторые различия среди второстепенных микроорганизмов, образующих трудноразличимые полоски.
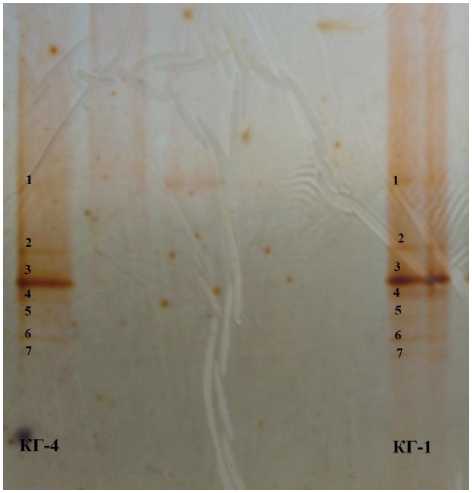
Рис. 3. Фореграмма DGGE КГ-4, КГ-1
Выводы:
-
1. Показано, что использование классических методов через «выделение чистых культур» не позволяет достоверно определить микробный профиль кефирных грибков, что связано с однотипностью морфологии колоний молочнокислых бактерий, потребностью для развития ряда бактерий специфических питательных сред и условий, а также тесными симбиотическими отношениями дрожжей и бактерий.
-
2. На основании изучения изолятов дрожжей двух образцов кефирных грибков, выделенных из всех выросших колоний морфологически характерных для дрожжей, показано, что присутствующие в микробном сообществе дрожжи не обладают в-галактозидазной активностью, в качестве субстрата для спиртового брожения не используют лактозу, активно используют глюкозу и с низкой активностью галактозу.
-
3. При использовании метода ПЦР во всех трёх исследованных образцах кефирных грибков определено присутствие бактерий:
-
- Lactococcus lactis , Lactobacillus sakei и Leuconostoc mesenteroides , обладающих в-галактозидазной активностью;
-
- Lactobacillus plantarum , не обладающих в-галактозидазной активностью;
-
- два вида уксуснокислых бактерий.
-
4. По функциональным свойствам выделенные молочнокислые бактерии можно подразделить на две группы:
- обладающие в-галактозидазной активностью, использующие в качестве субстрата для молочнокислого брожения лактозу, глюкозу и галактозу
- не обладающие в-галактозидазной активностью, активно осуществляющие молочнокислое брожение на глюкозе и с низкой активностью на галактозе.
-
5. При использовании метода денатурирующего градиентного гель электрофореза (без выделения чистых культур) определён идентичный микробный профиль двух образцов кефирных грибков, включающий семь доминирующих видов бактериальных культур.
В отдельных образцах выявлено присутствие разных видов бактерий рода Lactobacillus, Leuconostoc, способных осуществлять молочно- кислое брожение, но не обладающих в-галактозидазной активностью.
Заключение. На основании анализа и литературных данных и результатов проведенных исследований очевидно, что ассоциативная микробная культура кефирных грибков является высокоорганизованным сообществом, обладающим сложными вертикальными и горизонтальными трофическими связями. Присутствие в системе нескольких видов молочнокислых бактерий, обладающих в-галактозидазной активностью, даёт основание полагать о конкурентных взаимоотношениях между этими культурами за субстрат. Продуцентом системы является один из данных видов, обладающий наибольшей активностью использования лактозы в данных условиях, что подтверждается и в работе Хамнаевой Н.И. [8]. Исследованный механизм транспорта лактозы в клетки микроорганизмов и пути её метаболизма под действием фермента в-галактозидазы показывают возможность выхода в культуральную жидкость некоторого количества глюкозы, 2-дезокси-Д глюкозы и галактозы [9]. При этом показано, что только после исчерпания из среды глюкозы в клетках молочнокислых бактерий индуцируется синтез в-галактозидазы, что определяет более высокое содержание в среде галактозы по сравнению с глюкозой. Эти данные обосновывают возможность проявления конкурентных взаимоотношений также среди видов не обладающих в-галактозидазной активностью, но активно использующих глюкозу для молочнокислого брожения. Роль дрожжей в микробном сообществе кефирных грибков, очевидно, заключается не только в использовании глюкозы (и определённого количества галактозы) при кислой реакции среды для спиртового брожения, но в большой степени в стимулировании бактериальных культур продуктами метаболизма и автолиза. Образующийся спирт используется уксуснокислыми бактериями. Проведенные исследования показали, что достоверная характеристика микробного профиля кефирных грибков может быть получена при использование метода «без выделения чистых культур». Для разработки концептуальной модели ассоциативной культуры кефирных грибков необходимо провести сравнительные исследования микробного профиля их разных образцов. На основании проведенных исследований может быть представлена следующая схема трофической цепи в кефирных грибках (рис. 4).
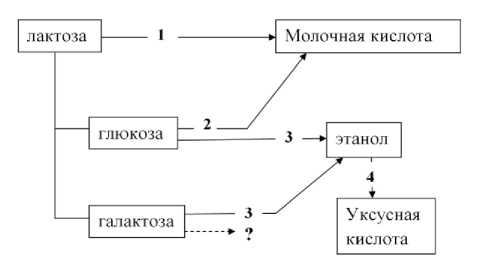
Рис. 4. Общая схема трофической цепи ассоциативной культуры кефирных зёрен:
1 –молочнокислые бактерии, синтезирующие β-галактозидазу; 2 –молочнокислые бактерии не синтезирующие β-галактозидазу; 3 – дрожжи; 4 – уксуснокислые бактерии
Список литературы Исследование микробного профиля структурированной ассоциативной культуры микроорганизмов – кефирных грибков
- Lopitz-Otsoa, R. Kefir: А symbiotic yeasts-bacteria community with alleged healthy capabilities/F. Lopitz-Otsoa, A. Rementeria, N. Elguezabal, J. Garaizar//Rev. Iberoam Micol. 2006. 23. P. 67-74.
- Garrote, G.L. Chemical and microbiological characterisation of kefir grains/G.L. Garrote, A.G. Abraham, G.L. de Antoni//Journal of Dairy Research. 2001. 68. P. 639-652.
- Chen, H-Ch. Microbiological study of lactic acid bacteria in kefir grains by culture-dependent and culture-independent methods/H-Ch. Chen, Sh.-Y. Wang, M.-J. Chen//Food Microbiology. 2008. 25. P. 492-501.
- Ганина, В.И. β-галактозидазная активность молочнокислых бактерий и бифидобактерий/В.И. Ганина, Л.В. Калинина, Е.В. Большакова//Молочная промышленность. 2002. №8. С. 36-37.
- Миллер, Дж. Эксперименты в молекулярной генетике. -М.: Мир, 1976. 436 c.
- Chen, T-H. Microbiological and chemical properties of kefir manufactured by entrapped microorganisms isolated from kefir grains/T-H. Chen, S-Y. Wang, K-N. Chen et al.//Journal of Dairy Science. 2009. 92. P. 3002-3013.
- Елинов, Н.П. Микробиота природной ассоциации «Тибетский рис»/Н.П. Елинов, О.Г.Ларина//Проблемы медицинской микологии. 1999. Том. 1, №1. С. 51-56.
- Хамнаева, Н.И. Научные и практические основы использования биотехнологических свойств кефирных грибков. Автореферат диссертации на соиск. уч. степ. д.т.н. -М., 2001. 47 с.
- Молотов, С.В. Получение и свойства мутантов молочнокислых стрептококков, дефектных по способности к утилизации глюкозы/С.В. Молотов, Р.А. Алхимова, Н.В. Пименова, В.В. Суходолец//Биотехнология. 1994. №11-12. С. 9-12.
