Исследование влияния АзоТАБа на фототаксис Planaria Torva

Автор: Барсков Кирилл Владиславович, Ерофеев Иван Станиславович, Агладзе Константин Игоревич
Журнал: Труды Московского физико-технического института @trudy-mipt
Рубрика: Бионанофизика
Статья в выпуске: 1 (17) т.5, 2013 года.
Бесплатный доступ
Работа посвящена изучению действия производного азобензола с фотоконтролируемой структурой (АзоТАБа) на движение плоских червей вида Planaria Torva. В настоящее время известно, что транс-АзоТАБ является эффективным ингибитором ионных каналов мембраны возбудимых клеток, нейронов и кардиомиоцитов. В то же время цис-АзоТАБ подобной активности не проявляет. Представлялось интересным исследовать, как влияет присутствие изомеров АзоТАБа на простейшее поведение многоклеточных организмов, обладающих зачатками дифференцированной нервной системы, таких как планарии. Были исследованы траектории движения планарий при освещении видимым и УФ-светом как в среде, содержащей АзоТАБ в различных концентрациях, так и в его отсутствии. Кроме того, были измерены скорости передвижения планарий в воде в свободном движении, при фототаксисе и в случае фототаксиса в присуствии АзоТАБа. Также показана обратимость блокирования двигательных функций планарий под действием УФ в присутствии АзоТАБа.
Азотаб, фотоконтроль, фототаксис
Короткий адрес: https://sciup.org/142185902
IDR: 142185902
Effect of AzoTAB on phototaxis in Planaria Torva
This paper is devoted to the effect of an azobenzol derivative with photocontrolable structure (AzoTAB) on the motion of flatworms Planariatorva. Trans-AzoTAB efficiently inhibits ion channels in excitable cell membrane, neurons and cardiomyocytes, while cis- AzoTAB does not. In the current research we study how the presence of AzoTAB isomers controls the simplest behaviour of pluricellular organisms with the rudiments of differentiated nervous system like planaria. We also study the trajectories of planaria which are illuminated with visible and UV light in the medium with AzoTAB of various concentrations. In addition, we measure the velocities of planaria positive and negative phototaxis in the media containing AzoTAB isomers. We demonstrate that the tinhibitory effect of the AzoTAB on planaria motion under the UV light is reversible.
Текст научной статьи Исследование влияния АзоТАБа на фототаксис Planaria Torva
В последние несколько лет было обнаружено, что, используя вещества, на. основе азо-бензена, можно обратимо подавлять активность ионных каналов возбудимых клеток с помощью света. [1, 7, 9]. Наиболее эффективным для этой цели оказался бромида, азобен-зен триметиламмония (АзоТАБ), который под действием света подвергается цис-транс изомеризации. При этом транс-изомер АзоТАБа. блокирует возбудимые клетки, такие как кардиомиоциты, мышечные клетки и нейроны [7]. В данной работе исследовалось влияние АзоТАБа. в целом на. организм, и в качестве экспериментальной модели были выбраны пла-нарии. Планарии — семейство плоских червей из подотряда. Tricladida класса, ресничных червей. Планарии удобны для работы во многих отношениях [2-6,10]. С одной стороны, они достаточно примитивны и неприхотливы, с другой стороны — планарии имеют зачатки мозга, и нервной системы, а. их тело представляет собой нервно-мышечный каркас. Таким образом, планария представляет собой удобный объект для изучения влияния различных веществ на. возбудимые клетки. Планарии избегают ультрафиолетового излучения, так называемый отрицательный фототаксис. В растворе АзоТАБа. с большой концентрацией транс-изомера двигательная способность организма, падает. При облучении ультрафиолетом АзоТАБ переходит в цис-форму, при этом двигательная функция планарии восстанавливается, и червь старается покинуть зону облучения. Целью данной работы было выявить эти эффекты, исследовать фототаксис планарии при различных интенсивностях ультрафиолета, исследовать влияние АзоТАБа. на. фототаксис, таким образом, описав действие вещества на организм. Необходимо было набрать статистику поведения плаиарий в различных условиях и описать эффекты количественно в таких параметрах, как скорость, траектория и время нахождения в том или ином участке планариума.
2. Материалы и методы2.1. Содержание планарий
Планарии содержались в пресной воде при постоянной температуре 20 °C. Смена воды осуществлялась раз в неделю. Питание проводилось 2-3 раза в неделю свежим мясом или свежей печенью. В экспериментах использовались особи длиной более 5 мм.
2.2. Приготовление раствора АзоТАБа
АзоТАБ был синтезирован ЦВТ «ХимРар». Для работы были приготовлены водные растворы АзоТАБа концентраций 50 мкМоль/л, 100 мкМоль/л и 200 мкМоль/л. Сначала делался стоковый раствор концентрацией 10 мМоль/л, из которого путем разбавления в соответствующей пропорции получались требуемые растворы.
2.3. Измерения
Для проведения измерений было изготовлено два «планариума». Первый представлял собой ванночку из оргстекла размерами 7 х 13 см, высотой 10 см, с высотой бортиков 1 см. В ванночку наливалась вода или раствор АзоТАБа и запускалась планария. Концентрация раствора АзоТАБа варьировалась от 50 мкМ до 200 мкМ. Планариум имел два дна, между которыми находилась миллиметровая бумага для численного анализа траектории. Снизу осуществлялась подсветка из светодиодов. Часть планариума освещалась сверху ультрафиолетовым светодиодом. В эксперименте был использован светодиодный модуль (LC-L2, Hamamatsu) 365 нм, который освещал участок ванночки ультрафиолетом. Мощность излучения варьировалась от 0,7 мВт/см2 до 7 мВт/см2. Передвижения планарии фиксировались стандартной веб-камерой [12]. Съемка проводилась покадрово с одинаковым временным интервалом. Время между кадрами съемки варьировалось от 1 секунды до 5 секунд (рис. 1-2).
Для более удобного анализа скоростей, а также эффекта остановки планарии на границе луча ультрафиолета был также изготовлен «одномерный» планариум, в котором червь мог передвигаться только вдоль одной координаты. Длина планариума составляла 13 см при ширине около 0,5 см. Часть одномерного планариума также освещалась ультрафиолетом и исследовалась координата и скорость передвижения в зависимости от градиента и мощности излучения, а также для различных концентраций АзоТАБа (рис. 3-4).
Таким образом, меняя концентрацию АзоТАБа и мощность ультрафиолетового светодиода, мы исследуем статистику поведения планарий. Характеристики установки представлены в таблице 1.
Таблица!
Характеристики установки
УФ-диод LC-L2, Hamamatsu
|
Длина волны А, нм |
365 |
|
Мощность, мВт/см2 |
0,7-7 |
|
Радиус пятна, см |
2,5 |
Веб-камера
|
Разрешение, пиксели |
320 х 240 |
|
Интервал между кадрами, с |
1-5 |
АзоТАБ
|
Концентрация, мкМоль/л |
50-200 |
|
Температура, °C |
20 |
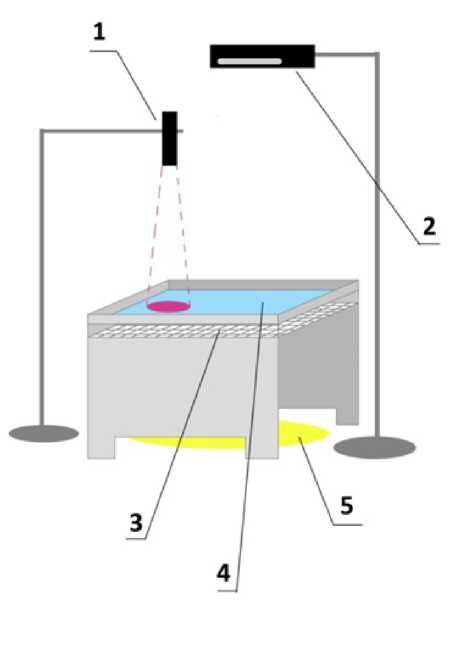
Рис. 1. Схема, установки: 1 — ультрафиолетовый диод, 2 — веб-камера, 3 — миллиметровая бумага, 4 — вода, или раствор АзоТАБа, 5 — светодиодная подсветка.

Рис. 2. Фотография двумерного плаиариума
Двумерный планариум
|
Толщина стенок, мм |
5 |
|
Размеры ванночки, мм х мм |
70 х 130 |
|
Высота, мм |
100 |
Одномерный планариум
|
Толщина стенок, мм |
2 |
|
Размеры ванночки, мм х мм |
5 х 130 |
2.4. Обработка результатов
Набор последовательных изображений, полученных с веб-камеры, обрабатывался программой ImageJ [11]. Для автоматизации обработки изображений был написан плагин для ImageJ, который вычислял положение планарии методом «вычитания фона». Затем он вычислял координату планарии и записывал результат в файл, а. также рисовал траекторию движения червя (рис. 5, 6).
Зная зависимость координаты от времени, можно посчитать скорость планарии от времени. Планариум разбивался на. участки, часть которых была, освещена, ультрафиолетом, а часть нет. Из множества экспериментов производилась выборка — время пребывания в освещенном и неосвещенном участках.
Для численного анализа, скоростей планарии использовался одномерный планариум. Плагин для ImageJ вычислял координату в зависимости от времени и записывал результат в файл. Далее считалась статистика, скоростей в зависимости от координаты для множества, планарий и разброс координат положений планарий.
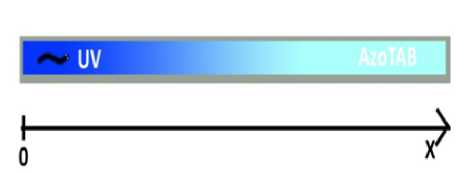
Рис. 3. Одномерный планариум. Червь может двигаться только вдоль координаты х. В результате фототаксиса, плаиария старается покинуть освещенную зону. С возрастанием х возрастает концентрация транс-изомера, который замедляет движение плаиарии вплоть до полной её остановки

Рис. 4. Вид с веб-камеры. Чёрный указатель обозначает текущее положение плаиарии

Рис. 5. Траектория движения плаиарии в воде

Рис. 6. Траектория движения плаиарии па. координатной плоскости
3. Результаты и обсуждение3.1. Схема эксперимента
Для исследования влияния АзоТАБа на. фототаксис необходимо было сравнить движение планарий при различных условиях. Принципиальных схем эксперимента, было две.
-
1. Сравнение траекторий, скоростей и направлений движения планарий в обычной воде, в воде при облучении видимым светом и в воде при облучении ультрафиолетом различной интенсивности (рис. 7). Данные по фототаксису плаиарии сравнивались с контролем: свободными передвижениями плаиарии в отсутствие освещения. Для того чтобы отличить влияние ультрафиолета, от влияния обычного видимого света, эксперименты проводились с лучом видимого света и траектории движения плаиарии сравнивались для всех трех случаев. Таким образом, можно было статистически показать влияние ультрафиолета на поведение планарий.
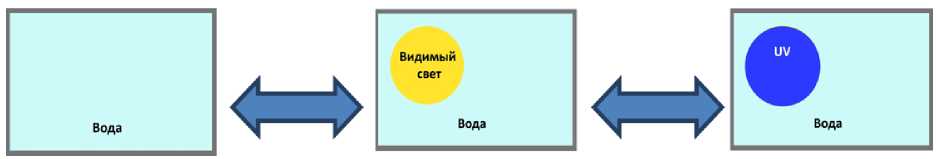
Рис. 7. Схема, эксперимента. 1. Сравнение влияния видимого света, и ультрафиолета, на. плаиарию в воде
-
2. Сравнение траекторий, скоростей и направлений движения планарий в обычной воде, в растворе с различной концентрацией (50-200 мкМоль/л) АзоТАБа в транс-изомере и в цис-изомере при облучении ультрафиолетом (рис. 8). При этом в области освещения ультрафиолетом у планарий начинается отрицательный фототаксис, а АзоТАБ переходит в цис-изомер, что позволяет организму двигаться. При достижении границы освещенной области планария попадает в область с транс-изомером АзоТАБа, который блокирует возбудимость нервных и мышечных клеток, и червь останаливается на границе освещенной области. Так можно исследовать интегральный эффект, оказываемый веществом на организм в целом. Таким образом, можно статитически показать влияние АзоТАБа в трансизомере на поведение планарии, обратимость погашения активности возбудимых клеток, а также влияние цис- и транс-изомеров на фототаксис планарий.
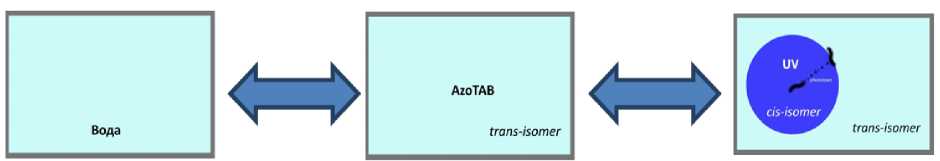
Рис. 8. Схема, эксперимента. 2. Сравнение движений планарии в воде и растворе АзоТАБа. различных концентраций (50-200 мкМоль/л) в транс- и цис-изомерах
-
3.2. Исследование траекторий движения планарии
Влияние видимого света и ультрафиолета на планарию в воде
В воде движение планарии достаточно хаотическое. Доползая до края планариума, червь, как правило, некоторое время двигался вдоль бортика, после чего снова, выползал к центральной части планариума. (рис. 9). При освещении видимым светом принципиально поведение планарии не меняется. Первое время планария избегает света, но через время, порядка, минуты, свет никак не влияет на. траекторию (рис. 10). При освещении ультрафиолетом мощностью 3,5 мВт/см2 планария старается избегать зоны освещения. Покидая зону освещения, планария начинает двигаться хаотически (рис. 11). Разделим планариум на. два. участка: правый и левый. Левый участок будет освещаться видимым светом или ультрафиолетом. Сравним время пребывания планарий в левой части в трех случаях: в воде без освещения, в воде при освещении видимым светом и в воде при освещении ультрафиолетом. В каждом из трех тестов суммарное время движения составляло Т = 2400 с. В воде процент пребывания в левой части Рв = 0,49 ± 0,05; при освещении левой части видимым светом Рс = 0,52 ± 0,05; при освещении ультрафиолетом Руф = 0,29 ± 0,03.
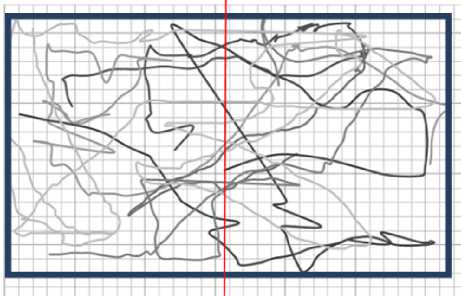
Рис. 9. Набор траекторий планарий в воде. Процент времени пребывания в левой части Рв = 0,49 ± 0,05
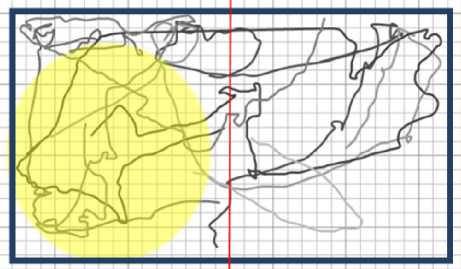
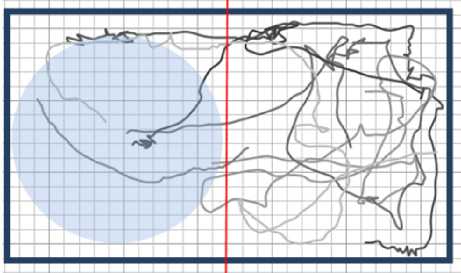
Рис. 10. Набор траекторий плаиарий в воде при освещении видимым светом. Жёлтым обозначена. освещённая область. Процент времени пребывания в левой части Рс = 0,52 ± 0,05
Рис. 11. Набор траекторий плаиарий в воде при освещении ультрафиолетом. Сипим обозначена освещенная зона. Планарии избегают зоны ультрафиолета, т.п. отрицательный фототаксис. Процент времени пребывания в левой части Ру<)> = 0,29 ± 0,03
Влияние АзоТАБа на движение планарии
При увеличении концентрации АзоТАБа. двигательные способности планарии резко падали. При этом АзоТАБ находится в транс-изомере. Сравнительные траектории для воды и растворов АзоТАБа. концентрацией 50 мкМоль/л, 100 мкМоль/л и 200 мкМоль/л приведены на. рис. 12. Во всех четырёх случаях траектории получены за. одинаковое время t = 300 с.
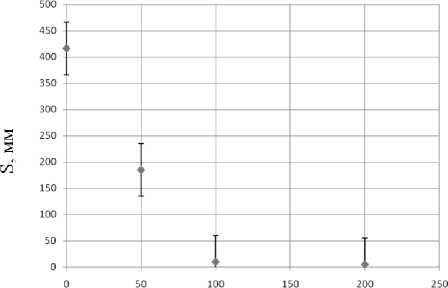
Из зависимости пути, пройденного планарией за. 300-секундный тест, от концентрации раствора. АзоТАБа. видно, что уже при концентрации 100 мкМоль/л активность практически полностью гасится (рис. 12).
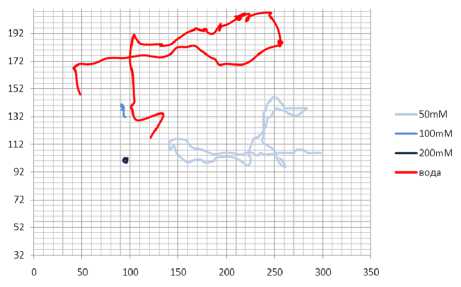
Рис. 12. Сравнительные траектории планарии в воде и растворах АзоТАБа. в транс-форме различной концентрации. Время движения 300 с
Концентрация азотаба, мкМоль/л
Рис. 13. Зависимость пути S, пройденного пла-парией за. 300-секундный тест от концентрации раствора. АзоТАБа.
Влияние АзоТАБа на фототаксис плаиарий
Из и. 3.2 становится понятно, что в растворе АзоТАБа 50 мкМоль/л планария более заторможена, чем в воде, однако способна двигаться. При освещении ультрафиолетом мощ- ностью 3,5 мВт/см2 начинается отрицательный фототаксис, планария старается избегать зоны освещения (рис. 14). Ситуация схожа с обычной водой, только скорость планарии в АзоТАБе меньше (для оценки скоростей был поставлен опыт с одномерным планариумом, см. п. 3.3). В растворе АзоТАБа 100 мкМоль/л планария прекращала двигаться и находилась на одном месте. При освещении ультрафиолетом АзоТАБ переходил в цис-изомер, планария получала возможность двигаться и старалась покинуть зону освещения. На границе освещенной зоны транс-изомер АзоТАБа блокировал движение планарии, червь или продолжал двигаться вдоль края луча или замирал на границе (рис. 15).
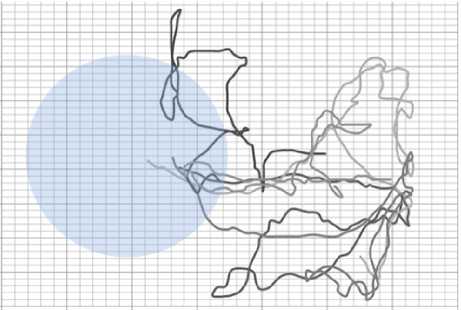
Рис. 14. Набор траекторий плаиарий в растворе АзоТАБа. концентрации 50 мкМоль/л при освещении ультрафиолетом мощностью 3,5 мВт/см2. Планария покидает освещенную зону; концентрации АзоТАБа. недостаточно, чтобы заблокировать двигательные функции
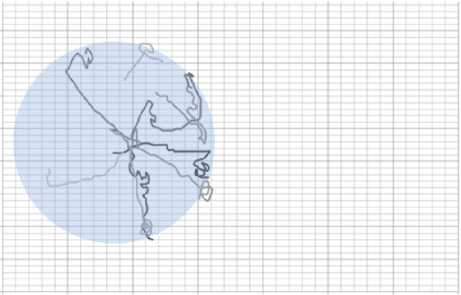
Рис. 15. Набор траекторий плаиарий в растворе АзоТАБа. концентрации 50 мкМоль/л при освещении ультрафиолетом мощностью 3,5 мВт/см2. Планария доходит до границы луча, и попадает в область с транс-изомером АзоТАБа, который блокирует возбудимость нервных и мышечных клеток, и червь останаливается на. границе луча.
3.3. Анализ скоростей планарии
Для анализа, скоростей был использован одномерный планариум. Из графиков зависимости координаты от времени находилась скорость перемещения планарии. В воде плана-рия свободно перемещается из конца, в конец (рис. 16). При облучении части планариума ультрафиолетом планария выползает из области облучения и дальше двигается свободно (рис. 17). В 100 мкМоль/л растворе АзоТАБа. при освещении ультрафиолетом мощностью 3,5 мВт/см2 планария покидает область освещения, постепенно замедляясь, вплоть до полной остановки (рис. 18).
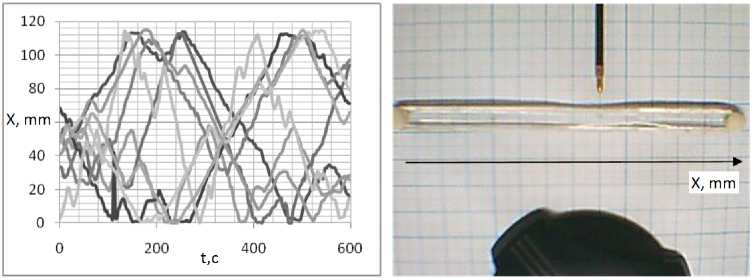
Рис. 16. Зависимость координаты ж положения планарии от времени в воде
Из графиков находим среднюю скорость движения при спокойном движении, при освещении ультрафиолетом мощностью 3,5 мВт/см2, а также в растворах АзоТАБа концентрацией 100 мкМоль/л в цис- и транс-изомерах. Результаты представлены на. рис. 19.
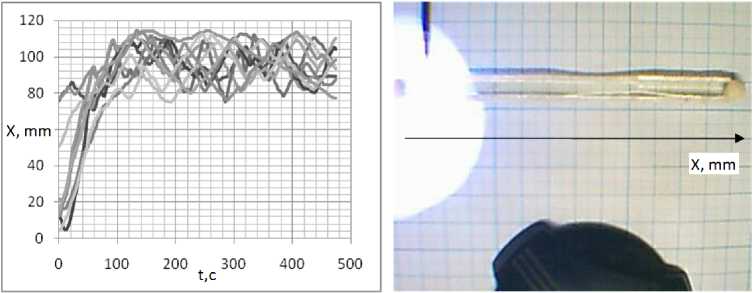
Рис. 17. Зависимость координаты ж положения плаиарии от времени в воде при освещении части плаиариума ультрафиолетом мощностью 3,5 мВт/см2
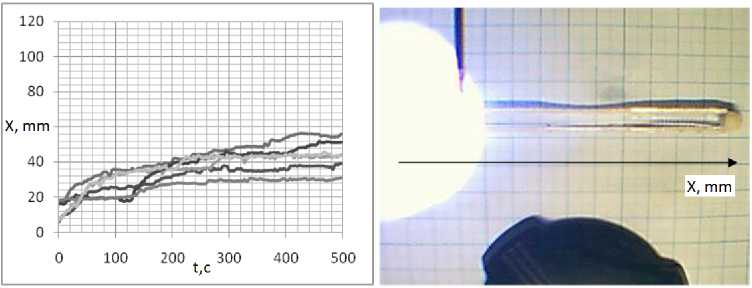
Рис. 18. Движение плаиарии в растворе АзоТАБа. концентрации 100 мкМоль/л при освещении части плаиариума ультрафиолетом мощностью 3,5 мВт/см2
Из диаграммы видно, что скорость при фототаксисе возрастает практически в два раза. Транс-изомер практически полностью блокирует двигательные способности. АзоТАБ в дне-форме частично восстанавливает двигательную активность.
4. Выводы
При проведении исследования траекторий движения планарий в воде, а также в воде при облучении видимым светом и ультрафиолетом было выявлено, что в неосвещенном планариуме процент пребывания плаиарии в левой половине Рв = 0,49 ± 0,05 процент пребывания плаиарии в освещенной видимым светом половине Рс = 0,52 ± 0,05, а процент пребывания плаиарии в освещенной ультрафиолетовым светом половине Руф = 0,29 ± 0,03. Это подтверждает наличие отрицательного фототаксиса у Planaria Torva.
Исследование траекторий свободного движения планарий в воде и в растворах АзоТАБа (транс-изомера) различной концентрации показало, что при увеличении концентрации АзоТАБа двигательная активность плаиарии снижается. В частности, характерные расстояния, пройденные планарией за 300-секундный тест в воде, — So = 420 ± 30 мм, а в растворе АзоТАБа 50 мкМоль/л — S50 = 180 ± 30 мм. Этот факт согласуется с существующими представлениями о влиянии транс-изомера АзоТАБа на возбудимые клетки.
Под действием ультрафиолетового излучения двигательная активность частично восстанавливается, что позволяет нам говорить об обратимости блокирования двигательных функций транс-изомером АзоТАБа.
Измерение скоростей передвижения плаиарии в воде в свободном передвижении и при фототаксисе, а также при фототаксисе в растворе АзоТАБа позволяет говорить нам о конкурирующем действии эффекта фототаксиса и блокирования активности транс-изомером АзоТАБа, что приводит к такому интересному эффекту, как «застревание» плаиарии на
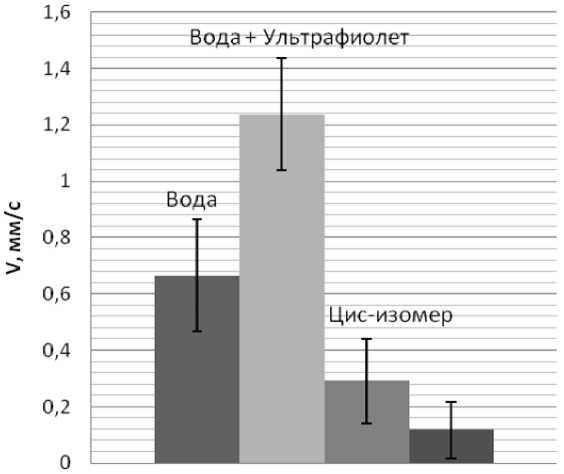
Рис. 19. Средняя скорость передвижения плаиарии в различных средах. Скорость в воде vB = 0,6±0,2 мм/с, в воде при освещении УФ — гіуф = 1,2±0,2 мм/с, в 100 мкМоль/л растворе транс-АзоТАБа — vtr = 0,12±0,10 мм/с, в 100 мкМоль/л растворе гщс-АзоТАБа — vcis = 0,30±0,15 мм/с границе засвеченной ультрафиолетом области в растворе АзоТАБа.
Таким образом, данная работа рассматривает новый подход к фотосенсибилизации целого организма и открывает перспективы в будущем разработать методы контроля поведения и передвижения многоклеточных организмов с нервной системой.
Данная работа была выполнена в рамках исследований, осуществляющихся по гранту № 11.G34.31.0015 от 30 ноября 2010 г. Правительства Российской Федерации для государственной поддержки научных исследований, проводимых под руководством ведущих ученых в российских образовательных учреждениях высшего профессионального образования, а также Министерством образования и науки Российской Федерации.
Список литературы Исследование влияния АзоТАБа на фототаксис Planaria Torva
- Fortin D. L., Banghart M. R. [et al.]. Photochemical control of endogenous ion channels and cellular excitability//Nature Methods. -2008. -V. 5, N 4. -P. 331-338.
- Тирас Х. П., Сахарова Н.Ю. Прижизненная морфометрия планарий//Онтогенез. -1984. -Т. 15, № 1. -С. 42-48.
- Agata K., Soejima Y., Kato K., Kobayashi C., Umesono Y., Watanabe K. Structure of the planarian central nervous system (CNS) revealed by neuronal cell markers//Zoological Science. -1998. -V. 15. -P. 433-440.
- Carpenter K. S., Morita M., Andbest J. B. Ultrastructure of the photoreceptor of the planarian Dugesia dorotocephala//Journal of Ultrastructure Research. -1974. -N 148. -P. 143-158.
- MacRae E. K. Observations on the fine structure of photoreceptor cells in the planarian Dugesia tigrina//Journal of Ultrastructure Research. -1964. -N 10. -P. 334-149.
- Inoue T., Kumamoto H., Okamoto K. [et al.]. Morphological and functional recovery of the planarian photosensing system during head regeneration//Zoological Science. -2004. -V. 21. -P. 275-283.
- Magome N., Agladze K. I. Patterning and excitability control in cardiomyocyte tissue culture//Physica D. -2010. -V. 239. -P. 1560-1566.
- Donaldson S. H. Jr., Lee C. T. Jr., Chmelkaa B. F., Israelachvili J. N. General hydrophobic interaction potential for surfactant/lipid bilayers from direct force measurements between light-modulated bilayers//PNAS. -2011. -V. 108, N 38. -P. 15699-15704.
- Erofeev I. S., Magome N., Agladze K. I. Digital photocontrol of the network of live excitable cells//Письма в ЖЭТФ. -2011. -Т. 94, № 6. -С. 513-516.
- Cebri`a F., Kudome T., Nakazawa M., Mineta K., Ikeo K., Gojobori T., Agata K. The expression of neural-specific genes reveals the structural and molecular complexity of the planarian central nervous system//Mechanisms of development. -2002. -V. 116, N 1 -2. -P. 199-204.
- Abramoff M. D., Magalh˜aes P. J., Ram S. J. Image processing with ImageJ. -Laurin Publishing, 2004. -ISSN 1081-8693.
- Конушин А. C., Ветров Д. П., Воронин П. А., Синдеев М. С., Ломакина-Румянцева Е. И. Система видеонаблюдения за поведением лабораторных животных с автоматической сегментацией на поведенческие акты//Труды конференции Graphicon-2008. -2008. -P. 199-205.
