Из опыта преподавания. III. Кристаллография икосаэдрических вирусов

Автор: Войтеховский Ю.Л.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 4 (304), 2020 года.
Бесплатный доступ
Статья посвящена морфологической систематике икосаэдрических вирусов. Показано, что фиксация капсидов триангуляционными числами не различает изомеров. Описание капсидов точечными группами симметрии (- 5 - 3m и 532 ) существенно детализировано. В их многообразии выделены гомологические серии, связанные попарно. В пределах серии капсиды связаны преобразованием подобия. Серии связаны переходом к дуальным полиэдрам. Определены капсиды-генераторы, не сводимые к более простым формам и порождающие все разнообразие икосаэдрических капсидов.
Икосаэдрические вирусы, морфология капсида, точечная группа симметрии, триангуляционное число, гомологические серии, капсиды-генераторы, преобразование подобия, дуальные полиэдры
Короткий адрес: https://sciup.org/149128752
IDR: 149128752 | УДК: 548 | DOI: 10.19110/geov.2020.4.6
From teaching experience. III. Crystallography of icosahedral viruses
The article is devoted to the morphological systematics of icosahedral viruses. It is shown that fixation of capsids by the triangulation numbers does not distinguish the isomers. Description of capsids by symmetry point groups (- 5 - 3m and 532 ) is greatly detailed. In their diversity, homologous series connected in pairs are distinguished. Within the series, capsids are linked by a similarity transformation. The series are connected by the transition to the dual polyhedra. Capsid generators that are not reducible to simpler forms and produce the entire variety of icosahedral capsids are defined.
Текст научной статьи Из опыта преподавания. III. Кристаллография икосаэдрических вирусов
Давно замечено, что природа рациональна. Структуры, оптимальные в том или ином физическом контексте, копируются в минеральных и живых объектах. Плоскости, простые и винтовые оси симметрии в организации минералов, животных и растений буквально бросаются в глаза. По меткому выражению Н. В. Белова, «осью 5-го порядка жизнь защищается от кристаллизации». Вокруг нас идет согласование эволюционных стратегий. Неудивительно, что в поле зрения кристаллографии попадает все больше структур, к описанию которых применимы математический язык теории симметрии и физические методы структурного анализа. Представляется правильным включать рассмотрение таких примеров в университетский лекционный курс.
Кристаллография живого вещества впервые и мощно заявила о себе расшифровкой структур ДНК и РНК Дж. Уотсоном и Ф. Криком в 1953 г. (Нобелевская премия 1962 г.). С тех пор теоретическим и приборными анализами охвачены разные живые объекты. В том числе нами показано широкое распространение в минеральной и живой природе полиэдрических структур типа фуллеренов, в которых разрешены только 5-угольные (ровно 12) и 6-угольные (все остальные) грани [5]. При этом в самых симметричных фуллеренах легко узнаваем додекаэдрический мотив, ведь простейший фуллерен — именно додекаэдр, а самый стабильный фуллерен Сбо — усечённый икосаэдр. В скелетах радиолярий встречены оба мотива (Circogonia dodecahedra Hackel и Circogonia icosahedra Hackel).
Но наиболее ярко они выражены в капсидах икосаэдрических (сферических) вирусов (рис. 1). Тот факт, что додекаэдр и икосаэдр геометрически дуальны друг другу, позволяет легко перейти от одного к другому и дать классификацию и номенклатуру для всех структур на их основе. «Номенклатура и классификация вирусов — трудная область вирусологии... Группировка может быть основана на морфологических и физиологических критериях. Идеал, к которому мы стремимся — это создать такую классификацию, которая отражала бы эволюционные и филогенетические взаимоотношения организмов, а также обеспечивала бы удобную и рациональную систему номенклатуры» [1, с. 18—23]. В части морфологии икосаэдрических вирусов задачи классификации и номенклатуры решаются в предлагаемой статье.
Принципы классификации и номенклатуры
Принципы строения капсидов икосаэдрических вирусов из капсомеров (субъединиц, сферических белковых глобул) и их классификация предложены ранее [3]. Всякий капсид (белковая оболочка, в которую упакована свернутая в клубок молекула ДНК или РНК) построен из 20 равносторонних треугольных мегаграней — фрагментов плоской сетки, образованной гло-
Гексоны (всего 20)
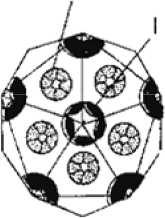
Пентоны (всего 12)
Полые шипы на вершинах икосаэдра
путем протеолиза
Фибриллярные отростки
Капсид содержит
12 пеятонов i по одному на каждую вершину) и 240 гексонов
Неустановленное число капсомеров на гранях
Удаление наружного капсида
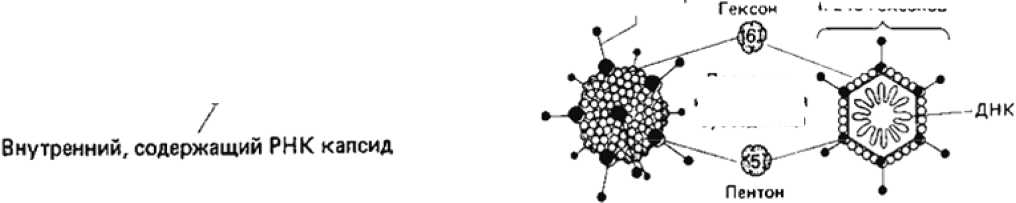
РЕ08ИРУС (55-60 нм, вирус животных}, показаны наружный и внутренний капсиды
АДЕНОВИРУС Поперечный срез аденовируса,
160—90 нм, вирус животных) гюказывающии расположение ДНК
Протомер, или белковая субъединица
Рис. 1. Строение икосаэдрических вирусов [2]
Fig. 1. The structure of icosahedral viruses [2]
булами по принципу плотнейшей шаровой упаковки. Линии глобул не всегда совпадают с рёбрами икосаэдра, что и создавало проблему поиска общей формулы для их числа в капсиде.
В статье [3] анонсирована теорема, перечислившая разрешённые для икосаэдрических капсидов «триангуляционные числа» Т = Pf2, где Р = h2 + hk + k2, h > k > 0 — любые пары целых чисел без общих делителей (при этом h > 0, т. к. h = 0 влечет k = 0, что противоречит физическому смыслу триангуляционного числа как площади грани капсида), f = 1, 2, 3... Доказательство впервые приведено в статье [4, с. 1124—1125]. По сути, h и k — координаты конца базального ребра мегаграни в косоугольной (60°) декартовой системе координат, согласованной с плотнейшей упаковкой глобул (рис. 2).
В основу описания икосаэдрических капсидов сегодня положены именно триангуляционные числа Т (табл. 1). В этом есть известное удобство. Число глобул в капсиде: М = 10Т + 2. Они образуют морфологические субъединицы: 12 пентамеров и 10(Т — 1) гексамеров. При этом Т = (m — 1)2, где m — число глобул на ребре мегаграни, т. е. если линия глобул и ребро икосаэдра сонаправлены (верхняя строка табл. 1, класс (f, 0), см. далее) [6]. Кроме того, 20Т — число элементарных треугольных граней у любого капсида. Для многих вирусов эти характеристики известны: бактериофаг фХ174: Т = 1, M = 12, m = 2, (h, k) = (1, 0); вирус жёлтой мозаики турнепса: Т = 3, M = 32, (h, k) = (1, 1); вирус полиомы: Т = 4, M = 42, m = 3, (h, k) = (2, 0); паповавирус: Т = 7, М = 72, (h, k) = (2, 1); реовирус: Т = 9, M = 92, m = 4,
(h, k) = (3, 0); ротавирус: Т = 13, М = 132, (h, k) = (3,1); вирусы герпеса и ветряной оспы: Т = 16, M = 162, m = 5, (h, k) = (4, 0); аденовирус и вирус собачьего гепатита: Т = 25, M = 252, m = 6, (h, k) = (5, 0); радужный вирус: Т = 81, M = 812, m = 10, (h, k) = (9, 0). Анализ таблицы обнаруживает изомеры — вирусы с одинаковым Т. Для Т = 49 их можно различить по симметрии: капсид (h, k) = (7, 0) имеет точечную группу симметрии (т. г. с.) — 5 - 3m , капсид (5, 3) — т. г. с. 532 . * Но для Т = 91 оба капсида (9, 1) и (6, 5) имеют т. г. с. 532 . Символ (h, k) фиксирует строение любого капсида однозначно. Именно его следует положить в основу номенклатуры.
Гомологические сериив классах —5—Зм и 532
В статье [3, с. 15, табл. 1] предложено деление икоса-эдрических капсидов на три класса. Первый (верхняя строка табл. 1) порождается простейшим капсидом (1, 0) — икосаэдром, в вершинах которого расположены белковые глобулы. Остальные капсиды класса, все с т. г. с. – 5 – 3m , получаются увеличением грани икосаэдра с помощью коэффициентов подобия f = 2, 3, 4… при той же ориенти-
Таблица 1. Триангуляционные числа T Table 1. Triangulation numbers T
|
k\h |
1 |
1 2 1 |
1 3 1 |
1 4 1 |
1 5 | |
1 6 |
7 |
8 1 |
1 9 1 |
1 10 |
|
0 |
1 |
4 |
9 |
16 |
25 |
36 |
49 |
64 |
81 |
100 |
|
1 |
3 |
7 |
13 |
21 |
31 |
43 |
57 |
73 |
1 91 1 |
1 111 |
|
2 |
12 |
19 |
28 |
39 |
52 |
67 |
84 |
103 |
124 |
|
|
3 |
27 |
37 |
1 49 1 |
63 |
79 |
97 |
117 |
139 |
||
|
4 |
48 |
61 |
76 |
93 |
112 |
133 |
156 |
|||
|
5 |
75 |
1 91 1 |
109 |
129 |
151 |
175 |
||||
|
6 |
108 |
127 |
148 |
171 |
196 |
|||||
|
7 |
147 |
169 |
193 |
219 |
||||||
|
8 |
192 |
217 |
244 |
|||||||
|
9 |
243 |
271 |
||||||||
|
10 |
300 |

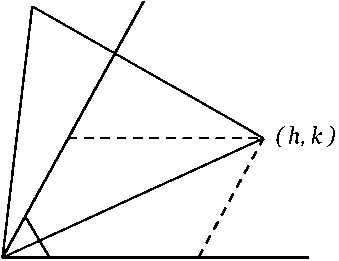
Рис. 2. Положение грани капсида (равносторонний треугольник) относительно линии глобул (горизонтальная ось h )
Fig. 2. The position of the capsid facet (equilateral triangle) relative to the line of globules (horizontal axis h )
ровке относительно плотнейшей упаковки глобул (рис. 2). Этот класс логично обозначить (f, 0). Судя по имеющимся публикациям (к сожалению, публикуются не первичные снимки высокого разрешения, а раскрашенные модели), короновирус SARS-CoV-2 (табл. 2, врезка) имеет капсид (2, 0), Т = 4, аналогичный вирусу полиомы. Это второй капсид в классе Т = f2, начинающемся с (1, 0) — бактериофага ϕX174. За SARS-CoV-2 следуют: (3, 0) — реовирус, (4, 0) — вирус герпеса и ветряной оспы, (5, 0) — аденовирус и вирус собачьего гепатита, … (9, 0) — радужный вирус.
Второй класс (диагональ табл. 1) порождается аналогичным преобразованием подобия из капсида (1, 1) — додекаэдра, над гранями которого надстроены пентагональные пирамиды. Его можно обозначить (f, f). Капсиды этого класса тоже имеют т. г. с. — 5-3m. Остальные капсиды (внутри треугольника в табл. 1) имеют только поворотные оси, то есть т. г. с. 532. Судя по тому, что они названы в статье [3] skew classes (скошенные, асимметричные классы), каждый символ (h, k) означает отдельный класс, в каждом — один капсид. Это не логично, так как преобразованием подобия любой капсид порождает бесконечную серию одинаково устроенных капсидов: (h, k) ^ (fh, fk)... Табл. 1 вся состоит из таких серий, начинающихся с (h, k), у которых h и k не имеют общих делителей.
Табл. 1 содержит и более интересные связи. Капсид (1, 1) получается из (1, 0) как усечённый икосаэдр из додекаэдра — дуальным переходом и отсечением всех вершин. Как это выразить алгебраически? На целочисленной решётке (рис. 2) рёбра (hb k 1 ) и (h2, k2) двух капсидов выражаются друг через друга линейно. Пусть искомое преобразование имеет вид: (h 1 , k 1 ) ^ (h2, k2) = (ah 1 +bk 1 , ch 1 +dk 1 ). В статье [6] показано, что оно утраивает триангуляционное число. Из условия Т2 = 3Т 1 находим, что для любого (hb kx) оно разрешимо лишь в случае: (h 1 , k 1 ) ^ (h1+2k1, h1-k1). Подстановкой легко убедиться в переходах (f, 0) ^ (f, f) ^ (3f, 0) ^ (3f, 3f). Тем самым две серии (f, 0) и (f, f) объединяются в класс — 5 - 3m (рис. 3, соединения 16-48- ..., 25-75... и др. подразумеваются). Аналогично объединяются в классы 532 пары (гомологические по отношению друг к другу) серий, образованных skew -капсидами (h, k) и (h + 2k, h - k). Читатель легко сделает это самостоятельно с помощью табл. 1. При этом капсиды (h, k), не сводимые к более простым, названы нами генераторами. Их критерий: h и k взаимно просты, h - k не делится на 3 (табл. 2). Заметим, что изомеры существуют и среди генераторов.
Общее преобразование капсида
Нельзя ли найти преобразование капсида в любой другой: (hb kx) ^ (h2, k2) — как это сделано для фуллеренов, моделирующих микроэволюции скелетов радиолярий [7]? По сути, задача состоит в повороте вектора (h 1 , k 1 ), образующего угол а 1 с осью h, до вектора (h2, k2), образующего угол а2 с осью h, и дополнительном растяжении/сжатии (рис. 4). В ортогональной декартовой системе координат преобразование имеет вид:
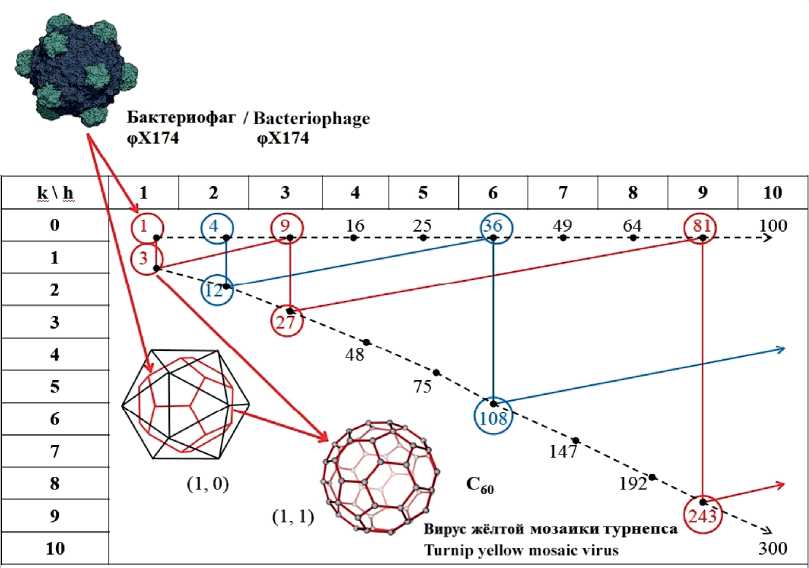
Рис. 3. Связь двух гомологических серий - 5 - 3m
Fig. 3. The relationship of two homologous series - 5 - 3m
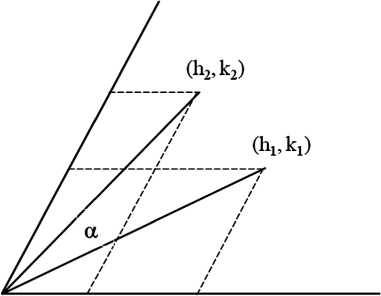
Рис. 4. К описанию общих преобразований икосаэдриче-ского капсида
Fig. 4. To the description of general transformations of an icosahedral capsid
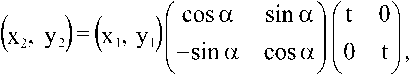
Таблица 2. Триангуляционные числа T для капсидов-генераторов

где a = a2 — a i , коэффициент растяжения t = (T2/T 1 )1/2.
Формулы перехода к косоугольной (60°) системе координат: X i = T 1 1/2 cosa 1 , У 1 = T 1 1/2 sinab x2 = T21/2 cosa2, y2 = T21/2 sina2. Выражаем тригонометрические функции через стороны треугольников (рис. 4) и после преобразований получаем:
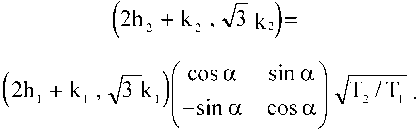
Задав (h 1 , k 1 ) и (h2, k2), находим коэффициент растяжения (T2/T 1 )1/2 и угол 0 < a < 60°.
Рассмотренные выше частные случаи следуют из полученного уравнения. Для преобразований подобия (h 1 , k 1 ) ^ (h2, k2) = (f h 1 , f k 1 ) подстановкой получим, что матрица поворота единичная: cos a = 1, sin a = 0, то есть a = 0. Действительно, указанные серии объединяют капсиды с гранями, одинаково ориентированными в системе координат (рис. 2). Для переходов между гомологичными сериями (h 1 , k 1 ) ^ (h2, k2) = (h 1 +2kb h1—k1) находим общее решение: a = arc sin [^ — 3k 1 (h1+k1) / 2T 1 ]. При k 1 = 0, a = 30°, при k 1 = h 1 , a = -30°; в обоих случаях коэффициент растяжения равен A 3.
Действительно, в дуальных преобразованиях последовательные переходы отличаются лишь знаком поворота.
Выводы
Икосаэдрические вирусы — весьма содержательный объект кристаллографического, комбинаторно-геометрического и алгебраического исследования. Сегодня неясно, какие из вирусов, теоретически упорядоченных в табл. 1 и 2, реально действуют в природе. Их многообразие должно ограничиваться размерным фактором. Но уже очевидна важная роль капсидов (1, 0), (2, 1) и (3, 1), стоящих в начале таблицы, не сводимых к более простым, а наоборот, продуцирующих (в морфологическом смысле) двумя указанными способами (преобразованием подобия и дуальным переходом к гомологической серии) подавляющую часть известного многообразия. В поисках способов разрушения капсидов следует обратить на них особое внимание.
Описывает ли найденное выше общее преобразование капсидов реальный процесс? Нам это неизвестно. Но можно задать вирусологам связанный вопрос: сопровождаются ли мутации коронавирусов изменением длины свернутой в капсиде ДНК/РНК? Если да, то из соображений целесообразности, с очевидностью, воплощенной природой в структуре капсида, следует ожидать, что вирус подгоняет его под необходимый объем содержимого. Возможно, здесь кроется лазейка для разрушающего физического/химического воздействия на механизм построения капсида мутирующего вируса. Из выполненного исследования можно сделать выводы.
-
• Номенклатуру икосаэдрического капсида следует строить на символе (h, k) его икосаэдрической грани, однозначно фиксирующем её ориентацию относительно плотнейшей упаковки белковых глобул.
-
• Классификация капсидов по т. г. с. — 5 - 3т (с плоскостями) и 532 (без них) логична. Но класс —5—3т состоит из двух гомологических серий (f, 0) и (f, f), где f = 1, 2, 3..., связанных дуальным переходом (h 1 , k 1 ) ^ (h 1 + 2k 1 , h 1 — k 1 ).
-
• Класс 532 тоже состоит из серий (f h, f k), порождаемых генераторами (h, k), узнаваемыми по критерию: h и k взаимно просты, h — k не делится на 3. У каждой серии в классе есть гомологичная, связанная дуальным переходом (h 1 , k 1 ) ^ (h 1 + 2k 1 , h 1 — k 1 ).
-
• Предположительно, коронавирус SARS-CoV-2 имеет капсид (2, 0), т. г. с. —5—3т , Т = 4, аналогичный 43

вирусу полиомы, второй в серии гомологов Т = f2, порождаемой генератором (1, 0) — бактериофагом фХ174. Его эффективность определена не специфической морфологией, а функциональными особенностями.
-
• Получено общее преобразование, описывающее переходы (h1, k1) → (h2, k2). Икосаэдрические капсиды — редкий случай биологической организации, когда удалось получить столь общий результат, даже если он имеет лишь теоретическое значение.
-
• Представляет интерес следующая задача: частота встречаемости икосаэдрических вирусов классов — 5 - 3m и 532 , в частности различных видов (h, k), ещё детальнее — вирусов-изомеров.
Список литературы Из опыта преподавания. III. Кристаллография икосаэдрических вирусов
- Лурия С., Дорнелл Дж., Балтимор Д., Кэмпбелл Э. Общая вирусология. М.: Мир, 1981. 680 с.
- Рис Э., Стернберг М. Введение в молекулярную биологию: от клеток к атомам. М.: Мир, 2002. 142 с.
- Caspar D. L. D., Klug A. Physical principles in the construction of regular viruses // Cold Spring Harbor Symp. Quant. Biol. 1962. V. 27. P. 1-24.
- Schmalz T. G., Seitz W. A., Klein D. J., Hite G. E. Elemental carbon cages // J. Am. Chem. Soc. 1988. V. 110. N 4. P. 1113-1127.
- Voytekhovsky Yu. L. Biomineral analogues in ontogeny and phylogeny // Paleont. J. 2015. V. 49. N 14. P. 1691-1697.
- Voytekhovsky Yu. L. Homological series of icosahedral viruses and fullerenes // Paleont. J. 2016. V. 50. N 13. P. 1505-1509.
- Voytekhovsky Yu. L., Stepenshchikov D. G. Fullerene transformations as analogues of radiolarian skeleton microevolution // Paleont. J. 2016. V. 50. N 13. P. 1544-1548.
