Исследование влияния радиационного фактора на структурно-анатомические показатели растений в условиях модельного эксперимента

Автор: Сысоева Е.С., Поливкина Е.Н., Паницкий А.В.
Рубрика: Научные статьи
Статья в выпуске: 1 т.34, 2025 года.
Бесплатный доступ
Ионизирующее излучение, как экологический фактор, приводит к адаптационным процессам, влияющим на выживаемость, рост и развитие растений. Листовая пластина является перспективным органом-индикатором структурно-анатомических изменений вследствие накопления радиоактивных веществ. Цель данной работы заключалась в изучении влияния радиационного фактора на структурно-анатомические показатели листьев Phaseolus vulgaris L. в 3-х последовательных поколениях в условиях модельного эксперимента при корневом поступлении 90Sr. Растения выращивали на почвенных образцах с высокой удельной активностью 90Sr (5´108 Бк/кг), отобранных на территории бывшего Семипалатинского испытательного полигона. В качестве основных структурно-анатомических параметров листа использовали толщину адаксиального и абаксиального эпидермиса, а также мезофилла. Отбор проб листьев производили после их полного формирования, затем консервировали Копенгагенской смесью. Мощность дозы от внутреннего и внешнего облучения для Phaseolus vulgaris L. за счёт 90Sr составила 9,5´10-1 Гр, что указывает на проявление адаптивных реакций, а не эффект гормезиса. Распределение 90Sr по органам Phaseolus vulgaris L. представлено убывающим рядом «листья>стебли>плоды». Установлены достоверные изменения толщины адаксиального и абаксиального эпидермиса, а также мезофилла листовой пластины (при p90Sr.
Ионизирующее излучение, листовая пластина, мезофилл, адаксиальный эпидермис, абаксиальный эпидермис, гомеостаз, структурно-анатомические параметры, радиобиология, охрана окружающей среды
Короткий адрес: https://sciup.org/170209556
IDR: 170209556 | УДК: 539.1.047:581.15 | DOI: 10.21870/0131-3878-2025-34-1-95-105
The use of experimental model for investigating response of plants structures and anatomy to radiation
Ionizing radiation as environmental factor, causes adaptive processes that affect plant and anatomical changes caused by the accumulation of radioactive substances. This study aimed to investigate the effect of radiation on the structure and anatomy of Phaseolus vulgaris L. leaves in three generations under conditions of a model experiment involving root uptake of 90Sr. Plants were grown in soil samples with high levels of specific activity of 90Sr (5´108 Bq/kg), selected from the former Semipalatinsk testing site. The adaxial and abaxial epidermal layers and mesophyll thickness were used as the main structural and anatomic parameters. Leaf samples were taken after full formation and preserved in a Copenhagen mixture before irradiation. The dose rates from internal and external radiation for Phaseolus vulgaris L. were determined. Due to 90Sr being 9.5´10-1 Gy, this indicates the manifestation of adaptive responses, rather than the effect of hormetic response. The distribution of 90Sr among the organs of Phaseolus vulgaris L. is represented by a series decreasing from “leaves>stems>fruits”. Significant changes in the thickness of adaxial and abaxial epidermal layers, as well as mesophyll thickness in the leaf lamina (p90Sr accumulates and is fixed.
Текст научной статьи Исследование влияния радиационного фактора на структурно-анатомические показатели растений в условиях модельного эксперимента
В настоящее время особое внимание уделяют вопросу о способности растений адаптироваться к быстро изменяющимся условиям окружающей среды, которые могут оказывать негативное воздействие на природные популяции и экосистемы в целом [1, 2]. Пластичность и генетический ресурс растений, необходимые для адаптации, могут истощаться из-за увеличения интенсивности стрессовых воздействий [3]. Так как растения не могут избежать стрессовых условий среды, их стратегии минимизации последствий стрессовых воздействий ограничены специфическими изменениями в метаболических путях и модификацией экспрессии генов [4].
Ионизирующее излучение, как экологический фактор, приводит к адаптационным процессам, связанным с выживаемостью, ростом и развитием растений. Они отражают способность к динамическому равновесию между организмом и изменяющимися условиями среды существования, в частности радиационным фоном [5]. Каждому виду растений присущи определённые
границы адаптивной нормы к ионизирующим излучениям. Оптимальный уровень определяется естественным радиационным фоном. После облучения в растительных популяциях наблюдаются процессы восстановления их морфологической и анатомической структуры, в основном за счёт отмирания наиболее повреждённых клеток, а также в результате восстановительных процессов на клеточном и онтогенетическом уровнях [6]. Однако, исследования, проведённые рядом авторов через несколько лет после аварии на Чернобыльской АЭС в популяциях травянистых фитоценозов этой зоны, показали, что ионизирующее излучение оказывает угнетающее действие на некоторые виды растений: повышается число морфологических аномалий, изменяется структура популяций [7]. При этом отмечено и стимулирующее действие радиационного излучения на посевные качества семян древесных растений этой же зоны и повышение их устойчивости к облучению [8].
Под влиянием радиационного фактора наблюдаются изменения на различных уровнях биологической организации. На молекулярно-генетическом – генные мутации, нарушение воспроизведения генетического материала, обмена веществ. Изменения на органно-тканевом уровне могут заключаться в нарушении строения и функций тканей, и, как следствие, органов. Изменения же в популяциях и экосистемах включают в себя адаптацию к меняющимся условиям среды, нарушение биологического круговорота веществ и энергии [9].
В зависимости от условий облучения и вида растения радиационное воздействие может приводить как к ингибирующим, так и к стимулирующим эффектам на морфологическом уровне [10]. В частности, при увеличении α - и β -активности почвы происходит изменение внутренней структуры стебля: первичная кора, размеры паренхимных клеток, размеры ксилемных сосудов увеличиваются [11, 12]. В листьях изменяется форма эпидермальных клеток, их количество, однако, число слоёв столбчатого мезофилла остаётся неизменным. При этом, за счёт увеличения размеров клеток происходит утолщение листовой пластинки, а также увеличение числа проводящих элементов [13, 14]. Растительные популяции, подвергающиеся хроническому облучению, могут характеризоваться, с одной стороны, ускоренным мутационным процессом, а с другой – различными в зависимости от мощности дозы уровнями активации систем адаптации к стрессовым факторам окружающей среды [15]. В данном аспекте листовая пластина является перспективным органом-индикатором изменений структурно-анатомических показателей вследствие накопления радиоактивных веществ как при корневом, так и при фоллиарном поступлении. Таким образом, цель проведённого исследования заключалась в изучении влияния радиационного фактора на структурно-анатомические показатели листьев 3-х последовательных поколений Phaseolus vulgaris L. в условиях модельного эксперимента при корневом поступлении радионуклида 90Sr.
Материалы и методы
Модельный эксперимент
В качестве экспериментальной культуры использовали фасоль обыкновенную (Phaseolus vulgaris L.). Выбор исследуемой культуры обусловлен коротким вегетационным периодом (114-127 сут), устойчивостью к вредителям и болезням. Выращивание 3-х последовательных поколений Phaseolus vulgaris L. проводилось в контролируемых условиях экспериментальной оранжереи на почвенных образцах с территории бывшей испытательной площадки «4а» (рис. 1). Контрольную группу растений выращивали на фоновой почве с идентичными физико-химическими свойствами. В период с 1953 по 1957 гг. на площадке «4а» проводились исследовательские работы по созданию радиологического оружия в ракетном и бомбовом исполнении (оружие массового поражения невзрывного типа – боеприпасы с боевыми радиоактивными веществами (БРВ)). В качестве боевого снаряжения использовали жидкие и порошкообразные рецептуры, представлявшие собой радиоактивные отходы атомной промышленности, а также продукты, полученные после облучения специально подобранных веществ нейтронами работающего атомного реактора, т.е. путём образования наведённой активности. Испытания включали рассеяние БРВ путём подрыва отдельных снарядов, сброса бомб или распыления БРВ с самолётов. Почвы данной территории относятся к типу зональных светло-каштановых нормальных, неполноразвитых, характеризующихся защебнённостью [16]. Основные физико-химические свойства почвы, исполь- зованной в эксперименте, представлены в табл. 1.
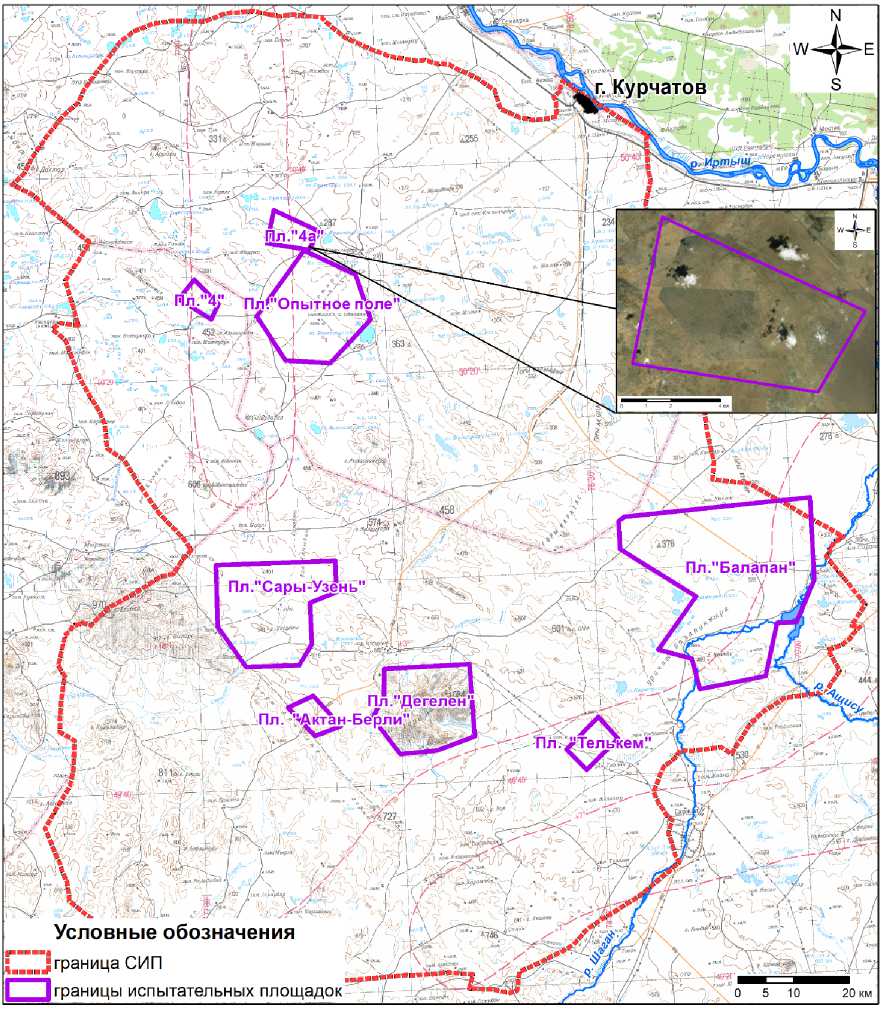
Рис. 1. Схематичное расположение площадки «4а» на территории Семипалатинского полигона.
Таблица 1
Физико-химические свойства экспериментальных почвенных образцов
|
Гумус, % |
рН вод |
Ʃ солей (ммоль/ 100 г) |
Ca обмен (ммоль/ 100 г) |
Физ. глина (<0,01 мм), % |
Ил (<0,001 мм), % |
Csвa л , мгхкг-1 |
Srвa л , мгхкг-1 |
CaBan, мгхкг-1 |
|
3,2 |
6,9 |
2,7 |
3 |
25,2 |
6,3 |
0,4±0,1 |
25±3 |
(2,0±0,03)х10-3 |
Как видно из табл. 1, почвы, использованные в эксперименте, по механическому составу относятся к легкосуглинистым (содержание физической глины 25,2%) с низким содержанием гумуса (3,2%) и слабокислой реакцией почвенного раствора (рН вод – 6,9).
Основным загрязнителем в почве данной площадки является радионуклид 90Sr, удельная активность которого в исследуемых почвах достигает 5 x 10 8 Бк/кг [16].
Посев 3-х последовательных поколений Phaseolus vulgaris L. производился сухими семенами (рис. 2). Масса семян, используемых при посадке, составила 1,5-2 г, при длине 2-2,5 см. В ходе эксперимента на протяжении всего вегетационного цикла поддерживалась оптимальная влажность почвы (50-60% от полной влагоёмкости), а также обеспечивались достаточный уровень освещённости (10000 Лк) и благоприятный температурный режим (20-27 °С) посредством систем фитоосвещения и терморегулирования.
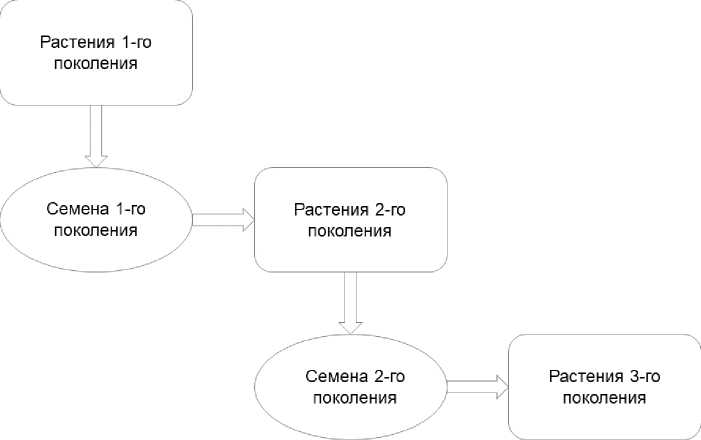
Рис. 2. Схема вегетационного эксперимента.
Образцы листьев отбирали после их полного формирования в конце вегетационного развития в 3-кратной повторности, затем консервировали с использованием Копенгагенской смеси (70% спирта, 27% воды и 3% глицерина) [17]. Из консервированных образцов получали срезы при помощи санного микротома МЗП-01 «Техном» (РФ). Фрагмент листа, помещённый на замораживающее устройство микротома, перед каждым движением микротомного ножа автоматически поднимался на заданную высоту (толщину среза), которая зависела от состояния растения и жёсткости волокон. В среднем толщина среза для листа исследуемой культуры составила 30-60 мкм. Полученный срез ткани листа осторожно смывали с ножа на предметное стекло при помощи пипетки и дистиллированной воды, затем остатки воды удаляли фильтровальной бумагой и заме- няли её на 98% глицерин, поверх накладывали покровное стекло. Исследования полученных микропрепаратов проводили при помощи микроскопа Micros MC 300, с камерой Vision Cam V500/21 M (Австрия) при 4, 10, 20, 40 и 100х увеличении. Измерения структурно-анатомических параметров листовой пластины производили с использованием ПО BioWizad 4.2.
В качестве основных исследуемых параметров листа использовали толщину мезофилла, адаксиального и абаксиального эпидермиса (рис. 3).
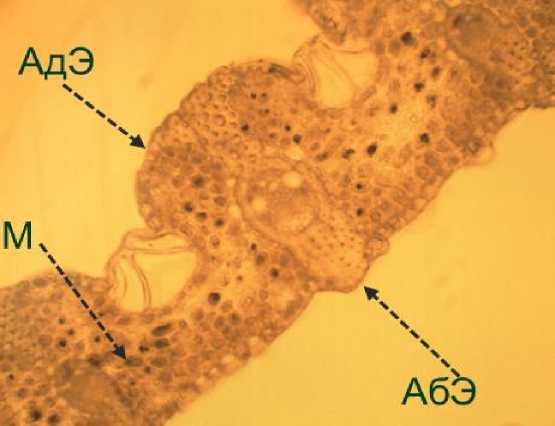
Рис. 3. Поперечный срез листа Phaseolus vulgaris L. (увеличение 20 х ).
АдЭ – адаксиальный эпидермис; АбЭ – абаксиальный эпидермис; М – мезофилл.
Лабораторые исследования
Радионуклидный анализ. Пробы растений промывали и ополаскивали дистиллированной водой, затем высушивали при температуре 80-100 °С в сушильном шкафу до постоянной массы, после чего подвергались грубому измельчению до длины 1-3 см при помощи секатора. Более тонкое измельчение проводилось на лабораторной мельнице. Измельчённая проба обугливалась в муфельной печи при начальной температуре 200-250 °С, с постепенно повышающейся температурой до 350-400 °С. Обугленные пробы передавали на радиохимическое выделение с последующим бета- и альфа-спектрометрическим измерением.
Пробы почвы высушивали до воздушно-сухого состояния в сушильном шкафу при температуре 60-70 °С, затем после удаления включений весь объём пробы тщательно перемешивали, истирали в фарфоровой ступке и просеивали через сито с диаметром отверстий 1 мм. После этого методом квартования отбирали необходимые навески почвы для определения удельной активности 90Sr.
Измерение удельной активности радионуклидов в пробах почвы и растений осуществляли в соответствии со стандартизированными методическими указаниями [18, 19]. Радионуклид 90Sr определяли радиохимическим выделением с последующим измерением на бета-спектрометре TRI-CARB для растительных проб и на бета-спектрометре «Прогресс» для проб почвы. Предел обнаружения для 90Sr составил 1-20 Бк/кг, а погрешность измерений не превышала 15%.
Контроль качества. Исследования выполняли с использованием аналитического и испытательного оборудования, прошедшего поверку согласно Закону Республики Казахстан от 7 июля 2000 г № 53-II «Об обеспечении единства измерений». Для контроля качества результа- тов анализа в каждую партию из 10 исследуемых образцов добавляли один «повторный», который рандомно выбирали из исследуемой партии. Так же, для контроля возможного перекрёстного загрязнения образцов, в партию из 10 проб вводили «контрольный» образец, который формировался из образцов с известным содержанием радионуклидов. Анализ «повторного» и «контрольного» образцов проводили одновременно со всеми остальными образцами анализируемой партии.
Статистическая обработка результатов. Всего подготовлено 300 микропрепаратов листа, произведено около 2400 измерений 3-х структурно-анатомических параметров листа Phaseolus vulgaris L. Для каждого из исследуемых параметров определяли минимальные и максимальные значения, рассчитывали среднее арифметическое и ошибку среднего арифметического, медиану, среднеквадратичное отклонение и коэффициент вариации.
Перед началом обработки полученных данных проводили проверку выпадов для исключения из полученных выборок резко отличающихся значений (артефактов). Проверка данных производили по критерию, равному нормированному отклонению исследуемого выпада [20].
T = — >Tst , (1)
где T – критерий выпада; V – выделяющееся значение (артефакт); М – среднее арифметическое для исследуемой выборки; σ – среднее квадратичное отклонение для исследуемой выборки; Т st – стандартные значения критерия выпадов.
Для сравнения эмпирического распределения с теоретическим применяли критерий Колмогорова-Смирнова ( X ), который рассчитывается согласно выражению:
^ эмпир ^, (2)
где D – максимальное значение абсолютной разности между накопленными частотами эмпирического и теоретического распределений; n – объём выборки.
Полученные значения критерия (при уровне значимости р=0,05) во всех случаях меньше теоретических значений, что говорит о нормальном распределении значений в выборке [21].
Результаты и обсуждение
В результате исследования установлено, что наиболее интенсивное накопление 90Sr в органах Phaseolus vulgaris L. характерно для листовой пластины (~ 4,4 x 10 6 Бк/кг), меньшее содержание зафиксировано в стеблях (~ 2,3 x 10 6 Бк/кг), минимальное накопление, с разницей на один порядок, отмечено в плодах исследуемой культуры (~ 7,7 x 10 5 Бк/кг).
Оценка дозовых нагрузок внутреннего и внешнего облучения растений проводилась согласно рекомендациям Публикации 108 МКРЗ [22]. Мощность дозы внутреннего и внешнего облучения растений рассчитывалась как произведение удельной активности радионуклида в растении на соответствующий дозовый коэффициент:
D=Axd , (3)
где A – удельная активность сырой пробы растения или удельная активность подстилающей почвы (Бк/кг); d – дозовый коэффициент внутреннего или внешнего облучения растения (мкГр/сут)/(Бк/кг).
Согласно проведённым расчётам, мощность внутренней поглощённой дозы за счёт 90Sr для Phaseolus vulgaris L. за вегетационный период (114-127 сут) составила 9,5x10-1 Гр, доза от внешнего облучения незначительна и составляет порядка 1,7x10-4 Гр. Суммарная мощность до- зы от внутреннего и внешнего облучения для исследуемой культуры превышает предел малых доз (0,1 Гр) [23, 24]. Таким образом, растения проявляют адаптивные реакции в ответ на высокие дозы ионизирующего излучения, значительно отличающиеся от гормезиса.
Таблица 2
Средние значения анатомических параметров листа
|
Параметр |
Вариационно-статистические показатели |
||||||
|
Диапазон изменения |
x ±SD, мкм (о) CV, % |
Ме |
|||||
|
эксп. |
контр. |
эксп. |
контр. |
эксп. |
контр. |
||
|
1-ое поколение (n=300) |
|||||||
|
s ад.э. |
8-19 |
7-14 |
13±0,2 (3) 19 |
10±0,1 (1,5) 14 |
13 |
10 |
|
|
s аб.э. |
8-22 |
7-17 |
15±0,2 (3) 21 |
12±0,1 (2) 15 |
15 |
12 |
|
|
s мф. |
104-282 |
83-177 |
202±3 (43) 21 |
125±2 (27) 21 |
199 |
113 |
|
|
2-ое поколение (n= |
300) |
||||||
|
s ад.э. |
8-16 |
7-14 |
12±0,1 (2) 13 |
10±0,1 (2) 18 |
12 |
10 |
|
|
s аб.э. |
9-19 |
7-17 |
13±0,1 (2) 14 |
12±0,2 (2) 19 |
14 |
12 |
|
|
s мф. |
102-258 |
83-177 |
194±2 (29) 15 |
124±2 (25) 20 |
190 |
117 |
|
|
3-е |
поколение (n=300) |
||||||
|
s ад.э. |
8-16 |
7-14 |
11±0,2 (1,7) 14 |
10±0,1 (2) 18 |
11 |
10 |
|
|
s аб.э. |
9-19 |
7-17 |
13±±0,1 (2) 16 |
12±0,1 (2) 20 |
13 |
12 |
|
|
s мф. |
102-258 |
83-177 |
155±2 (25) 18 |
123±2 (22) 21 |
154 |
114 |
|
Примечание: s ад.э. – толщина адаксиального эпидермиса, мкм; s аб.э. – толщина абаксиального эпидермиса, мкм; s мф. - толщина мезофилла, мкм; x ±SD - среднее арифметическое, ошибка среднего арифметического; Ме – медиана; σ – среднеквадратичное отклонение; CV – коэффициент вариации, %.
На основании результатов исследования для листьев Phaseolus vulgaris L. установлены достоверные изменения изучаемых структурно-анатомических показателей (при p<0,05) в 3-х последовательных поколениях. Согласно полученным данным (табл. 2) наиболее высокие значения исследуемых параметров отмечены у листьев растений 1-го поколения: толщина адаксиального эпидермиса – 13 мкм, толщина абаксиального эпидермиса – 15 мкм, толщина мезофилла – 202 мкм. Более низкие значения характерны для листовой пластины 2-го и 3-го поколений: толщина адаксиального эпидермиса – 12 и 11 мкм, толщина абаксиального эпидермиса – 13 мкм, толщина мезофилла – 194 и 155 мкм. Минимальные же значения структурно-анатомических параметров отмечены для листьев контрольной группы: толщина адаксиального эпидермиса – 10 мкм, толщина абаксиального эпидермиса – 12 мкм, толщина мезофилла – 125 мкм.
Необходимо отметить, что в большинстве случаев значение медианы почти совпадает со средним значением структурно-анатомических параметров как в экспериментальной, так и в контрольной выборках. Данный факт указывает на однородность исследуемых данных, что подтверждают коэффициенты вариации, значения которых во всех случаях значительно ниже 50%. Так, максимум вариации отмечен для абаксиального эпидермиса и мезофилла (26%), а минимум – для адаксиального эпидермиса (13%).
Толщина мезофилла в экспериментальной группе 1-го, 2-го и 3-го поколений больше, чем в контрольной в среднем на 38, 36 и 21%, толщина адаксиального эпидермиса – на 23, 17 и 9%, а абаксиального – на 20 и 8% соответственно. В листьях растений 3-го поколения толщина адаксиального и абаксиального эпидермиса на 15 и 13% достоверно меньше по сравнению с 1-ым поколением.
Увеличение толщины тканей Phaseolus vulgaris L. может быть обусловлено тем, что при неблагоприятных условиях в клетках растений возрастает содержание протекторных веществ (углеводов, аминокислот), участвующих в защитных реакциях и стабилизирующих состояние цитоплазмы, что приводит к увеличению размеров клеток и толщины тканей [25, 26]. Также, накапливаясь в апопласте листовой пластины, 90Sr образует слабые связи с моносахаридами, в результате чего может возрастать пластичность клеточных оболочек [27, 28]. В совокупности действие этих факторов, вероятно, спровоцировало увеличение размера клеток листьев Phaseolus vulgaris L., и, соответственно, тканей. В результате установленных изменений, произошедших в листовой пластине, увеличивается площадь для фотосинтеза, замедляется проникновение поллютантов в более глубокие слои тканей, уменьшается потеря воды через испарение (транспирацию), что способствует поддержанию гомеостаза организма на клеточном уровне биологической организации.
Заключение
В результате проведённых исследований на примере культуры Phaseolus vulgaris L. выявлены механизмы адаптивных реакций на ионизирующее излучение вследствие интенсивного накопления 90Sr листовой пластиной, приводящие к поддержанию гомеостаза организма на клеточном уровне. Установлено, что ионизирующее излучение вызывает увеличение толщины мезофилла, адаксиального и абаксиального эпидермиса листовой пластины.
Наиболее высокие значения исследуемых структурно-анатомических параметров отмечены у растений 1-го поколения, более низкие значения характерны для листовой пластины 2-го и 3-го поколений, минимальные значения отмечены для контрольной группы, что подтверждает особую роль листовой пластины как органа, в котором происходит накопление и фиксация поллютантов, в частности 90Sr.
Результаты также могут быть применены при разработке принципов и методов оценки состояния биоценозов и экосистем в условиях радиоактивного загрязнения.
Исследования выполнены в рамках научно-технической программы BR24792713 «Развитие атомной энергетики в Республике Казахстан».
