Исследование содержания свободного пролина в растениях кукурузы, трансформированных in planta с использованием LBA4404, несущего PBI2E с двухцепочечным рнк-супрессором гена пролиндегидрогеназы

Автор: Михальская Светлана Ивановна, Матвеева Александра Юрьевна, Сергеева Лариса Евгеньевна, Кочетов Алексей Владимирович, Тищенко Елена Николаевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Получены трансгенные Т1-растения кукурузы ( Zea mays L.), содержащие двухцепочечный РНК-супрессор гена пролиндегидрогеназы. Проанализирован уровень свободного L-пролина в побегах и корнях проростков в условиях сульфатно-хлоридного засоления. Показано, что стресс-устойчивость сопровождается повышением содержания этого осмолита.
Солеустойчивость, супрессор гена пролиндегидрогеназы, трансгенез
Короткий адрес: https://sciup.org/148205659
IDR: 148205659 | УДК: 579.254.2
Investigation of the free proline contents in corn plants transformated in planta using LBA4404, harboring PBI2E with double-stranded rna-suppressor proline dehydrogenase
The transgenic corn Т1-plants ( Zea mays L.) with double-stranded RNA-suppressor proline dehydrogenase were ob- tained. The levels of free L-proline in shoots and roots under sulfate-chloride salinity were estimated. There was shown that corn salt tolerance was conferred by elevated level of this osmolyte.
Текст научной статьи Исследование содержания свободного пролина в растениях кукурузы, трансформированных in planta с использованием LBA4404, несущего PBI2E с двухцепочечным рнк-супрессором гена пролиндегидрогеназы
Метаболическая инженерия предоставила новые возможности для решения проблемы устойчивости культурных растений к осмотическим стрессам. Одно из перспективных её направлений связано с генами, контролирующими метаболизм L-пролина (Pro). Генетические и физиолого-биохимические исследования показали, что L-пролин – полифунк-циональная аминокислота, которая может принимать участие в сложных интегральных процессах адаптации растений. Развивается также представление о Pro как о сигнальной/регуляторной молекуле в ходе роста, дифференцировки клеток и их программированной гибели [1-6]. Повышение уровня свободного L-пролина - физиологическая реакция многих видов растений в ответ на разные типы абиотических стрессов. Вместе с тем, роль Pro и его участие в процессах адапта-ции/устойчивости не всегда очевидна, о чём свидетельствуют результаты исследований уровня толерантности трансгенных растений и диких видов растений [2, 7-9].
Эндогенный уровень L-пролина растений в норме и при стрессе координировано регулируется его синтезом, катаболизмом и транспортом [10, 1, 2]. Гены, контролирующие метаболизм Pro, кодируются ядерным геномом, и наиболее детально исследованы у арабидопсиса и табака. Проанализированные на сегодняшний день гены синтеза - P5CS (энзим ∆1-пирролин-5-карбоксилатсинтетаза, ЕС.2.7.2.11.1.2.1.41) и катаболизма - ProDH (энзим пролиндегидрогеназа, ЕС.1.5.99.8.) ряда растений находятся в центре внимания при разработке молекулярных биотехнологий, направленных на повышение уровня стрессоустойчивости культурных
растений . Они представлены единичными копиями с высоким уровнем гомологии, каждая из которых может выполнять различные функции, связанные с ответной реакцией на стресс и/или с пролиферацией, дифференцировкой клеток растений [11, 1, 2]. В связи с чем целесообразен предварительный анализ эффективности их применения для получения биотехнологических продуктов.
Что касается ProDH , то для повышения уровня свободного пролина исследуются возможности частичной супрессии генов, контролирующих катаболизм L-пролина, с использованием векторных конструкций, создаваемых на основе чужеродных генов пролиндегидрогеназы, расположенных в антисмысловой ориентации или в форме обращённого повтора. При этом есть основания полагать, что применение siRNA –технологий является более эффективным [5, 6, 12, 13].
Целью данной работы был анализ содержания L-пролина в проростках кукурузы, трансформированной in planta с использованием обезоруженного агробактериального штамма LBA 4404, содержащего плазмиду с двухцепочечным (дц)РНК-супрессором гена пролиндегидрогеназы, в условиях стресса, вызванного сульфатно-хлоридным засолением.
Объектом исследования служила инбредная линия кукурузы 250 (селекции Института физиологии растений и генетики НАН Украины), трансформированная in planta с использованием обезоруженного агробактериального штамма LBA 4404, несущего бинарный вектор pBi2E с дцРНК-супрессором гена пролиндегидрогеназы, полученным на основе гена арабидопсиса ProDH1 (рис. 1).
Присутствие трансгена в геноме кукурузы анализировали ПЦР-методом так, как описано ранее [14].
RB LB
|
NOSpro |
npt II |
NOSterm |
35Spro |
ProDH-ex1 |
int1 |
ProDH-ex1 |
NOSterm |
Рис. 1. Блок-схема Т-ДНК области pBi2E. NOSpro и 35Spro – соответственно промоторы гена нопалинсинтазы и 35S вируса мозаики цветной капусты; ProDH-ex1 и int1 - фрагменты первого экзона и интрона гена ProDH1 арабидопсиса, соответственно; NOSterm – сигнал полиаденилирования гена нопалинсинтазы; npt II – ген неомицинфосфотрансферазы E. coli, RB, LB – повторы, ограничивающие Т-область
Уровень свободного L-пролина (Pro) определяли методом Чинарда с модификациями [15]. Для сравнительного анализа использовали 14-, 18- и 21-суточные проростки. Пробы отбирали и фиксировали в одно и то же время суток. Стрессовые условия создавали добавлением в среду культивирования летальные дозы солей морской воды (2,0% и 2,5%). Стрессовому воздействию подвергали 7-суточные проростки, выращенные в нормальных условиях (1 вариант), и зерновки, проращиваемые и культивируемые в стрессовых условиях в течение 28 дней (2 вариант). Содержания Pro определяли в корнях и побегах 18-суточных трансгенных проростков, полученных при проращивании зерновок (по 8 штук), взятых из 6-ти разных початков. Контролем служили побеги или корни нетрансгенных вариантов в нормальных/стрессовых условиях. При статистической обработке использовали критерий Стьюдента.
Для углубления представления о роли генов ProDH в процессах адаптации и жизнедеятельности кукурузы исследовали содержание свободного L-пролина в проростках кукурузы, трансформированной in planta с применением LBA4404, несущего плазмиду с дцРНК-супрессором гена пролиндегидрогеназы, в условиях засоления, приводившего к гибели контрольных (нетрансгенных) вариантов. Параллельно, такой подход позволяет проанализировать эффективность частичной супрессии генов ProDH кукурузы с использованием siRNA-технологий.
На рис. 2 представлены результаты содержания свободного Pro в побегах проростков, адаптированных к засолению (общий возраст проростков 14 суток, вариант 1). Видно, что уровень этой аминокислоты у ряда индивидуальных трансгенных вариантов варьировал, однако по абсолютной величине, за исключением трёх случаев, существенно превышал содержание Pro в проростках контрольных побегов. По-видимому, в вариантах V, V1 и X1 трансген был нефункциональным или уровень его транскрипции был незначительным. Полученные данные дают основание предполагать, что эта аминокислота выступает в качестве протектора при адаптации к ионному стрессу. Отметим, что повышение уровня свободного пролина у трансгенных растений кукурузы, содержащих антисмысловой супрессор гена пролиндегидрогеназы, на фоне хло-ридного засоления ранее показано Моисеевой Е.М. и соавт. [15].
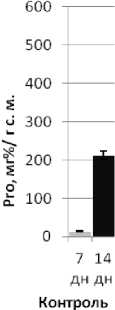
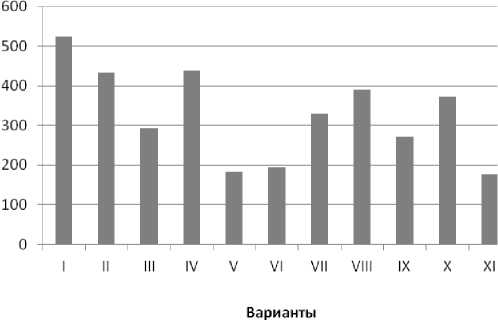
Рис. 2 . Содержание свободного L-пролина в побегах адаптировавшихся к сульфатно-хлоридному засолению (2,0% солей морской воды) 14-суточных проростков кукурузы (1 – Х1). Контроль: 7 сут в нормальных условиях и через 7 сут в условиях стресса
Зерновки трансгенных растений кукурузы характеризовались также способностью к прорастанию в условиях постоянного действия стрессового фактора (вариант 2). Их проращивали сначала на фоне 2,0% засоления (летальная доза стрессора), затем 7-суточные проростки переносили в более жёсткие условия стресса (2,5% солей морской во- ды) и культивировали ещё 2 недели. Рис. 3 отображает содержание свободного пролина в 21-суточных проростках. Видно, что в условиях стресса уровень Pro существенно превышал показатель контроля. Это свидетельствует в пользу того, что этот осмолит является одним из факторов, принимающих участие в поддержании жизнедеятельно-

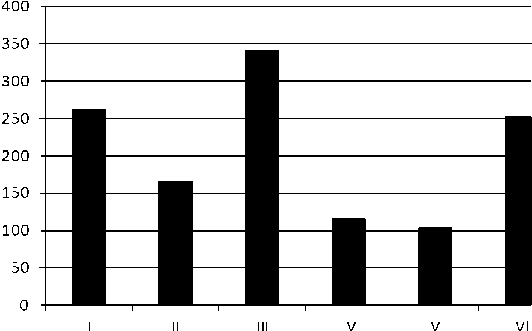
Рис. 3 . Содержание свободного L-пролина в побегах 21-суточных проростков, культивируемых в условиях возрастающего сульфатно-хлоридного засоления (последовательно 2,0% и 2.5% солей морской воды). К – побеги нетрансгенных 7-суточных проростков в условиях стресса (2,0% солей морской воды)
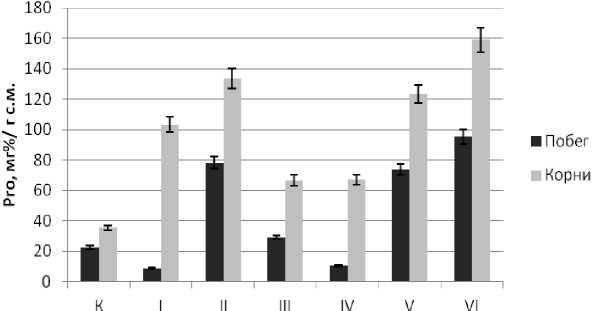
Рис. 4 . Содержание свободного L-пролина в корнях и побегах 18-суточных трансгенных проростков, которые культивировались в условиях постоянного действия сульфатно-хлоридного засоления (2,5% солей морской воды). К – побеги трансгенных проростков в отсутствие стресса
сти кукурузы в стрессовых условиях. Очевидно, что уровень свободного пролина является результатом координированного функционирования системы синтеза – катаболизма.
Сравнительное изучение содержания свободного Pro в побегах и в корнях 18-суточных трансгенных проростков показало, что уровень его накопления в корнях превышал этот показатель в побегах, как в норме, так и в условиях стресса (рис. 4). Наблюдаемые различия в нормальных условиях культивирования трансгенных проростков свидетельствуют о тканеспецифичности аккумуляции этой аминокислоты. Следует отметить, что в контрольных вариантах (дикий тип) уже на 14-й день опыта содержание пролина составляло в побегах (138,66 ± 30,41 мг%/г с.м.), корнях (146,63 ± 19,86 мг%/г с.м.) и достоверно не различалось. Далее продолжалась прогрессирующая гибель контроля. Высокие показатели уровня пролина в данном случае, очевидно, являлись отражением деградации пролин-содержащих компартментов клеток. Эти результаты согласуются с литературными данными [16, 17]. При летальных дозах сульфатно-хлоридного засоления у трансформантов по сравнению с контролем (трансгенные проростки) про- исходило существенное увеличение содержания свободного пролина. Больший уровень аккумуляции этой аминокислоты в корнях, возможно, являлся отражением депонирования токсичных ионов.
Таким образом, солеустойчивость трансформантов, содержащих двухцепочечный РНК-супрессор гена пролиндегидрогеназы кукурузы, сопровождался аккумуляцией свободного L-пролина.
Работа поддержана грантом совместных научных проектов НАН Украины (16-05-2012) - СО РАН (№11)
-
1. Kavi Kishor P.B., Sangam S., Amrutha R.N. et al. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implication in plant growth and abiotic stress tolerance // Current Sci. 2005. V. 88. N 3. P. 424438.
-
2. Szabados L, Savoure A . Proline: a multifunctional amino acid // Trends in plant science. 2009. V.15. N 2. P. 89- 97.
-
3. Maggio A., Miyazaki S., Veronese P. et al. Does proline accumulation play an active role in stress-induced growth reduction? // The Plant Journal. 2002. V. 3. N 16. P. 699-712.
-
4. Cecchini NM, Monteoliva MI, Alvarez ME. Proline dehydrogenase is a positive regulator of cell death in different kingdoms // Plant Signal Behav. 2011. V. 6. N 8. P. 11951197.
-
5. Кочетов А.В., Титов С.Е., Колодяжная Я.С. и др . Повышение содержания пролина и осмотического давле- ния клеточного сока у трансформантов табака, несущих антисмысловой супрессор гена пролиндегидрогеназы // Генетика. 2004. Т. 40. № 2. С. 282-285.
-
6. Ибрагимова С.С., Колодяжная Я.С., Герасимова С.В., Кочетов А.В. Частичная супрессия гена пролиндегидрогеназы увеличивает устойчивость растений к различным видам абиотических стрессов // Физиология растений. 2012. Т. 59. С. 99-107.
-
7. Mani S., Van de Cotte B, Van Montagu M., Verbruggen N . Altered levels of proline dehydrogenase cause hypersensitivity to proline and its analogs in Arabidopsis // Plant Physiology. 2002. V. 128. P. 73-83.
-
8. Kaplan F, Kopka J, Sung DY et al. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content // Plant J. 2007. V. 50. P. 967–981.
-
9. Sharma S. Villamor J. G., P. E. Verslues. Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential // Plant Physiology. 2011. V. 157. P. 292-304.
-
10. Peng Z., Lu Q., Verna D.P. Reciprocal regulation of delta 1-pirroline-5-carboxylate syntetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants // Mol. Gen. Genet. 1996. N 3. P. 334-341.
-
11. Miller G., Stein H., Honig A. et al. Responsive modes of Medicago sativa proline dehydrogenase genes during salt stress and recovery dictate proline accumulation // Planta. 2005. V. 222/ N 1. P. 70-79.
-
12. Титов С.Е. Изучение генетически модифицированных растений табака ( Nicotiana tabacum L.), экспрессирую-
щих антисмысловой супрессор гена пролинденидроге-назы: Автореф. дисс. …канд. биол. наук. Новосибирск, 2008. 16 с.
-
13. Tateishi Y., Nakagama T., Esaka M. Osmotolerance and growth stimulation of transgenic tobacco cells accumulating free proline by dehydrogenase expression with doublestranded RNA interference technique // Physiologia Plantarum. 2005. 125. P.1399-3054.
-
14. Михальская С.И., Адаменко Н.И., Моргун Б.В . и др . Компетентность к Agrobacterium –опосредованной трансформации сегментов побега элитных инбредных линий кукурузы // Біотехнологія. 2012. Т. 5. № 3. С. 98-103.
-
15. Сергеева Л.Е., КомисаренкоА.Г.,Бронникова Л.И. и др. Содержание свободного пролина в тканях подсолнечника при реализации морфогенетического потенциала in vitro // Біотехнологія. 2013. Т. 6. № 1. С. 113-118.
-
16. Моисеева Е.М., Агапонов Д.А., Вешков В.А. и др . Повышение содержания пролина в растениях кукурузы, экспрессирующих фрагмент гена пролиндегидрогеназы в антисмысловой ориентации // Физиология растений. 2012. Т. 59. № 3. С. 457-460.
-
17. Михальская С.И., Адаменко Н.И., Моргун Б.В . и др . Компетентность к Agrobacterium – опосредованной трансформации сегментов побега элитных инбредных линий кукурузы // Біотехнологія. 2012. Т. 5. № 3. С. 98103.
-
18. Battaglia M. , Solorzanlo R.M., Hernandez P. et al. Prolinerich cell wall proteins accumulate in growing regions and phloem tissue in response to water deficit in common bean seedlings // Planta. 2007. V. 225. P. 1121-1133.
-
19. Goo J.N., Park A.R., Park W.J., Park O.K. Selection of Ara-bidopsis genes encoding and secreted plasma membrane proteins // Plant Mol. Biol. 1999. V. 41. P. 415-423.
INVESTIGATION OF THE FREE PROLINE CONTENTS IN CORN PLANTS TRANSFORMATED IN PLANTA USING LBA4404, HARBORING pBi2E WITH DOUBLE-STRANDED RNA-SUPPRESSOR PROLINE DEHYDROGENASE
1 Institute of Plant Physiology and Genetics of National Academy of Science of Ukraine, Kyiv 2 Institute of Cytology and Genetics, Russian Academy of Science, Novosibirsk
The transgenic corn Т1-plants ( Zea mays L.) with double-stranded RNA-suppressor proline dehydrogenase were obtained. The levels of free L-proline in shoots and roots under sulfate-chloride salinity were estimated. There was shown that corn salt tolerance was conferred by elevated level of this osmolyte.
Список литературы Исследование содержания свободного пролина в растениях кукурузы, трансформированных in planta с использованием LBA4404, несущего PBI2E с двухцепочечным рнк-супрессором гена пролиндегидрогеназы
- Kavi Kishor P.B., Sangam S., Amrutha R.N. et al. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implication in plant growth and abiotic stress tolerance//Current Sci. 2005. V. 88. N 3. P. 424-438.
- Szabados L, Savoure A. Proline: a multifunctional amino acid//Trends in plant science. 2009. V.15. N 2. P. 89-97.
- Maggio A., Miyazaki S., Veronese P. et al. Does proline accumulation play an active role in stress-induced growth reduction?//The Plant Journal. 2002. V. 3. N 16. P. 699-712.
- Cecchini NM, Monteoliva MI, Alvarez ME. Proline dehydrogenase is a positive regulator of cell death in different kingdoms//Plant Signal Behav. 2011. V. 6. N 8. P. 1195-1197.
- Кочетов А.В., Титов С.Е., Колодяжная Я.С. и др. Повышение содержания пролина и осмотического давления клеточного сока у трансформантов табака, несущих антисмысловой супрессор гена пролиндегидрогеназы//Генетика. 2004. Т. 40. № 2. С. 282-285.
- Ибрагимова С.С., Колодяжная Я.С., Герасимова С.В., Кочетов А.В. Частичная супрессия гена пролиндегидрогеназы увеличивает устойчивость растений к различным видам абиотических стрессов//Физиология растений. 2012. Т. 59. С. 99-107.
- Mani S., Van de Cotte B, Van Montagu M., Verbruggen N. Altered levels of proline dehydrogenase cause hypersensitivity to proline and its analogs in Arabidopsis//Plant Physiology. 2002. V. 128. P. 73-83.
- Kaplan F, Kopka J, Sung DY et al. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content//Plant J. 2007. V. 50. P. 967-981.
- Sharma S. Villamor J. G., P. E. Verslues. Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential//Plant Physiology. 2011. V. 157. P. 292-304.
- Peng Z., Lu Q., Verna D.P. Reciprocal regulation of delta 1-pirroline-5-carboxylate syntetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants//Mol. Gen. Genet. 1996. N 3. P. 334-341.
- Miller G., Stein H., Honig A. et al. Responsive modes of Medicago sativa proline dehydrogenase genes during salt stress and recovery dictate proline accumulation//Planta. 2005. V. 222/N 1. P. 70-79.
- Титов С.Е. Изучение генетически модифицированных растений табака (Nicotiana tabacum L.), экспрессирующих антисмысловой супрессор гена пролинденидрогеназы: Автореф. дисс. …канд. биол. наук. Новосибирск, 2008. 16 с.
- Tateishi Y., Nakagama T., Esaka M. Osmotolerance and growth stimulation of transgenic tobacco cells accumulating free proline by dehydrogenase expression with doublestranded RNA interference technique//Physiologia Plantarum. 2005. 125. P.1399-3054.
- Михальская С.И., Адаменко Н.И., Моргун Б.В. и др. Компетентность к Agrobacterium-опосредованной трансформации сегментов побега элитных инбредных линий кукурузы//Бiотехнологiя. 2012. Т. 5. № 3. С. 98-103.
- Сергеева Л.Е., Комисаренко А.Г.,Бронникова Л.И. и др. Содержание свободного пролина в тканях подсолнечника при реализации морфогенетического потенциала in vitro//Бiотехнологiя. 2013. Т. 6. № 1. С. 113-118.
- Моисеева Е.М., Агапонов Д.А., Вешков В.А. и др. Повышение содержания пролина в растениях кукурузы, экспрессирующих фрагмент гена пролиндегидрогеназы в антисмысловой ориентации//Физиология растений. 2012. Т. 59. № 3. С. 457-460.
- Михальская С.И., Адаменко Н.И., Моргун Б.В. и др. Компетентность к Agrobacterium -опосредованной трансформации сегментов побега элитных инбредных линий кукурузы//Бiотехнологiя. 2012. Т. 5. № 3. С. 98-103.
- Battaglia M., Solorzanlo R.M., Hernandez P. et al. Prolinerich cell wall proteins accumulate in growing regions and phloem tissue in response to water deficit in common bean seedlings//Planta. 2007. V. 225. P. 1121-1133.
- Goo J.N., Park A.R., Park W.J., Park O.K. Selection of Arabidopsis genes encoding and secreted plasma membrane proteins//Plant Mol. Biol. 1999. V. 41. P. 415-423.
