История становления и современное распространение видов рода рогоз (Typha L.) Вятско-Камского Предуралья
 Вятско-Камского Предуралья")
Автор: Капитонова О.А., Дюкина Г.Р.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-4 т.11, 2009 года.
Бесплатный доступ
Показан характер современного распространения видов рода Typha на территории Вятско-Камского Предуралья. На основе представлений об историческом развитии и адаптациогенезе рогозов определены возможные причины редкой встречаемости одних и широкого распространения других таксонов. Установлены эколого-фитоценотические стратегии видов рогозов. Показано, что для всех видов базовой является R-стратегия, на основе которой возможно формирование промежуточных вариантов (CR-стратегии). Предварительная оценка сукцессионного статуса биоценозов с доминированием рогозов дает представление о них как о видах, выполняющих преимущественно функции пионеров в динамике прибрежно-водных экосистем.
Рогоз, макрофиты, эколого-фитоценотические стратегии, предуралье
Короткий адрес: https://sciup.org/148198397
IDR: 148198397 | УДК: 581.5(470.5)(045)
History of becoming and modern distribution of species of genus cattail (Typha L.) of Vyatka-Kama region
Character of modern distribution of species of genus Typha in territory Vyatka-Kama Region is shown. The reasons of rare occurrence of one species and a wide circulation of other species of cattails are certain. Are established the ecology-phytocenosis strategy of species of cattails. It is shown, that for all species R-strategy on the basis of which formation of intermediate variants (CR-strategy) is possible is base. The estimation the succession status of biocenosis with domination the cattails, pioneers carrying out mainly function in dynamics water ecosystem is given.
Текст научной статьи История становления и современное распространение видов рода рогоз (Typha L.) Вятско-Камского Предуралья
Общая филогенетическая реконструкция рогозов Евразии дана А.Н. Красновой [18]. Согласно ее представлениям, род Typha L. имеет четкие тропические корни. Его предковые меловые формы, про- изошедшие, вероятно, от гипотетических мезофитов тропического леса, перешедших вторично к условиям водной среды в составе своеобразной специализированной группы Helobiae (Болотнико-вые) [32], были связаны с периодически заболачивающимися окраинами водоемов. Центром происхождения и видового разнообразия рогозов, очевидно, следует считать обширные равнины Ирано-Туранской области, где усыхающий древний Тетис оставлял после себя многочисленные небольшие водоемы, мелководья которых и служили ареной эволюции рода, представленного в пределах этой территории большим таксономическим разнообразием. В палеогене происходит экологическая экспансия рогозов, и, видимо, к этому времени уже были сформированы основные секции рода. Наиболее древние из рогозов, сохранившие примитивные черты строения (например, T. elephantina Roxh.), осваивают экологические ниши в условиях тропического и субтропического климата. Другая, эволюционно более продвинутая часть рогозов расширяет область своего распространения в северном и восточном направлениях.
На территории Северной Евразии достоверные находки рогозов известны уже из олигоцена Западной Сибири [6], Казахстана, Малой Азии и Закавказья [19]. Рогозы характерны и для олигоценовых и неогеновых отложений европейской части континента [5, 37, 44], где они входили в состав палеокомплексов гигрофильного высокотравья. Подобные сообщества с участием рогозов были распространены, очевидно, и на территории современного ВКП. Ухудшение климатической обстановки в конце плиоцена – в плейстоцене вызвало перестройку древней арктотретичной флоры Евразии, что привело к захвату освобождающихся экологических ниш видами, обладавшими широкими пре-адаптационными возможностями. Территория ВКП оказалась в зоне перигляциального климата [2, 3], в пределах которой место третичных видов довольно быстро было занято менее теплолюбивыми формами [16]. В условиях климатической нестабильности плейстоцена происходит дифференциация рогозов по степени устойчивости к значительным температурным колебаниям и в целом к более холодному климату. Виды, выработавшие такую устойчивость, смогли существенно расширить свой ареал в плей-стоцен-голоценовое время. К ним, в частности, относятся наиболее широко распространенные в настоящее время рогозы широколистный (T. latifolia L. s.l.) и узколистный (T. angustifolia L.). Не случайно, уже в раннее послеледниковье пыльца этих растений в значительном количестве фиксируется даже на широте северного полярного круга [10], а южнее, в том числе на территории ВКП, эти виды входили в число доминантов растительного покрова обводненных пространств [31]. В большом количестве пыльца и плоды T. latifolia обнаружены также в отложениях позднеледниковья лесной зоны Восточно-Европейской равнины [27].
Стратегия другой части видов рогозов (например, из секции Engleria (Leonova) N. Tzvel. – T. laxmannii Lepechin и близкие к нему виды) заключалась в захвате экологического пространства аридных и субаридных областей; они эволюционировали в направлении выработки адаптаций к временному пересыханию местообитаний и увеличению концентрации солей в субстрате.
Таким образом, современное таксономическое разнообразие рогозов, насчитывающее по разным данным от 15 [20, 43] до 23 [18] видов, является следствием адаптивной радиации рассматриваемой группы, обладающей значительными преадаптаци-онными возможностями и высокими темпами эволюции специализированных организмов.
Выработанные рогозами адаптации к обитанию в экстремальных условиях значительно увеличили их шансы на выживание в настоящее время, когда типичные для рогозов местообитания испытывают сильнейший антропогенный пресс. Исследования, проведенные нами в пределах ВКП, показали, что сообщества именно этой группы растений зачастую выступают в качестве пионерных при зарастании антропогенно трансформированных и искусственных экотопов.
Гибридизация. Климатические колебания и связанная с ними нестабильность гидрорежима мест обитания рогозов, вероятно, уже в плейстоцене, а может быть и раньше, приводили к перекрыванию ареалов разных видов этого рода, результатом чего могла быть спонтанная гибридизация, весьма характерная для рогозов и в настоящее время [18, 22]. По мнению Н.Н. Цвелева [41], массовое формирование гибридов чаще всего происходит в критических ситуациях, например, во время направленных климатических изменений, когда один из видов, более приспособленный к новым условиям, наступает на позиции менее приспособленного вида. При этом процесс гибридизации рассматривается как возможный путь деспециализации таксонов гибридного происхождения [4, 40, 41], позволяющий изменить пределы толерантности гибридов, что представляется чрезвычайно важным для рогозов – растений, достигших определенного уровня структурной специализации своих органов. В результате гибридные
А
Б
В
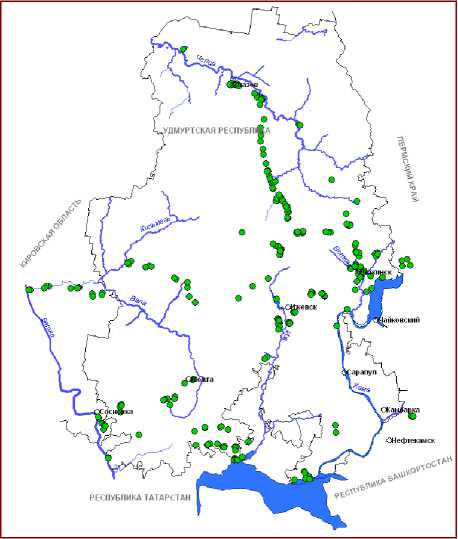
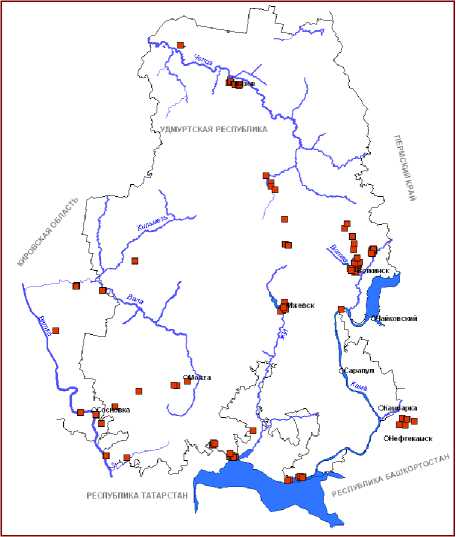
Рис. 1. Распространение видов рода Typha L. на территории Вятско-Камского Предуралья: А – T. latifolia , Б – T. intermedia , В – T. shuttleworthii , Г – T. laxmannii
Г
особи получают новые возможности для захвата экологического пространства, в том числе в направлении освоения местообитаний с экстремальными условиями. Дальнейшая эволюция гибридов может быть отличной от исторического развития родительских видов, что, в конце концов, может привести к формированию новых гибридогенных таксонов [38].
Таксономическое разнообразие и эколого-фитоценотические стратегии рогозов ВКП.
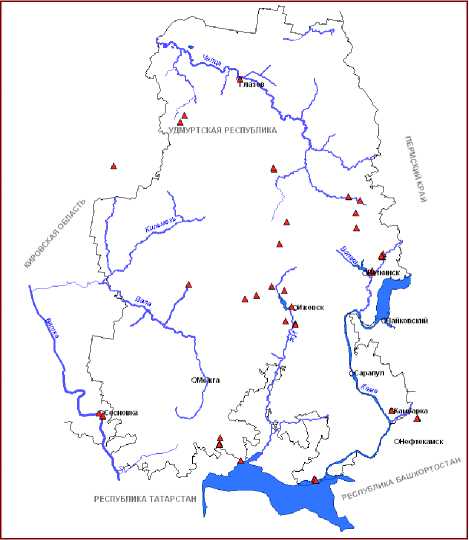
В настоящее время на территории ВКП нами установлено произрастание 8 видов рогозов . Из них к наиболее обычным и широко распространенным относятся T. latifolia L. и T. angustifolia L. Остальные относятся к спорадически встречающимся или редким таксонам (рис. 1,2).
Рогоз широколистный (T. latifolia) чаще всего заселяет трансформированные и искусственные биотопы, значительно реже встречаясь в первичных сообществах, проявляя тем самым признаки синантропного вида. Это достаточно вариабельный в экологическом отношении таксон, выдерживающий широкий диапазон изменения значений абио-
тических факторов.
Д
Ж
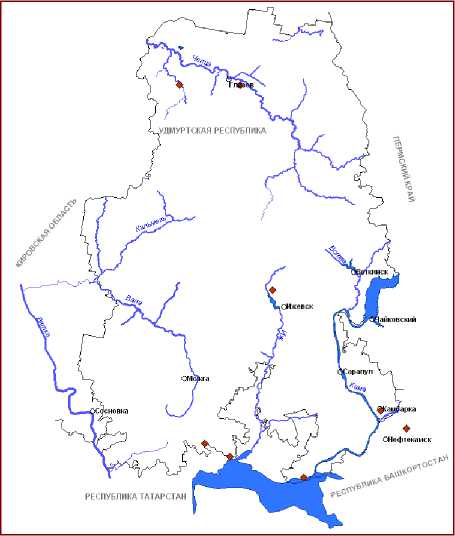
Рис.2. Распространение видов рода Typha L. на территории Вятско-Камского Предуралья: Д – T. angustifolia , Е - T. incana , Ж - T. x glauca , З - T. x smirnovii .
Е
В оптимальных условиях он обладает высокой конкурентоспособностью, что наглядно проявляется при совместном обитании нескольких видов этого рода: рогоз широколистный почти всегда занимает наиболее благоприятные для роста и развития участки с глубиной воды 10-30 (50) см, вытесняя в более глубоководную зону T. angustifolia , а в прибрежное мелководье и на берег – рогозы промежуточный ( T. intermedia Schur.) и Лаксмана ( T. laxmannii Lepechin). Конкурентоспособность T.
latifolia достигается за счет большой надземной и подземной биомассы, которую он способен формировать на любых типах субстрата. По нашим данным [7], сырая надземная биомасса этого вида в ВКП варьирует в пределах от 3,09 до 7,05 кг/м2, причем наибольшие значения этого показателя зарегистрированы на грунтах со значительным содержанием илистых фракций. Более глубоководные местообитания в целом оказываются для рогоза широколистного не благоприятными, и он не выдерживает конкуренции с рогозом узколистным [21]. Отмеченные характерные особенности T. latifolia дают основания рассматривать его как вид с вторичной виолент-эксплерентной эколого-фитоценотической стратегией (CR-стратег).
Историческое развитие T. angustifolia происходило, по-видимому, на весьма глубоководных участках со стабильным уровнем воды. Именно такие местообитания в естественных и искусственных водоемах являются в настоящее время наиболее характерными для данного вида, обычно уже не способного к возобновлению после полного, хотя и временного, высыхания грунтов [21]. Этот вид является обычным на мелководьях водохранилищ, где он формирует одновидовые либо с участием других гидрофильных растений заросли. Довольно обычен этот вид также и на нарушенных местообитаниях, в том числе испытывающих антропогенное загрязнение (например, придорожные обводненные местообитания), где он зачастую не проявляет признаков угнетения и имеет удовлетворительное возобновление. Таким образом, наряду с эксплерент-ными свойствами для данного вида характерны и выраженные виолентные признаки, связанные, прежде всего, с абсолютным доминированием в условиях благоприятного гидрорежима со стабильным уровнем воды, что позволяет отнести этот вид к вторичной CR стратегии.
Яркими примерами предпочтения рудеральных местообитаний на рассматриваемой территории можно считать рогозы промежуточный ( T. intermedia ), сизый ( T. x glauca Godron) и Смирнова ( T. x smirnovii E. Mavrodiev). Первый из них чаще всего встречается в разнообразных вторичных местообитаниях – в придорожных лужах, на зарастающих обводненных и увлажненных колеях, заболоченных нарушенных поймах, часто совместно с рогозом широколистным, которым вытесняется на периферийные участки сообществ. Изредка рогоз промежуточный способен образовывать и собственные небольшие заросли. По нашим наблюдениям, при антропогенном нарушении растительного покрова на переувлажненных местообитаниях, в особенности на грунтах легкого механического состава, на самых ранних стадиях сукцессионной серии одним из первых появляется именно рогоз промежуточный, который в дальнейшем обычно замещается другими, более конкурентоспособными видами последующих стадий сукцессии. Таким образом, T. intermedia можно охарактеризовать как ценофобный не конкурентоспособный вид преимущественно с эксплерентной эколого-фитоценотической стратегией (R-стратег).
Все известные к настоящему времени местонахождения T. x glauca (T. latifolia x T. angustifolia) и T. x smirnovii (T. latifolia x T. laxmannii) также приурочены к трансформированным либо искусственным биотопам, что в целом согласуется с литературными данными [23, 24], причем чаще всего эти нотовиды также обитают в совместных ценозах с рогозом широколистным.
Очевидно, спонтанная гибридизация T. angustifolia и T. latifolia происходила и раньше, хотя имеются данные, указывающие на ограниченность этого явления [45]. В настоящее же время широкие возможности для этого процесса открыты в связи с нарушением среды обитания рогозов. В литературе для территории России рогоз сизый указывается как редкий таксон [18, 29, 30]. В пределах же ВКП этот гибридогенный вид известен из более чем 10 местонахождений (2, Ж).
Появление и дальнейшее распространение на территории ВКП второго гибридогенного таксона – T. x smirnovii , возможно, следует рассматривать как одно из следствий общепланетарного процесса потепления климата, одним из проявлений которого в рассматриваемом регионе является смещение к северу зональных и подзональных границ [15], что приводит к изменению ареалов и экспансии на север видов южного распространения. Последнее имеет место и в пределах ВКП, что связано с существованием здесь миграционного пути в виде широкой долины р. Камы. Одним из видов, использующих эту миграционную трассу для расселения в северном направлении, является T. laxmannii , который в настоящее время уже вполне естественно входит в сообщества прибрежно-водных растений поймы р. Камы в ее среднем течении в пределах Удмуртской Республики [12]. Вне поймы Камы находки рогоза Лаксмана в рассматриваемом регионе связаны преимущественно с придорожными обводненными местообитаниями и другими вторичными биотопами [8, 11, 33], что демонстрирует его характерные особенности адвентивного вида. Широкое расселение этого вида отмечено и другими авторами: в качестве заносного T. laxmannii указан для территории Ленинградской [39], Тверской [28] и Рязанской [42] областей, Республики Татарстан [1], окрестностей г. Петрозаводска [17]. Перекрывание ареалов T. latifolia и T. laxmannii способствует гибридизации этих видов, а широкий спектр трансформированных и искусственных экотопов позволил закрепиться гибридным растениям, дальнейшее размножение которых осуществляется в основном вегетативным способом.
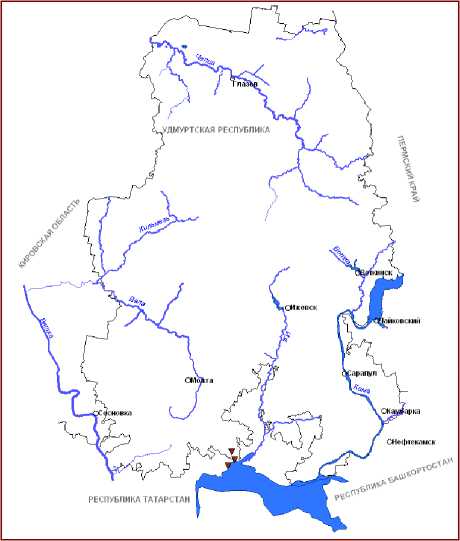
Новостью для рассматриваемого региона является находка T. shuttleworthii W.D.J. Koch et Sond. – вида западно-европейского распространения, редкого в пределах почти всего видового ареала [25]. Он относится к уязвимым видам в Германии, Швейцарии и Австрии, редким – в Болгарии, подвергающимся опасности – в Словакии и государстве Лихтенштейн, исчезнувшим таксонам – в Чехии [46, 47]. В России этот рогоз считается достаточно обычным в ряде западных регионов и приводится для Московской и Калужской областей, известен также с территории Краснодарского края, некоторых районов Предкавказья [22]. На территории
ВКП рогоз Шутлеворта был обнаружен нами в трех пунктах в пределах республик Удмуртия и Татарстан (рис. 1, В). Произрастание этого вида в ВКП, возможно, также следует рассматривать в контексте глобальных изменений климата.
В пределах основного ареала экотопологически связанный с естественными обводненными местообитаниями, на территории ВКП рогоз Шутлеворта встречается на вторичных местообитаниях, представленных мелководьями искусственных водоемов (небольших прудов). Тем не менее, по нашему мнению, в отличие от всех остальных рогозов региона, рассматриваемых нами в качестве синантропных видов [9], T. shuttleworthii не может быть отнесен к синантропному элементу флоры ВКП. Возможно, в прошлом (в третичное время) ареал T. shuttleworthii был значительно шире и охватывал также территорию современного Среднего Преду-ралья. Однако климатическая нестабильность плейстоцен-постплейстоценового времени, экспансия на север вслед за отступающим ледником более конкурентоспособных видов сходного экологического предпочтения, включая и виды рода Typha , способствовали сокращению области распространения рогоза Шутлеворта со смещением ее в более западные районы с океаническим климатом. Этот процесс обычно бывает связан с возникновением длительное время существующих анклавов, что, вероятно, и произошло в отношении T. shuttleworthii на территории современного ВКП. Таким образом, в рассматриваемом регионе данный вид, по нашему предположению, следует рассматривать в качестве реликтового таксона с преимущественно виолентной с долей эксплерентности эколого-фитоценотической стратегией (CR-стратег). На современном этапе состоянию популяций рогоза Шутлеворта может угрожать хозяйственная деятельность человека, связанная с нарушением, трансформацией и загрязнением типичных мест его обитания, следствием чего может быть исчезновение популяций или их генетическая ассимиляция с близкородственными видами.
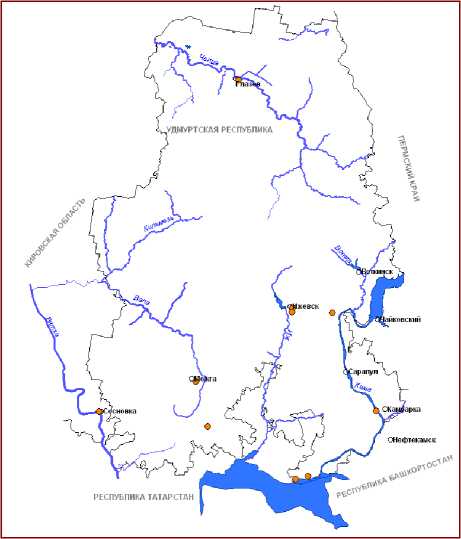
Еще одним видом, тяготеющим к нарушенным биотопам, является недавно описанный нами рогоз седой – T. incana Kapitonova et Dyukina [13], способный образовывать большие заросли в придорожных лужах и на мелководьях прудов. Этот вид, как и рогоз промежуточный, осваивает обводненные экотопы на начальных стадиях сукцессионной серии, впоследствии замещаясь другими видами. Рогоз седой на территории ВКП имеет в целом невысокую конкурентоспособность, что характеризует его как вид с преимущественно эксплерентной эколого-фитоценотической стратегией. Так, одна из популяций T. incana , занимавшая первоначально площадь более 500 м2, спустя 2 года после начала наблюдений практически была вытеснена ольхой черной ( Alnus glutinosa ), образовавшей плотные сомкнутые заросли.
ЗАКЛЮЧЕНИЕ
Анализ экологических и фитоценотических предпочтений известных на территории ВКП видов рогозов позволяет говорить об их в целом экспле-рентной эколого-фитоценотической стратегии (R-стратеги). У ряда видов на ее базе выработались промежуточные варианты, отражающие приспособительные реакции на условия, в которых происходило их становление. Так, в силу особенностей биологии и экологии T. latifolia , T. shuttleworthii и T. angustifolia могут быть отнесены к видам с преимущественно виолент-эксплерентным типом стратегии (CR-стратеги). Установленные эколого-фитоценотические стратегии указывают на то, что популяции рогозов входят главным образом в сообщества начальных стадий сукцессионных серий. Это дает им широкие возможности к заселению экстремальных условий с постоянным или периодическим влиянием экзогенных факторов, включая антропогенные (разнообразные вторичные переувлажненные и обводненные местообитания, мелководья водоемов с переменным гидрорежимом, нарушенные поймы, выработанные торфяники, карьеры и др.). В условиях стабильного режима функционирования экосистем (мелководья естественных и искусственных водоемов и водотоков с постоянным уровнем воды в течение вегетационного периода, ненарушенные болота) рогозы способны длительное время сохранять свои позиции доминирующего (или субдоминирующего) вида серийных сообществ, развивающихся под преимущественным влиянием эндогенных факторов.
Авторы признательны д-ру биол. наук, проф. О.В. Смирновой (ЦПЭПЛ РАН, г. Москва), д-ру биол. наук, проф. В.Г. Папченкову (ИБВВ РАН, п. Борок Ярославской обл.), д-ру биол. наук, проф. В.В. Туганаеву (УдГУ, г. Ижевск) за обсуждение представленных в статье материалов, а также канд. биол. наук А.Н. Пузыреву (УдГУ, г. Ижевск) за предоставление устных сведений о распространение T. laxmannii на территории Удмуртии.
Список литературы История становления и современное распространение видов рода рогоз (Typha L.) Вятско-Камского Предуралья
- Бакин О.В., Ситников А.П. Новые и редкие в Татарстане виды сосудистых растений//Бот. журн. -2005. -Т. 90. № 1. -С. 66-71.
- Бутаков Г.П. Граница максимального оледенения в Вятско-Камском регионе//Физико-географические основы развития и размещения производительных сил Нечерноземного Урала: Межвуз. сб. науч. тр. -Пермь: Пермский ун-т, 1981. -С. 102-110.
- Бутаков Г.П. Плейстоценовый перегляциал на востоке Русской равнины. Казань: Изд-во Казан. ун-та, 1986. -144 с.
- Грант В. Эволюция организмов. -М.: Мир, 1980. -253 с.
- Дорофеев П.И. О третичной флоре Белоруссии//Бот. журн. -1960. -Т. 45. № 10. -С. 1418-1434.
- Дорофеев П.И. Третичные флоры Западной Сибири. -М.-Л.: Изд-во АН СССР, 1963. -346 с.
- Дюкина Г.Р. Изменчивость структурно-функциональных показателей популяций рогозов (Typha L.) в зависимости от влияния эдафических факторов//Современное состояние и пути развития популяционной биологии: Матер. Х Всерос. популяционного семинара. -Ижевск: КнигоГрад, 2008. -С. 254-256.
- Дюкина Г.Р., Капитонова О.А. Род Typha L. в Удмуртии: таксономический состав, распространение, экология//Вестник Удм. ун-та. -2005. -№10. -С. 41-50.
- Дюкина Г.Р., Капитонова О.А. Рогозы как синантропный компонент флоры//Адвентивная и синантропная флора России и сопредельных стран: состояние и перспективы: Матер. III Международ. науч. конф. -Ижевск, 2006. -С. 41-42.
- Елина Г.А., Лукашов А.Д., Юрковская Т.К. Позднеледниковье и голоцен Восточной Фенноскандии (палеорастительность и палеогеография). -Петрозаводск: Карельский научный центр РАН, 2000. -242 с.
- Капитонова О.А. О распространении рогоза Лаксмана в Удмуртии//Биология внутренних вод: проблемы экологии и биоразнообразия: Тез. докл. XII Международной конф. молодых ученых. -Борок, 2002. -С. 7-8.
- Капитонова О.А., Дюкина Г.Р. О малоизвестных видах рогозов (Typha L.) во флоре Вятско-Камского междуречья//Материалы VI Всероссийской школы-конференции по водным макрофитам «Гидроботаника-2005». -Рыбинск: ОАО «Рыбинский Дом печати», 2006. -С. 246-266.
- Капитонова О.А., Дюкина Г.Р. Новый вид Typha (Typhaceae) из Удмуртии//Бот. журн. -2008. -Т. 93, № 7. -С. 1132-1134.
- Катанская В.М. Высшая водная растительность континентальных водоемов. -Л.: Наука, 1981. -187 с.
- Коломыц Э.Г., Розенберг Г.С. Палеопрогнозная концепция в региональной экологии (на примере Волжского бассейна)//Успехи совр. биол. -2004. -Т. 124, № 5. -С. 403-418.
- Колчанов В.И., Жуйкова И.А., Пахомов М.М., Прокашев А.М. Геологическое прошлое//Энциклопедия Земли Вятской. Т. 7. Природа. -Киров: Вятка, 1997. -С. 58-79.
- Кравченко А.В. Конспект флоры Карелии. -Петрозавдск: Карельский научный центр РАН, 2007. -403 с.
- Краснова А.Н. Структура гидрофильной флоры техногенно трансформированных водоёмов Северо-Двинской водной системы. -Рыбинск: ОАО «Рыбинский Дом печати», 1999. -200 с.
- Криштофович А.Н., Палибин И.В., Шапаренко К.К., Ярмоленко А.В., Байковская Т.Н., Ильинская И.А. Олигоценовая флора горы Ашутас в Казахстане//Палеоботаника. Вып. I/Под ред. А.Н. Криштофовича. -М.-Л.: Изд-во АН СССР, 1956. -180 с.
- Леонова Т.Г. Порядок рогозовые (Typhales)//Жизнь растений: В 6-ти т. Т. 6. Цветковые растения/Под ред. А.Л. Тахтаджяна. -М.: Просвещение, 1982. -С. 461-466.
- Мавродиев Е.В. Рогоз узколистный//Биологическая флора Московской области. Вып. 13/Под ред. В.Н. Павлова, В.Н. Тихомирова. М.: Изд-во Полиэкс, 1997. С. 4-29.
- Мавродиев Е.В. Морфолого-биологические особенности и изменчивость рогозов (Typha L.) России: Автореф. дис. … канд. биол. наук. М., 1999. 19 с.
- Мавродиев Е.В. Typha ´ smirnovii E. Mavrodiev (T. latifolia L. s. str. × T. laxmannii Lepechin) и некоторые другие гибридные рогозы с территории юго-востока России//Бюлл. МОИП. Отд. Биол. 2000. Т. 105. Вып. 4. С. 65-69.
- Мавродиев Е.В., Алексеев Ю.Е. О диагностике и систематическом положении Typha ? glauca Godron (Typha angustifolia L. ? T. latifolia L.)//Бюлл. МОИП. Отд. Биол. 1998. Т. 103. Вып. 6. С. 51-54.
- Макрофиты -индикаторы изменения природной среды/Под ред. С. Гейны, К.М. Сытника. Киев: Наукова думка, 1993. 434 с.
- Мейен С.В. Основы палеоботаники. М.: Недра, 1987. 403 с.
- Новенко Е.Ю., Зюганова И.С., Козлов Д.Н. Эволюция растительного покрова в позднем плейстоцене на территории Центрально-лесного заповедника//Известия РАН. Сер. Географ. 2008. № 1. С. 87-99.
- Нотов А.А., Шубинская Н.В., Маркелова Н.Р., Плетнев Д.М., Спирина У.Н. Новые и редкие адвентивные растения Тверской области//Бюл. МОИП. Отд. биол. 2002. Т. 107. Вып. 2. С. 47-48.
- Папченков В.Г. О новых и редких видах флоры Татарстана//Бот. журн. 1993. Т. 78. № 9. С. 73-79.
- Папченков В.Г. Растительный покров водоемов и водотоков Среднего Поволжья: Монография. Ярославль: ЦМП МУБиНТ, 2001. 200 с.
- Прокашев А.М., Жуйкова И.А., Пахомов М.М. История почвенно-растительного покрова Вятско-Камского края в послеледниковье. Киров, 2003. 143 с.
- Проханов Я.И. Класс однодольных, его таксономическая обоснованность и положение в системе сосудистых растений//Проблемы филогении высших растений. М: Наука, 1974. С. 75-103.
- Пузырев А.Н. О находках адвентивных видов растений на шоссейных дорогах Удмуртии//Вестн. Удм. ун-та. Биология. 2006. № 10. С. 25-36.
- Тахтаджян А.Л. Вопросы эволюционной морфологии растений. Л.: Изд-во Ленинград. ун-та, 1954а. 215 с.
- Тахтаджян А.Л. Происхождение покрытосеменных растений. М.: Сов. наука, 1954б. 96 с.
- Тахтаджян А.Л. Система и филогения цветковых растений. М., Л.: Наука, 1966. 611 с.
- Федченко Б.А. Семейство Рогозовые//Флора СССР. Т. 1. М.-Л.: АН СССР, 1934. С. 209-216.
- Цвелёв Н.Н. О значении гибридизационных процессов в эволюции злаков (Poaceae)//История флоры и растительности Евразии. Л.: Наука, Ленингр. отд., 1972. С. 5-16.
- Цвелев Н.Н. Определитель сосудистых растений Северо-Западной России (Ленинградская, Псковская и Новгородская области). СПб.: Изд-во СПХФА, 2000. 781 с.
- Цвелёв Н.Н. Гибридизация как один из факторов увеличения биологического разнообразия и геномный критерий родов у высших растений//Проблемы теоретической морфологии и эволюции высших растений: Сб. избранных тр./Под ред. Д.В. Гельтмана. М.; СПб.: Товарищество научных изданий КМК, 2005а. С. 141-148.
- Цвелёв Н.Н. О значении гибридизации в эволюции высших растений//Проблемы теоретической морфологии и эволюции высших растений: Сб. избранных тр./Под ред. Д.В. Гельтмана. М.; СПб.: Товарищество научных изданий КМК, 2005б. С. 149-154.
- Щербаков А.В., Девятов А.Г., Барзионова Т.В. Находки редких видов водных сосудистых растений на востоке Рязанской области//Бюл. МОИП. Отд. Биол. 2004. Т. 109. Вып. 3. С. 77-78.
- Casper S.J., Krausch H.-D. Pteridophyta und Anthophyta//Suwasserflora von Mitteleuropa. Jena: Gustav Fischer, 1980. V. 23. 403 s.
- Graebner P. Typhaceae//Das Pflanzenreich. Leipzig, 1900. H. 2 (IV.8). 18 s.
- Kuehn M. Marcinko, Minor J.E., White B.N. An examination of hybridization between the cattail species Typha latifolia and Typha angustifolia using random amplified polymorphic DNA and chloroplast DNA markers//Molecular Ecology. V. 8: 12. 2002. P. 1981-1990.
- Typha shuttleworthii//European Environment Agency. URL: http://eunis.eea.europa.eu/(дата обращения: 23.11.2006).
- Uhrin S., Bača F. A new locality of Typha shuttlewothii in Slovakia//Biologia/Bratislava. 60/1. 2005. S. 105.
