Исследование обоняния серых и гренландских тюленей с помощью метода оперантного обучения

Автор: Литвинов Юрий Владимирович, Пахомов Мирон Владимирович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Морская зоология
Статья в выпуске: 2 т.21, 2018 года.
Бесплатный доступ
Представлены результаты исследования обоняния серых и гренландских тюленей в процессе выработки двигательного условного рефлекса на запах объекта; установлена способность подопытных тюленей дифференцировать различные запахи. Исследование включало три этапа, в ходе которых тюленей обучали дифференцировать: 1) контейнер с раствором камфоры от контейнера с дистиллированной водой; 2) контейнер с раствором диметилсульфида (ДМС) от контейнера с дистиллированной водой (на этих этапах определяли возможность выработки у исследуемых тюленей двигательного условного рефлекса на обонятельный раздражитель); 3) при парном предъявлении контейнер с раствором диметилсульфида от контейнера с раствором камфоры (выявляли способность тюленей различать запахи). Эксперимент проводили в условиях аквакомплекса ММБИ (в Кольском заливе); в качестве подопытных животных были выбраны две самки серого тюленя ( Halichoerus grypus, Fabricius, 1791) в возрасте семи лет и две самки гренландского тюленя ( Pagophilus groenlandicus, Erxleben, 1777) в возрасте двух лет. Для проведения экспериментов стимульные вещества разводили в дистиллированной воде до такой концентрации, чтобы интенсивность запаха была минимальна, затем раствор помещали в контейнер. Результаты работы продемонстрировали способности подопытных тюленей вырабатывать двигательный условный рефлекс на обонятельные раздражители и отличать один запах от другого. Скорость обучения серых и гренландских тюленей в процессе дифференцировки контейнера с запахом от пустого контейнера была сравнительно одинакова, а при дифференцировке двух запахов гренландские тюлени обучались на порядок медленнее серых тюленей. Данное отличие в скорости обучения, вероятно, связано с видовыми особенностями исследуемых тюленей и свидетельствует о более высокой пластичности нервной системы серых тюленей.
Серый тюлень, гренландский тюлень, обоняние, камфора, диметилсульфид
Короткий адрес: https://sciup.org/142215146
IDR: 142215146 | УДК: 57.045 | DOI: 10.21443/1560-9278-2018-21-2-336-343
Investigation of olfaction of gray and harp seals by the operant training method
The results of studying the possibility of producing a motor conditioned reflex in the gray and harp seals on the smell of an object and the ability of experimental seals to differentiate certain odours have been presented. The study has contained three stages. In the first stage the seals were trained to differentiate a container from the camphor solution from a container with distilled water. In the second - to differentiate the container with a solution of dimethyl sulfide from the container with distilled water. At these stages, the possibility of developing a conditioned motor reflex to the olfactory stimulus in the seals studied has been studied. At the third stage, the ability of experimental animals was examined by paired presentation of a container with the solution of dimethyl sulphide from a container with camphor solution. The experiment has been carried out under the MMBI aquacomplex in the Kola Bay. The experimental animals were two female gray seal ( Halichoerus grypus, Fabricius, 1791) at the age of 7 years and two females of harp seal ( Pagophilus groenlandicus, Erxleben, 1777) at the age of 2 years. For the experiments, the stimulants were diluted in distilled water to such concentration that the odour intensity was minimal, then the solution was placed in a container. The results of the work have demonstrated the ability of experimental seals to develop a motor conditioned reflex to olfactory stimuli and the ability to distinguish one odour from another. When teaching the differentiation of a container with a smell, the rate of training of gray and harp seal is relatively the same, while learning to differentiate two smells, harp seals are trained an order of magnitude slower than gray seals. This difference in the speed of training is probably due to the specific features of the seals studied and indicates a higher plasticity of the nervous activity of gray seals.
Текст научной статьи Исследование обоняния серых и гренландских тюленей с помощью метода оперантного обучения
Морские млекопитающие долгое время рассматривались как аносматики или микросматики [1; 2]. У китообразных обоняние почти утрачено; обонятельные доли мозга и обонятельные нервы отсутствуют полностью (зубатые киты) или имеются в зачаточном состоянии (усатые киты) [3]; также в их геноме утрачены или находятся в неактивном состоянии гены, отвечающие за обоняние [4].
Наблюдения за поведением ластоногих в естественных условиях позволяют сделать вывод, что обоняние, возможно, играет важную роль в их социальной коммуникации, репродуктивном поведении [5] и выборе пищи [6].
Обонятельное поведение и назо-назальные контакты южноафриканских морских котиков (Arctocephalus pusillus, Schreber, 1775), содержащихся в неволе, были описаны G. J. Ross [7]. Самки гренландского тюленя (Pagophilus groenlandicus, Erxleben, 1777) определяли своих щенков визуально, вокально и с помощью обоняния, но окончательное подтверждение наблюдалось только после обнюхивания щенков [8]. Кормлению почти всегда предшествововал назо-назальный контакт [8; 9], в течение первых двух дней после рождения самки обнюхивали щенков значительно чаще, чем во время остального периода грудного вскармливания [8]. Самки северного морского котика (Callorhinus ursinus, Linnaeus, 1758) также часто обнюхивают щенков после рождения и во время периода лактации и используют назо-назальный контакт как средство идентификации своих щенков [10]. Этот же контакт как заключительное признание самкой своего щенка был описан у южноамериканских морских котиков (Arctocephalus australis, Zimmermann, 1783) [11], кергеленских морских котиков (Arctocephalus gazella, Peters, 1875) [12], австралийских морских львов (^eophoca cinerea, Peron, 1816) [13] и сивучей (Eumetopias jubatus, Schreber, 1776) [14].
Важную роль обоняние играет и в репродуктивном поведении морских млекопитающих. Самцы новозеландских морских котиков (Arctocephalus forsteri, Lesson, 1828) обнюхивали самок в области головы и задней части тела в период размножения, по-видимому оценивая готовность самок к спариванию [15]. Обонятельное поведение сивучей различалось как между животными разного пола и возраста, так и в разные периоды их жизни (репродуктивный и пострепродуктивный) [16]. У самцов кольчатых нерп (Phoca hispida, Schreber, 1775) в период размножения усиливалась секреция сальных и апокринных желез, которые издавали сильный специфический аромат [17]. Возможно, он используется для маркировки территории и/или для привлечения самок [18].
Южноафриканские морские котики могли дифференцировать запах рыбы от аромата эфирных масел, а также различать виды рыб по запаху [6].
В научной литературе представлены экспериментальные исследования физиологии и поведения ушастых тюленей; работы по изучению настоящих тюленей представлены недостаточно и в основном посвящены рассмотрению результатов наблюдения за поведением животных. По этой причине нами было проведено исследование обоняния представителей двух видов настоящих тюленей с использованием метода оперантного обучения.
Материалы и методы
Эксперимент проводили в условиях аквакомплекса ММБИ на акватории Кольского залива; подопытные животные – две самки серого тюленя (Halichoerus grypus, Fabricius, 1791 ) в возрасте семи лет и две самки гренландского тюленя (Pagophilus groenlandicus, Erxleben, 1777) в возрасте двух лет. К началу экспериментальных работ все животные были здоровы и обучены по схеме "выбор из двух стимулов".
В качестве обонятельных стимулов использовались 10%-е растворы камфорного масла и диметилсульфида. Запах камфоры был выбран потому, что он, во-первых, согласно современной стереохимической теории обоняния относится к первичным запахам, а во-вторых, тюлени, обитающие в Арктике, не могли сталкиваться с данным запахом ранее, поэтому камфора являлась для них индифферентным раздражителем. Запах диметилсульфида, предположительно, имеет важное биологическое значение для ластоногих. Диметилсульфид образуется в ходе метаболического разложения диметилсульфониопропионата, выделяемого в воду фитопланктоном в период его цветения [19]. Установлено, что северная качурка (Oceanodroma leucorhoa, Vieillot, 1818) [20], странствующий альбатрос (Diomedea exulans, Linnaeus, 1758) [21] и голубой буревестник (Halobaena caerulea, Gmelin, 1789) [22] находят места кормления по запаху диметилсульфида. Пингвины резко увеличивают свою двигательную активность в местах распыления ДМС [23]. Возможно, что по наличию в воде ДМС находят места скопления планктона некоторые виды акул [24] и рифовых рыб [25]. Экспериментально выявлена высокая чувствительность обыкновенных тюленей (Phoca vitulina, Linnaeus, 1758) к диметилсульфиду и установлено, что она на порядки выше, чем у людей [26].
Эксперимент был разделен на три этапа. На первом этапе животных обучали дифференцировать контейнер с раствором камфоры от пустого контейнера; на втором – дифференцировать контейнер с раствором ДМС от пустого контейнера; на третьем – дифференцировать контейнер с ДМС от контейнера с раствором камфоры. Для проведения экспериментов стимульные вещества разводили в дистиллированной воде до такой концентрации, чтобы интенсивность запаха была минимальна, затем раствор помещали в контейнер.
Экспериментальная установка представляла собой две трубы, закрепленные на деревянном брусе (рис. 1). Трубы имели длину 30 см и диаметр 6 см, благодаря чему тюлени могли свободно просовывать в них переднюю часть головы и дышать. В каждой трубе было вырезано отверстие диаметром 8 см, в которое помещались контейнеры с тестовыми веществами. С противоположного конца трубы закреплен электрический вентилятор, который создавал ток воздуха в трубе, что позволяло минимизировать влияние таких факторов, как ветер и посторонние запахи.

Рис. 1. Экспериментальная установка Fig. 1. Experimental installation
Экспериментальную установку помещали в вольер на расстоянии 1 м от стартовой позиции животного. Тюленей обучали по команде засовывать переднюю часть головы в трубу и оставаться в таком положении до 10 с. Также животных приучали к звуку работающего вентилятора.
В контейнер помещали тестовое вещество (положительный стимул), в другой контейнер – дистиллированную воду (отрицательный стимул) и устанавливали их в трубы. На третьем этапе, когда изучалась способность исследуемых тюленей дифференцировать между собой растворы диметилсульфида и камфоры, в один контейнер помещали диметилсульфид (положительный стимул), в другой – камфору (отрицательный стимул). Взаиморасположение контейнеров меняли в случайном порядке, но не более двух одинаковых комбинаций подряд, чтобы избежать "ассоциации с местоположением" у исследуемых тюленей.
Тюлень занимал стартовую позицию и по команде должен был найти трубу с положительным стимулом и зафиксировать переднюю часть головы в нем на 10 с. Животному разрешалось обследовать обе трубы. В случае правильного выполнения следовало пищевое подкрепление в виде рыбы. Эксперименты проводились в утреннее (10:00) и вечернее (17:00) время.
С животными проводили обучающие тренировки, когда тренер подталкивал к правильному выбору; количество предъявлений за тренировку не лимитировалось; учитывался только результат последней тренировки, который должен быть выше вероятности случайного выбора. Готовность к проведению контрольных испытаний определял тренер тюленя. Обучающие тренировки проводили в осенне-зимний период при стабильной безветренной погоде без атмосферных осадков и полном отсутствии посторонних запахов.
В ходе контрольных испытаний проводилось по 20 тренировок с 20 демонстрациями пар стимулов на каждой; за верные выборы стимула тюлень получал фиксированное пищевое поощрение. Тренировки осуществлялись по возможности при полном штиле (или максимум при 2 баллах по шкале Бофорта) и отсутствии сильных посторонних запахов, атмосферных осадков и иных раздражителей. Перед началом тренировки с тюленем проводились тестовые испытания для определения уровня работоспособности, состоящие из команд на возбуждение и торможение ЦНС и команд по переходу из состояния возбуждения в состояние торможения и обратно. К тренировке приступали только при высоком уровне работоспособности животного. Эти условия соблюдались для того, чтобы минимизировать влияние внешних факторов среды и изменений состояния подопытного животного на количество верных выборов.
Результаты и обсуждение
На первом этапе обучение серого тюленя 1 заняло шесть тренировок (в результате наблюдалось 75 % верных выборов), серого тюленя 2 – три тренировки (80 %), гренландского тюленя 1 – семь тренировок (80 %), гренландского тюленя 2 – семь тренировок (75 % верных выборов).
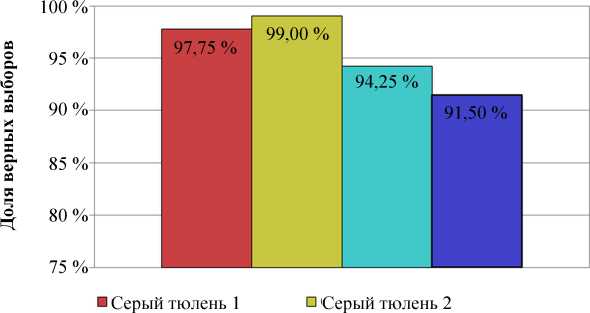
При проведении контрольных испытаний все тюлени достоверно отличали контейнер с раствором камфоры от ложного контейнера (рис. 2), при этом серые тюлени в среднем совершали одну ошибку за тренировку, гренландские тюлени – две-три ошибки за тренировку.
□ Гренландский тюлень 1 □ Гренландский тюлень 2
Рис. 2. Доля верных выборов при дифференцировке контейнера с раствором камфоры от пустого контейнера
Fig. 2. The proportion of correct choices when differentiating a container with a solution of camphor from an empty container
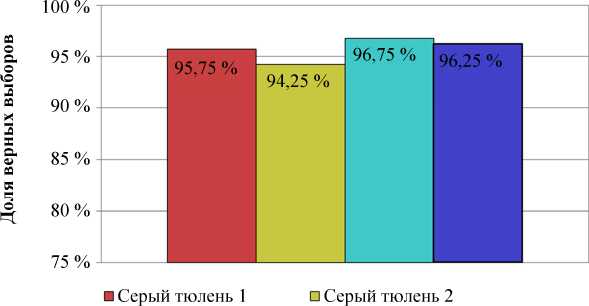
На втором этапе серый тюлень 1 обучился отличать контейнер с раствором ДМС в ходе четвертой тренировки (75 % верных выборов), серый тюлень 2 – седьмой тренировки (75 %), гренландские тюлени обучились во время двенадцатой тренировки (80 % верных выборов) (рис. 3). При проведении контрольных испытаний все тюлени достоверно отличали контейнер с раствором ДМС от пустого контейнера. При этом серые тюлени в среднем совершали по две ошибки за тренировку, гренландские – по одной ошибке.
□ Гренландский тюлень 1 □ Гренландский тюлень 2
Рис. 3. Доля верных выборов при дифференцировке контейнера с раствором ДМС от пустого контейнера Fig. 3. The proportion of correct choices when differentiating a container with a solution of LCA from an empty container
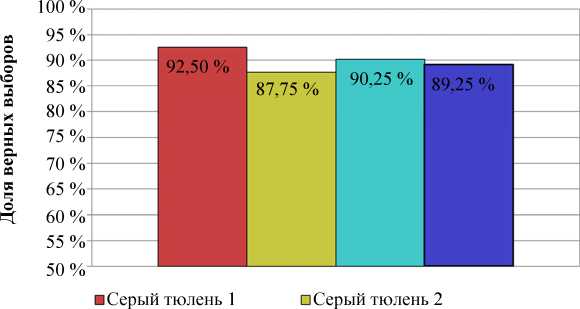
На третьем этапе серые тюлени уже на первой тренировке достоверно отличали контейнер с раствором ДМС от контейнера с раствором камфоры, при этом запах ДМС тюлени выбирали с первого предъявления пары стимулов. У серого тюленя 1 результат составил 95 % верных выборов, у серого тюленя 2 – 90 %. Обучение гренландского тюленя 1 заняло 20 тренировок (75 % верных выборов); гренландского тюленя 2 – 21 тренировку (85 % верных выборов); при этом на первых тренировках, несмотря на отсутствие подкрепления, тюлени в большинстве случаев (более 70 % выборов) выбирали контейнер с раствором камфоры (рис. 4).
□ Гренландский тюлень 1 □ Гренландский тюлень 2
Рис. 4. Доля верных выборов при дифференцировке контейнера с раствором ДМС от контейнера с раствором камфоры Fig. 4. The proportion of correct choices when differentiating a container with a solution of LCA from a container with a solution of camphor
При проведении контрольных испытаний все тюлени достоверно отличали контейнер с раствором ДМС от контейнера с раствором камфоры, в среднем совершая две-три ошибки за тренировку.
Заключение
Результаты проведенных исследований демонстрируют возможности тюленей двух видов вырабатывать двигательный условный рефлекс на обонятельные раздражители, а также их способности отличать один запах от другого. В процессе дифференцировки контейнера с запахом серые тюлени обучались быстрее гренландских тюленей, при дифференцировке двух запахов гренландские тюлени обучались на порядок медленнее. Схожие динамики обучения данных видов и результаты контрольных испытаний отмечались и при проведении исследований, касающихся выработки условных рефлексов на раздражители других модальностей [27]. Данное отличие в скорости обучения, вероятно, связано с видовыми особенностями исследуемых тюленей и свидетельствует о более высокой пластичности нервной деятельности серых тюленей, обитающих в условиях побережья, где информационная среда более насыщена, в отличие от гренландских тюленей, которые значительную часть жизни проводят либо в открытых водах, совершая сезонные миграции, либо вблизи паковых льдов, где редко встречаются с большими наборами запахов.
Список литературы Исследование обоняния серых и гренландских тюленей с помощью метода оперантного обучения
- Lowell W. R., Flanigan Jr W. F. Marine mammal chemoreception//Mammal review. 1980. V. 10, Iss. 1. P. 53-59. DOI: https://doi.org/10.1111/j.1365-2907.1980.tb00233.x.
- Brown R. E. The marine mammals: orders Cetacea, Pinnipedia, and Sirenia//Social odours in mammals/eds R. E. Brown, D. W. Macdonald. Oxford University Press, 1985. V. 2. P. 723-731.
- Thewissen J. G. M., George J., Rosa C., Kishida T. Olfaction and brain size in the bowhead whale (Balaena mysticetus)//Marine Mammals science, 2011. V. 27, Iss. 2. P. 282-294. j.1748-7692.2010.00406.x DOI: https://doi.org/10.1111/
- Feng P., Zheng J., Rossiter S. J., Wang D., Zhao H. Massive losses of taste receptor genes in toothed and baleen whales//Genome Biology and Evolution. 2014. V. 6, Iss. 6. P. 1254-1265. 10.1093/gbe/evu095 DOI: https://doi.org/
- Shusterman J. R. Behavioral capabilities of seals and sea lions: a review of their hearing, visual, learning and diving skills//The Psychological Record. 1981. V. 31, Iss. 2. P. 125-143. 10.1007/BF03394729 DOI: https://doi.org/
- Laska M., Svelander M., Amundin M. Successful acquisition of an olfactory discrimination paradigm by South African fur seals, Arctocephalus pusillus//Physiology & Behavior. 2008. V. 93, Iss. 4-5. P. 1033-1038.
- Ross G. J. Nuzzling behaviour in captive cape fur seals//International Zoo Yearbook. 1972. V. 12, Iss. 1. P. 183-184. DOI: https://doi.org/10.1111/j.1748-1090.1972.tb02321.x.
- Kovacs K. M. Maternal behaviour and early behavioural ontogeny of harp seals, Phoca groenlandica//Animal Behaviour. 1987. V. 35, Iss. 3. P. 844-855. DOI: https://doi.org/10.1016/S0003-3472(87)80120-3.
- Kovacs K. M. Mother-pup reunions in harp seals, Phoca groenlandica: cues for the relocation of pups//Canadian Journal of Zoology. 1995. V. 73, N 5. P. 843-849 DOI: https://doi.org/10.1139/z95-099
- Bartholomew G. A. Mother-young relations and the maturation of pup behavior in the Alaska fur seal//Animal Behaviour. 1959. V. 7, Iss. 3-4. P. 163-171. DOI: https://doi.org/10.1016/0003-3472(59)90005-3.
- Phillips A. V. Behavioral cues used in reunions between mother and pup South American fur seals (Arctocephalus australis)//Journal of Mammalogy. 2003. V. 84, Iss. 2. P. 524-535. 10.1644/1545-1542(2003)0842.0.CO;2 DOI: https://doi.org/
- Dobson F. S., Jouventin P. How mothers find their pups in a colony of Antarctic fur seals//Behavioural processes. 2003. V. 61, Iss.1-2. P. 77-85. DOI: https://doi.org/10.1016/S0376-6357(02)00164-X.
- Pitcher B. J., Harcourt R. G., Schaal B., Charrier I. Social olfaction in marine mammals: wild female Australian sea lions can identify their pup's scent//Biology Letters. 2011. V. 7. P. 60-62. URL: http://rsbl.royalsocietypublishing.org/content/roybiolett/7/1/60.full.pdf.
- Чечина О. Н. Видотипичное поведение сивучей (этологический атлас). Севастополь: Паритет-Информ, 2004. 120 c.
- Miller E. H. Social behaviour between adult male and female New Zealand fur seals, Arctocephalus forsteri (Lesson) during the breeding season//Australian Journal of Zoology. 1974. V. 22, N 2. P. 155-173 DOI: https://doi.org/10.1071/ZO9740155
- Белонович О. А., Рожнов В. В., Мамаев Е. Г., Бурканов В. Н. Обонятельные контакты сивучей (Eumetopias jubatus)//Морские млекопитающие Голарктики: сб. науч. тр. по материалам четвертой междунар. конф., Санкт-Петербург, Россия, 10-14 сент. 2006 г. Санкт-Петербург: Совет по морским млекопитающим, 2006. С. 64-67.
- Hardy M. H., Roff E., Smith T. G., Ryg M. Facial skin glands of ringed and grey seals, and their possible function as odoriferous organs//Canadian Journal of Zoology. 1991. V. 69, N 1. P. 189-200 DOI: https://doi.org/10.1139/z91-029
- Ryg M., Solberg Y., Lydersen C., Smith T. G. The scent of rutting male ringed seals (Phoca hispida)//Journal of Zoology. 1992. V. 226, Iss. 4. P. 681-689. DOI: https://doi.org/10.1111/j.1469-7998.1992.tb07509.x.
- Kiene R. P., Linne L. J., Bruton J. A. New and important roles for DMSP in marine microbial communities//Journal of Sea Research. 2000. V. 43, Iss. 3-4. P. 209-224. DOI: https://doi.org/10.1016/S1385-1101(00)00023-X.
- Nevitt G. A., Haberman K. Behavioral attraction of Leach's storm-petrels (Oceanodroma leucorhoa) to dimethyl sulfide//The Journal of Experimental Biology. 2003. V. 206, Iss. 9. P. 1497-1501. URL: http://jeb.biologists.org/content/jexbio/206/9/1497.full.pdf.
- Nevitt G. A., Losekoot M., Weimerskirch H. Evidence for olfactory search in wandering albatross, Diomedea exulans//Proceedings of the National Academy of Sciences. 2008. V. 105, Iss. 12. P. 4576-4581 DOI: https://doi.org/10.1073/pnas.0709047105
- Bonadonna F., Caro S., Jouventin P., Nevitt G. A. Evidence that blue petrel, Halobaena caerulea, fledglings can detect and orient to dimethyl sulfide//The Journal of Experimental Biology. 2006. V. 209, Iss. 11. P. 2165-2169. URL: http://jeb.biologists.org/content/jexbio/209/11/2165.full.pdf.
- Culik B. Finding food in the open ocean: foraging strategies in Humboldt penguins//Zoology. 2001. V. 104, Iss. 3-4. P. 327-338 DOI: https://doi.org/10.1078/0944-2006-00038
- Sims D. W., Quayle V. A. Selective foraging behaviour of basking sharks on zooplankton in a small-scale front//Nature. 1998. V. 393. P. 460-464 DOI: https://doi.org/10.1038/30959
- DeBose J. L., Lema S. C., Nevitt G. A. Dimethylsulfoniopropionate as a foraging cue for reef fishes//Science. 2008. V. 319, Iss. 5868. P. 1356 DOI: 10.1126/science.1151109
- Kowalewsky S., Dambach M., Mauck B., Dehnhardt G. High olfactory sensitivity for dimethyl sulphide in harbour seals//Biology Letters. 2006. V. 2. P. 106-109. DOI: http://dx.doi.org/10.1098/rsbl.2005.0380.
- Ишкулов Д. Г., Михайлюк А. Л., Пахомов М. В. Особенности цветовосприятия у серых тюленей//Вестник Кольского научного центра РАН. 2013. № 4 (15). С. 84-90.
