Исследование природы нервных волокон, вызывающих усиление сокращений двенадцатиперстной кишки при раздражении симпатического нерва

Автор: Свешников Дмитрий Сергеевич, Киричук Вячеслав Федорович, Смирнов Виктор Михайлович, Мясников Игорь Леонидович, Кучук Андрей Владимирович, Трофимова Юлия Евгеньевна, Трубецкая Мария Алексеевна
Журнал: Саратовский научно-медицинский журнал @ssmj
Рубрика: Физиология и патофизиология
Статья в выпуске: 3 т.7, 2011 года.
Бесплатный доступ
Цель работы: исследовать механизм усиления сокращений двенадцатиперстной кишки при раздражении симпатического нерва. Материал и методы. В экспериментах на собаках показали, что стимуляторные реакции, вызванные электрическим раздражением симпатического нерва на фоне совместной блокады а-адренорецепторов фентоламином и (3-адренорецепторов пропранололом практически полностью устраняются лизерголом — блокатором 5-НТ12-рецепторов. Результаты. Введение лизергола не оказывало существенного влияния как на фоновую моторную активность двенадцатиперстной кишки, так и на функцию блуждающего нерва. Сделали вывод о том, что эффекторный нейрон, с помощью которого реализуется стимуляторный феномен, является серотонинергическим, его влияния осуществляются с помощью 5-НТ12-рецепторов
Двенадцатиперстная кишка, регуляция, серотонинорецепторы
Короткий адрес: https://sciup.org/14917362
IDR: 14917362
Текст научной статьи Исследование природы нервных волокон, вызывающих усиление сокращений двенадцатиперстной кишки при раздражении симпатического нерва
Адрес: 117997, г. Москва, ул. Островитянова, 1.
Тел.: (495) 434–2511, 89032750166.
Целью настоящей работы было исследовать механизм усиления сокращений двенадцатиперстной кишки при раздражении симпатического нерва.
Методы. Острые опыты выполнили на 7 собаках обоего пола, массой 10–15 кг в условиях хирургической стадии этаминалового наркоза (нембутал 60 мг/кг, внутримышечно), при вскрытой грудной клетке и искусственной вентиляции легких. Опыты проводили в соответствии со стандартами этического комитета. Наркоз был достаточным, в ходе опыта животные не испытывали каких-либо неприятных ощущений. В опытах производили двустороннюю ваготомию на шее для исключения рефлекторных влияний. Методической особенностью нашего подхода являлось то, что, в отличие от других исследователей [5], мы впервые раздражали симпатический нерв в грудной полости, где он не содержит примесь парасимпатических волокон блуждающего нерва (грудную клетку вскрывали в IX межреберье справа, симпатический ствол пересекали на уровне VI грудного позвонка). Периферический отрезок симпатического нерва с помощью лигатуры помещали на биполярные раздражающие электроды с межэлектродным расстоянием 2 мм, нерв раздражали в течение 30 с прямоугольными электрическими импульсами, длительностью 1,5 мс различной интенсивности: 1-3-5-10-15 В, 10 и 20 Гц. Симпатический отдел вегетативной нервной системы блокировали фентоламином (2 мг/кг) — блокатором α-адренорецепторов и пропранололом (4 мг/кг) — блокатором в-адренорецепторов. Фар-
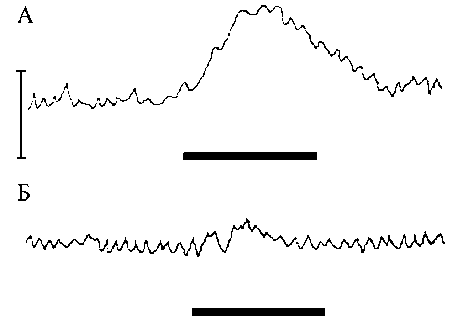
Рис. 1. Реакции двенадцатиперстной кишки на раздражение симпатического ствола (10 в, 20 Гц) в различных условиях: А – на фоне действия α- и β-блокаторов фентоламина и пропранолола, Б – на фоне совместного действия адреноблокаторов и лизергола – блокатора 5-HT1,2 – рецепторов. На каждом фрагменте запись давления в полости кишки и нулевая линия, она же (более жирная часть линии) отметка раздражения – 30 с. Шкала 10 мм рт. ст. для обоих фрагментов макологическую блокаду 5-HT12 — рецепторов осуществляли лизерголом (1 мг/кг), препарат вводили на фоне предварительного совместного действия фентоламина и пропранолола. Все блокаторы вводили внутривенно.
При статистическом анализе использовали проверку нормальности распределения количественных признаков по критерию Колмогорова — Смирнова, степень достоверности оценивали с помощью Т-теста Стьюдента в зависимой и независимой выборках. Обработка данных в зависимой выборке предусматривала анализ типа «до и после воздействия», а в независимой выборке обрабатывались данные прироста давления у интактных животных и на фоне блокаторов различных рецепторов. Критическое значение уровня значимости принималось равным 5% (р < 0,05).
Результаты. Эксперименты с лизерголом, блокатором 5-HT12 -рецепторов поставили на 7 собаках и обнаружили,, что усиление сокращений двенадцатиперстной кишки, вызванное раздражением симпатического нерва, практически полностью устраняется лизерголом (1мг/кг). На фоне лизергола количество стимуляторных ответов двенадцатиперстной кишки на раздражения нерва уменьшилось более чем в 3 раза, а степень выраженности оставшихся стимуля-торных реакций была незначительной. Так, при раздражении 5В, 10 Гц только у одного животного (14%) зарегистрирован стимуляторный ответ. Однако при более сильных импульсах 10 В и 15 В эффект сохранился у трех животных (43%), у четырех (59%) стимуляторный феномен был полностью выключен. Кроме того, сохранившиеся реакции были выражены слабо: гидростатическое давление в ответ на раздражение нерва 10 В 10 Гц в трех пробах повысилось всего лишь с 11,6±4,2 до 12,9±3,92 мм рт. ст. (11%, Р >0,05); при стимуле 15 В 10 Гц внутриполостное давление повышалось с 12,0±5,3 до 16±6 мм рт. ст., что также недостоверно (рис. 1). Более подробно частота встречаемости стимуляторных ответов (в абсолютных числах и процентах) представлена в табл. 1
Таблица 1
Частота встречаемости стимуляторных эффектов при раздражении симпатического ствола в грудной полости у собак в различных условиях (в каждой графе указано число животных с однотипными реакциями и их процент)
|
Условия опытов |
Амплитуда раздражающих импульсов частотой 10Гц |
||||
|
1В |
3В |
5В |
10В |
15В |
|
|
На фоне действия α-и β- адреноблокаторов |
0 0% |
3 42,9% |
5 71,4% |
7 100% |
7 100% |
|
На фоне совместного действия адреноблокаторов и лизергола — блокатора 5-HT1,2- рецепторов |
0 0% |
0 0% |
1 14,85% |
3 42,85% |
3 42,85% |
При этом введение лизергола не оказывало существенного влияния на фоновую моторную активность двенадцатиперстной кишки, и, следовательно, результаты фармакологического анализа являются
Содержание серотонина и катехоламинов в нервах крыс
Обсуждение. Таким образом, результаты наших опытов с лизерголом свидетельствуют о том, что эффекторный нейрон, с помощью которого реализуется стимуляторный феномен, является серотонинергическим, его влияния осуществляются с помощью 5-HT12 -рецепторов гладких мышц. Наше заключение о сер, отонинергическом механизме стимуляторного эффекта подтверждается также полученными ранее результатами о наличии серотонина в симпатическом стволе. В экспериментах использовались биохимическая методика Оксенкруга и гистохимическая Фалька — Хилларпа в модификации Крохиной [5, 6].
В частности, с помощью гистохимической методики Фалька — Хилларпа в модификации Крохиной в препаратах правого и левого больших чревных нервов (в сравнении с препаратами блуждающих и соматических нервов) В. М. Смирновым и С. Ф. Волынцевой [6] было обнаружено яркое свечение адренергических и серотонинергических структур. В чревных нервах на светло-сером фоне наблюдаются ярко светящиеся тонкие волокна. Узлы симпатической цепочки излучают очень яркое свечение в голубовато-желтой области спектра. Флуоресцентным методом выявлено наличие там большого количества серотонина. Препараты блуждающего нерва и соматических нервов светятся значительно слабее, чем симпатических стволов и ганглиев. Содержание серотонина и катехоламинов в нервных структурах представлены в табл. 2. Эти данные также свидетельствуют о том, что в составе симпатического ствола и большого чревного нерва имеются серотонинергические нервные волокна.
Заключение. Наше заключение о серотонинерги-чеcком механизме реализации стимуляторного феномена подтверждают многочисленные морфологические исследования, в которых установлена обширность распространения серотонинреактивных структур в энтеральной нервной системе. Выявлены также системы связывания серотонина в энтеральных симпатических окончаниях [7]. В симпатическом отделе спинного мозга обнаружены серотониновые рецепторы, где, по-видимому, локализованы центры интеграции серотонинреактивных структур [8].
Иммуноцитохимическими методиками идентифицированы серотонинсодержащие нейроны, главным образом располагающиеся в ауэрбаховом сплетении тонкой кишки человека [9, 10]. Серотонинергические нейроны и их волокна расположены преимущественно в межмышечном нервном сплетении, выявлены связи данного типа волокон с постганглионарными нейронами. Серотонин-иммунореактивные волокна образуют сплетения, прослеживаемые до циркулярного мышечного слоя. Встречаются они и в подслизистом слое. Кроме того, в тонкой кишке морской свинки обнаружены системы связывания серотонина — моноаминоксидаза подтипа А (МАО-А); МАО-А содержится в симпатических варикозных аксонах [9]. В миентеральном сплетении установлена иммунореактивность серотонина в нейронах, имеющих морфологические характеристики клеток I типа Догеля. Один длинный отросток этих клеток, который, вероятно, является аксоном, простирается аборально на значительные расстояния. Некоторые из этих отростков иннервируют нейроны не только миентерально-го, но и подслизистого сплетения.
Электрофизиологическими методиками зарегистрированы в нейронах ЭНС медленные ВПСП, вызванные серотонином, эти ВПСП обеспечивают длительные серии спайковых разрядов и составлют основу моторной активности [7, 10, 11].
Таким образом, наше заключение о серотонинергической природе нервных волокон симпатического ствола, усиливающих сокращение двенадцатиперстной кишки, косвенно подтверждается морфологическими и электрофизиологическими исследованиями.
Список литературы Исследование природы нервных волокон, вызывающих усиление сокращений двенадцатиперстной кишки при раздражении симпатического нерва
- Удельное М.Г. Нервная регуляция сердца. М.: Изд-во МГУ, 1961.380 с.
- Смирнов В.М., Свешников Д. С. Берсенева Е.А. Возможная роль серотонинреактивных структур в усилении двигательной активности двенадцатиперстной кишки, вызванной раздражением симпатического ствола//Авиакосмическая и экологическая медицина. 1999. Т. 33, № 5. С. 40-45.
- Delbro D., Fandriks L, Lisander В. Hexametonium-resistant, atropin-sensitiv vagal excitation of the feline stomach -activation of an unknown fibre system//Acta Physiol Scand. 1981. Vol. 112, № 4. P. 493-494.
- Fandriks L, Delbro D. Cholinergic excitatory motor responses of the feline colon elicited by high-strength sympathetic nerve//Acta Physiol Scand. 1983. Vol. 117, № 3. P. 473-475.
- Смирнов B.M., Клевцов В.А., Смирнова H.A., Лычко-ва А. Э. Механизм стимуляции сокращений желудка и двенадцатиперстной кишки большим чревным нервом//Физиол. журн. СССР. 1986. Т. 72, № 5. С. 650-655.
- В.М. Смирнов, С.Ф. Волынцева. Серотонинергическая природа нервных волокон, усиливающих сокращения желудка и кишечника//Структура и функции вегетативной нервной системы. Воронеж, 1995. С. 95-97.
- Gershon М. Enteric serotonergic neurones... finally!//J. Physiol. 2009. Vol. 587, part 3. P. 507-508.
- Jansen A.S., Hoffman J. L, Loewy A.D. CNS sites involved in sympathetic and parasympathetic control of the pancreas: a viral tracing study//Brain Res. 1997. Vol. 766, № 1-2. P. 29-38.
- Gershon M. The Second Brain: The Scientific Basis of Gut Instinct and a roundbreaking New Understanding of Nervous Disorders of the Stomach and Intestine//Harper Collins Publishers. 1999. P. 1022-1023.
- Talley N. Serotoninergic neuroenteric modulators//Lancet. 2001. Vol. 358. P. 2061-2068.
- Wood J. Enteric nervous system: reflexes, pattern generators and motility//Curr Opin Gastroenterol. 2008. Vol. 24, №2. P. 149-158.
