Исследование взаимодействия лектинов азоспирилл с β-глюкозидазой сладкого миндаля

Автор: Аленькина С.А., Никитина В.Е.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 1-1 т.11, 2009 года.
Бесплатный доступ
Показано, что лектины, выделенные с поверхности почвенных азотфиксирующих бактерий Azospirillum brasilense Sp7 и его мутанта по лектиновой активности A. brasilense Sp7.2.3 ингибируют активность β-глюкозидазы сладкого миндаля. Ингибирование фермента лектинами соответствует конкурентному типу с константами ингибирования (Кi) 14±0,2 мкг/мл для лектина A. brasilense Sp7 и 4±0,1 мкг/мл для лектина A. brasilense Sp7.2.3. В присутствии этанола и ряда солей (KCl, NaCl, СаCl2 - для родительского штамма и KCl, NaCl - для мутантного), происходит усиление ингибирования лектинами активности фермента, что позволило сделать вывод о том, что взаимодействие лектинов с ферментом обусловлено электростатическими и гидрофобными взаимодействиями.
Азоспириллы, лектины, β-глюкозидаза сладкого миндаля, ингибирование, электростатическое взаимодействие, гидрофобное взаимодействие
Короткий адрес: https://sciup.org/148198235
IDR: 148198235 | УДК: 577.122
Study of the interaction of azospirillum lectins with sweet almond β- glucosidase
The surface lectins isolated from the nitrogen-fixing soil bacterium Azospirillum brasilense Sp7 and from its mutant defective in lectin activity, A. brasilense Sp 7.2.3, inhibited the activity of sweet almond β-glucosidase. Enzyme inhibition by the lectins was of the competitive type, with inhibition constants (Ki) of 14±0,2 µg/ml for the A. brasilense Sp7 lectin and 4±0,1 µg/ml for the A. brasilense Sp7.2.3 lectin. Lectin inhibition of the enzymes activity was enhanced in the presence of ethanol and several salts (KCl, NaCl, and CaCl2 for the parent strain, and KCl and NaClfor the mutant strain), which permitted the conclusion that the lectin-enzyme interaction is conditioned by electrostatic and hydrophobic interactions.
Текст научной статьи Исследование взаимодействия лектинов азоспирилл с β-глюкозидазой сладкого миндаля
Образование азотфиксирующих систем, подобно как и любых других биологических межклеточных взаимодействий, согласно современным представлениям, включает функционирование молекул белковой природы — лектинов. Ранее с поверхности клеток Azospirillum brasilense Sp7 был выделен лектин. Лектин являлся гликопротеином, имел молекулярную массу 36 кВа, проявлял специфичность к L -фукозе (1,87 мМ) и к D -галактозе (20 мМ) [1]. Клетки мутантного штамма имели на поверхности лектиновые молекулы с той же молекулярной массой и углеводной специфичностью, что и родительские. Лектин, выделенный с поверхности мутантных клеток в результате транспозонового мутагенеза претерпел изменения, о чем говорит тот факт, что антитела, полученные к лектину A. brasilense Sp7, не взаимодействовали с лектином мутантных клеток [2]. Впервые обнаруженные нами агглютинирующие белки или лектины у азоспирилл и доказанное участие их в прикреплении к корням растений дали возможность предположить более активную роль лектинов в процессах,
Светлана Александровна Аленькина, старший научный сотрудник лаборатории микробиологии и микологии; Валентина Евгеньевна Никитина, заведующая той же лаборатории.
вносящих свой вклад в эффективность взаимодействия бактерий с растением-хозяином [3]. Было показано, что лектины азоспирилл способны модифицировать активность собственных и растительных гидролитических ферментов [4, 5]. Несмотря на то, что в литературе описаны случаи регуляции лектинами ферментов в клетках различного происхождения, данные о механизмах этих взаимодействий отсутствуют.
Целью настоящей работы явилось исследование кинетических параметров, характеризующих связывание лектинов азоспирилл с р -глюкозидазой сладкого миндаля (модельная система), а также выяснение особенностей механизма образования данного белок-белкового комплекса, в частности уточнение роли гидрофобных и электростатических взаимодействий.
В тексте использованы следующие сокращения: НФГП - 4-нитрофенил- -D-глюкопиранозид.
МЕТОДИКА
В работе были использованы лектины двух штаммов — Azospirillum brasilense Sp7, полученного из Института микробиологии РАН (г. Москва), и его мутанта, дефектного по лектиновой активности — Azospirillum brasilense Sp7.2.3 [2]. Культуры азоспирилл выращивали на жидкой синтетической среде для флоккуляции при 37°С в течение 18 ч [6]. Выделение и очистку лектинов проводили как описано ранее [7, 8].
Ингибирующее влияние лектинов на в -глюкозидазу («Serva») определяли по их влиянию на реакцию гидролиза 4-нитрофенил- в - D -глюкопиранозида. Для этого 0,1 мл фермента (3,7 мг/мл) и 0,1 мл лектина (40 мкг/мл) выдерживали 30 мин, а затем инкубировали с 0,1 мл НФГП (2 мМ в 0,1 М ацетатном буфере рН 4,6) 1ч при 37°С. После чего реакцию останавливали добавлением 0,4 мл 0,005 М глицин/NaOH буфера (pH 9,6). Количество образовавшегося нитрофенола определяли спектрофотометрическим методом при X = 425 нм (СФ-26) [9]. В качестве контроля были взяты фракции, не содержащие лектин.
Начальную скорость реакции расщепления НФГП определяли по наклону касательной к исходному отрезку кинетической кривой на начальном участке в течение 6 мин. Константу ингибирования определяли по уравнению Михаэлиса для случая конкурентного ингибирования [10]. Кинетические константы (Кт и Умакс) определяли из графика Корниш-Боудена как точки пересечения прямых с абсциссой и ординатой [11].
При изучении совместного действия лектинов с этанолом и солями в инкубационную смесь, состоящую из фермента и лектина добавляли по 0.1 мл этанола до 30 % ( v/v ) или 0,1 мл 100 мМ NaCl, КС1, СаС12, NH 4 C1.
Экспериментальные данные обрабатывали статистически с использованием t -критерия Стьюдента [12].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
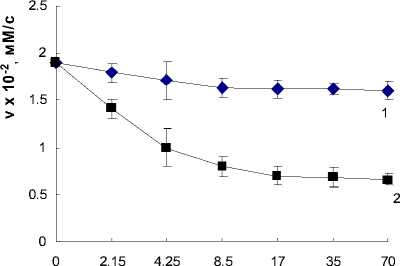
В более ранних работах [5] была обнаружена способность лектинов азоспирилл модифицировать активность гидролитических ферментов корней пшеницы. Используя модельную систему, включающую лектины изучаемых штаммов азоспирилл и коммерческий препарат в -глюкозидазы сладкого миндаля было показано, что лектины обоих штаммов в увеличивающихся концентрациях вызывали уменьшение активности фермента (рис. 1).
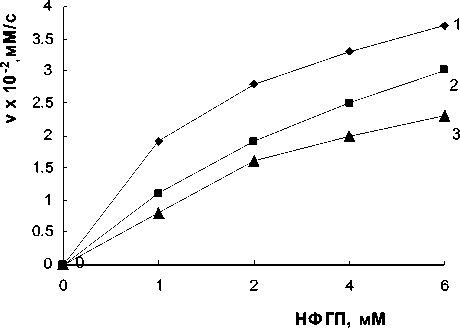
На рис. 2 а показаны кривые зависимости
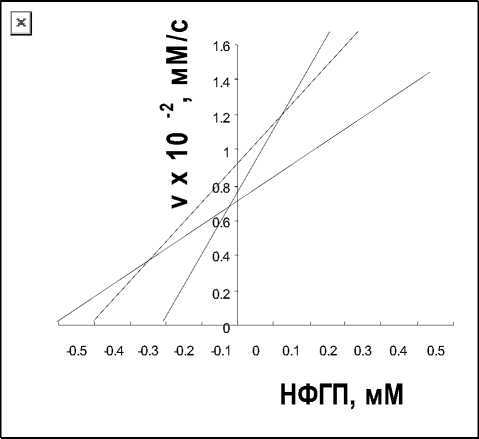
____________ Проблемы прикладной экологии скорости реакции от концентрации субстрата для обоих лектинов. Те же данные представлены на рис. 2 6 для A. brasilense Sp7 и A. brasilense Sp7.2.3 в координатах Корниш-Боудена. Произошло увеличение К m по сравнению с контрольным вариантом. Для A. brasilense Sp7 она составила 0,46±0,05тМ, для A. brasilense Sp7.2.3. — 0,5±0,02тМ.
конц. лектинов, мкг/мл
Рис. 1. Скорость реакции гидролиза n-нитрофенилф-О-глюкопиранозида в условиях ингибирования лектинами: 1 — A. brasilense Sp7; 2 — A. brasilense Sp7.2.3; условия реакции: 2мМ НФГП; концентрация в -глюкозидазы — 3,7 мг/мл; 0,1 М ацетатный буфер, рН 4,6; 37°С
В то же время максимальная скорость реакции при действии лектинов осталась такой же, как и в варианте без лектинов. Изменения кинетических констант и форма графика, характеризующего зависимость между скоростью реакции и концентрацией субстрата, свидетельствуют о том, что лектины азоспирилл являются конкурентными ингибиторами для в -глюкозидазы. Обычно при рассмотрении конкурентного ингибирования предполагается, что ингибитор стерически сходен с субстратом (полностью или частично), вследствие чего он как бы подменяет субстрат, связываясь с соответствующим участком в молекуле фермента. Видимо, в углеводной части лектинов содержатся участки, структурно схожие с субстратом, вследствие чего возникает конкуренция. С целью получения более точной количественной оценки ингибирующего влияния лектинов на активность фермента были определены
Известия Самарского научного центра Российской академии наук. 2009. Т. 11, № 1
значения констант ингибирования. Константа ингибирования (Кi) 14±0,2 мкг/мл — для лектина A. brasilense Sp7 и 4±0,1 мкг/мл — для лектина A. brasilense Sp7.2.3. Как видно из результатов, лектины родительского и мутантного штаммов имеют различное сродство к ферменту, что, вероятно, объясняется структурными различиями молекул.
а.
A. brasilense Sp7
A. brasilense
Sp7.2.3
б.
Рис. 2. а — зависимость активности р -глюкозидазы (1) от концентрации п-нитрофенил- р -D-глюкопиранозида в условиях ингибирования лектинами A. brasilense Sp7 (2) и A. brasilense Sp7.2.3 (3). Концентрация лектинов — 10 мкг/мл; р -глюкозидазы — 3,7 мг/мл; 0,1 М ацетатный буфер, рН 4,6; 37°С; б — соответствующие графики в координатах Корниш-Боудена
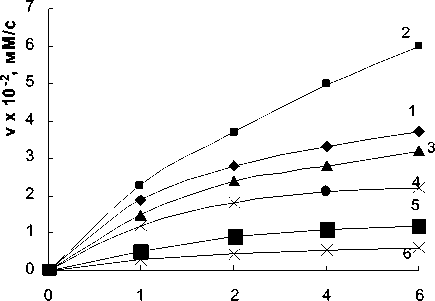
НФГП, мM
Рис. 3. Зависимость начальной скорости реакции от концентрации субстрата НФГП: 1 — без лектинов; 2 — при добавлении спирта; 3 — в присутствии лектина A. brasilense Sp7; 4 — в присутствии лектина A. brasilense Sp7.2.3; 5 — при добавлении этанола и лектина A. brasilense Sp7; 6 — при добавлении этанола и лектина A. brasilense Sp7.2.3.; концентрация лектинов — 10 мкг/мл; концентрация этанола — 30%; pH 4,6
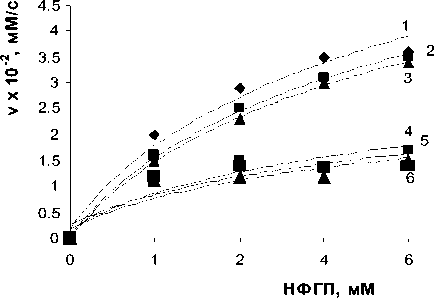
Рис. 4. Зависимость начальной скорости реакции от концентрации субстрата НФГП: 1 — без лектина; 2 — при добавлении лектина A. brasilense Sp7; 3 — лектин и 100 мМ NH 4 C1; 4 — лектин и 100 мМ KC1; 5 — лектин и 100 мМ NaCl; 6 — лектин и 100 мМ CaC1 2 ; концентрация в —глюкозидазы — 3,7 мг/мл; концентрация лектинов — 10 мкг/мл; 0,1 М ацетатный буфер, рН 4,6; 37°С.
Одним из главных факторов, ответственных за образование белок-белковых комплексов, являются электростатические взаимодействия. Они
____________Проблемы прикладной экологии могут не только ускорять формирование сложного белкового контакта, определять продолжительность жизни комплексов, но и определять правильное пространственное расположение контактирующих белков. Интенсивность взаимодействия между молекулами часто зависит от диэлектрической проницаемости среды.
Диэлектрическую проницаемость водных растворов можно понизить, добавляя различные смешивающиеся с водой органические растворители. Наиболее часто для этой цели используют этанол [13].
С целью экспериментальной оценки электростатического вклада во взаимодействие в—глюкозидазы и лектинов был исследован характер изменения кинетических параметров реакции при варьировании значений электропроводности реакционной среды. Поскольку взаимодействие лектинов с ферментом наблюдали по изменению кинетических параметров реакции гидролиза НФГП, предварительно была проведена оценка влияния органического растворителя на кинетические параметры собственно реакции гидролиза, катализируемой в—глюкозидазой.
Показано, что спирт оказывает активирующее действие на реакцию гидролиза. На рис. 3 представлены кривые зависимости скорости реакции гидролиза НФГП от концентрации субстрата в присутствии этанола для обоих штаммов.
Уменьшение диэлектрической проницаемости реакционной среды при внесении этанола до 30 % существенно активирует реакцию гидролиза НФГП (рис. 3, кривая 2).
Можно было ожидать, что активация этанолом при добавлении его к реакционной смеси в условиях ингибирования лектинами экспериментально будет проявляться в ослаблении ингибирования. Однако, как показывают данные, полученные в ходе исследования, ингибирование как лектином родительского, так и мутантного штаммов лектинами в присутствии этанола еще более усиливается. При этом кажущаяся константа Михаэлиса, равная 0,46±0,05 тМ в условиях ингибирования лектином родительского штамма и 0,5±0,02 тМ для лектина мутантного штамма, уменьшается в случае ингибирования лектинами и при добавлении спирта и составляет 0,08±0,01 тМ — для A.
Известия Самарского научного центра Российской академии наук. 2009. Т. 11, № 1
brasilense Sp7 и 0,06±0,01 mM — для A. brasilense Sp7.2.3. Вероятно, обнаруженное усиление ингибирования в присутствии этанола не может быть объяснено действием этанола на реакцию, катализируемую р-глюкозидазой. Видимо, добавление органического растворителя приводит к большей экспонированности гидрофобных областей лектинов и фермента и, возможно, вследствие усиления гидрофобного взаимодействия — к более прочному связыванию лектинами фермента. Это позволяет считать, что связывание лектинов с ферментом обусловлено не столько электростатическими взаимодействиями аминокислотных остатков, сколько взаимодействием гидрофобных областей.
Известно, что в белок-белковые взаимодействия вносят вклад электростатические взаимодействия типа солевых мостиков (отрицательно заряженные карбоксильные группы и положительно заряженные группы аминокислот образуют ионные пары). Присутствие различных ионов может приводить к нейтрализации ионных групп, входящих в солевые мостики и воздействовать на взаимодействие фермента и ингибитора. Кроме того, ионы могут связываться с заряженными группами активного центра, что также может влиять на скорость каталитического процесса [13] .
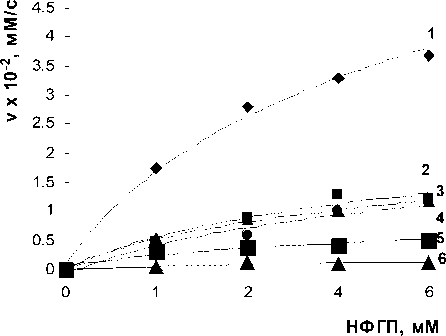
Рис. 5. Зависимость начальной скорости реакции от концентрации субстрата НФГП: 1 — без лектина; 2 —при добавлении лектина A. brasilense Sp7.2.3; 3 — лектин и 100 мМ NH4C1; 4 — лектин и 100 мМ CaC12; 5 — лектин и 100 мМ NaCl; 6 — лектин и 100 мМ КС1. Концентрация р -глюкозидазы — 3,7
мг/мл; концентрация лектинов — 10 мкг/мл; 0,1 М ацетатный буфер, рН 4,6; 37°С.
Было изучено влияние ионов Na+, К + , Са2+, NH4+ на ингибирование лектинами реакции гидролиза НФГП р -глюкозидазой. Необходимо отметить, что ионы оказывали неоднозначное влияние на собственно реакцию гидролиза НФГП р -глюкозидазой. NH 4 + не влиял на скорость реакции. Na+, К+, Са2+ оказывали ингибирующее действие. В случае с лектином A. brasilense Sp7, объединенное действие лектинов и NH 4 + также не влияло на ингибирующий эффект лектинов обоих штаммов. Na+, К+, Са2+ усиливали реакцию ингибирования для A. brasilense Sp7 (рис. 4).
Так как различия между ионами были незначительными, для получения кинетических характеристик был взят лишь вариант с Na+. Так, константа Михаэлиса, равная 0,46±0,05 тМ в условиях ингибирования лектином, при добавлении 100 тМ Na+ уменьшалась до 0,36±0,02 тМ. В случае с лектином A. brasilense Sp7.2.3 лишь Na+, К+ усиливали ингибирующий эффект, Са2+ не оказывал влияния на реакцию (рис. 5).
Так, константа Михаэлиса, равная 0,5±0,02 тМ в условиях ингибирования лектином, при добавлении 100 тМ Na+ уменьшалась до 0,4±0,01 тМ, т.е. уменьшение константы Михаэлиса было намного меньшим по сравнению с лектином родительского штамма. В обоих случаях уменьшение константы свидетельствует об увеличении сродства лектинов с ферментом, следствием чего и является усиление эффекта. Различное влияние солей на реакцию связано, видимо, с различными физико-химическими свойствами катионов. Таким образом, ингибирующий эффект лектинов азоспирилл обусловлен электростатическими и гидрофобными взаимодействиями аминокислотных остатков.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (грант НШ-6177.2006.4).
Список литературы Исследование взаимодействия лектинов азоспирилл с β-глюкозидазой сладкого миндаля
- Никитина В.Е., Пономарева Е.Г., Аленькина С.А.//Молекулярные основы взаимоотношений ассоциативных микроорганизмов с растениями. М.: Наука, 2005.
- Аленькина С.А., Петрова Л.П., Никитина В.Е. Получение и характеристика мутанта Azospirillum brasilense Sp7 по лектиновой активности//Микробиология. 1998. Т. 67. №. 6.
- Никитина В.Е., Аленькина С.А., Пономарева Е.Г., Савенкова Н.Н. Изучение роли лектинов клеточной поверхности азоспирилл во взаимодействии с корнями пшеницы//Микробиология. 1996. Т. 65, №. 2.
- Чернышева М.П., Аленькина С.А., Никитина В.Е., Игнатов В.В. Внеклеточные протеолитические ферменты штамма Azospirillum brasilense Sp7 и регулирование их активности гомологичным лектином//Прикл. биохимия и микробиология. 2005. Т. 41, № 4.
- Аленькина С.А., Паюсова О.А., Никитина В.Е. Сравнительное изучение влияния лектинов Azospirillum brasilense Sp7 и его мутанта на активность некоторых ферментов растительной клетки//Микробиология. 2004. Т. 73, № 6.
- Sadasivan L., Neyra C.A Flocculation in Azospirillum brasilense and Azospirillum lipoferum//J. Bacteriol. 1985. V. 163.
- Lis H., Sharon N. Soya bean agglutinin. Methods in Enzymology/Eds. Colowick S.P., Kaplan N.O. N.Y., London: Acad. Press, 1972. Part. B.
- Echdat Y., Ofek I., Yachow-Yan Y., Sharon N.,Mirelman D. Isolation of mannose-specific lectin from E.coli and its role in the adherence of the bacterial to epithelial cells//Biochem. Biophis. Res. Commun. 1978. V. 85.
- Сarbohydrate analysis/Ed. Chaplin M.E., Kennedy J.E. Oxford: IRL Рress. 1986.
- Linewiver H., Burk D. The Determination of Enzyme Dissociation Constants//J. Amer. Chem. Soc. 1934. V. 56.
- Корниш-Боуден Э. Основы математики для биохимиков. М.: Наука, 1983.
- Рокицкий П.Ф. Биологическая статистика. Минск: Вышейш. школа, 1973.
- Уэбб Л.. Ингибиторы ферментов и метаболизма. М.: Мир, 1966.
