Изменчивость биохимических показателей и адаптация краснопыльниковой и жёлтопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения
 в условиях избыточного увлажнения")
Автор: Аганина Юлия Евгеньевна, Тарханов Сергей Николаевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-1 т.18, 2016 года.
Бесплатный доступ
Изучена динамика содержания фотосинтетических пигментов (хлорофиллов и каротиноидов), свободного пролина, стрессовых белков, аскорбиновой кислоты и активности пероксидаз в хвое форм сосны ( Pinus sylvestris ), различающихся цветом микростробилов, в условиях постоянного избыточного увлажнения почв в низовье р. Северная Двина. Определены уровни индивидуальной изменчивости данных показателей в разные календарные периоды. Выявлены существенные различия в сезонной динамике биохимических показателей, свидетельствующие о большей чувствительности и нуждаемости в развитии защитных функций краснопыльниковой сосны по сравнению с жёлтопыльниковой.
Сосна, изменчивость, адаптация, краснопыльниковая и жёлтопыльниковая форма, фото-синтетические пигменты, пероксидазная активность, свободный пролин, стрессовые белки, аскорбиновая кислота, увлажнение, почва
Короткий адрес: https://sciup.org/148204411
IDR: 148204411 | УДК: 631.524.6:582.475.4
Variability of biochemical indicators and adaptation of F. (var.) erythranthera sanio and F. (var.) sulfuranthera kozubow forms of the pine (Pinus sylvestris L.) in the conditions of excess humidification
Dynamics of maintenance the photosynthetic pigments (chlorophyll and carotinoids), free proline, stressful proteins, ascorbic acid and activity of peroxidases in needles of the pine forms ( Pinus sylvestris ) differing with color of microstrobil in the conditions of continuous excess humidification of soils in the lower reach of Northern Dvina river is studied. Levels of individual variability of these indicators during the different calendar periods are determined. The essential distinctions in seasonal dynamics of biochemical indicators testifying to bigger sensitivity and needs in development the protective functions of f. (var.) erythranthera Sanio in comparison with f. (var.) sulfuranthera Kozubow are revealed.
Текст научной статьи Изменчивость биохимических показателей и адаптация краснопыльниковой и жёлтопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения
Высокий уровень генетического полиморфизма лесообразующих хвойных видов, проявляющийся в их формовом разнообразии, связывается с действием разнообразных механизмов адаптации и условий среды. Адаптивные возможности растений выявляются по их ответным реакциям на внешние воздействия. По мнению Н.Е Судачковой [12], можно допустить, что древесные растения, испытывающие стресс в экстремальных условиях местообитания в течение всей жизни, как например, болотные, притундровые леса, постоянно находятся в стадии стабилизации, т.е. не на том уровне метаболизма, который определён дереву его генетическими возможностями, а на том, который позволяет компенсировать затраты на стрессовый метаболизм. Постоянное избыточное увлажнение почв приводит к корневой гипоксии, вызывая хронический стресс у хвойных деревьев. В этих условиях дефицит кислорода отражается на интенсивности физиолого-биохимических процессов, как корневой системы, так и наземных органов.
Наибольшее число сведений, касающихся стрессового метаболизма у древесных растений, относится к исследованиям по обнаружению низкомолекулярных метаболитов с защитной функцией. Всего, по разным оценкам, насчитывается порядка 15-20 различных признаков неспецифической составляющей стресса у растений [16, 6, 10]. Для гипоксического стресса (не у всех видов и тканей древесных растений) характерно накопление крахмала, аминокислот и белков, органических кислот, анаэробной части цикла Кребса [17].
Цель работы : изучение сезонной динамики биохимических показателей хвои в связи с адаптацией разных форм сосны ( Pinus slvestris ), различающихся цветом пыльников, в стрессовых условиях.
Методика исследования. Объектами исследований являются северотаёжные сосняки кустарничковосфагновые на верховых торфяных почвах в низовье Северной Двины. В сосняках кустарничково-сфагновых
торф – сфагновый или пушице-сфагновый, низкой степени разложения. Торф имеет сильнокислую реакцию среды (рН солевой суспензии 2,6-3,2), высокую обменную и гидролитическую кислотность, очень низкую степень насыщенности основаниями (11-14%). Содержание зольных элементов в верхнем торфе составляет 2-4%. Торф верхних горизонтов характеризуется низкой объёмной массой, высокой полевой влажностью, близкой к полной влагоёмкости (90-94%).
Для определения биохимических признаков у форм сосны, выделенных по цвету пыльников (жёлтопыльниковая и краснопыльниковая), в 2015 г. отбирали образцы хвои на 10 деревьях каждой формы. Учитывая сезонную изменчивость этих признаков, отбор образцов производили на одних и тех же деревьях в начале роста побегов (конец мая), в конце завершения их линейного прироста (середина июля), в конце вегетационного периода (конец сентября) и перед перезимовкой (начало ноября). В лабораторных условиях спектрофотометрическим методом (на спектрофотометре NanoDrop 2000C) у разных форм сосны в хвое, сформировавшейся в 2014 году, определяли содержание фотосинтетических пигментов [20, 9], пероксидазную активность [2], содержание свободного пролина [22], стрессовых белков, аскорбиновой кислоты [1, 8, 19]. Долю хлорофиллов в светособирающем комплексе (ССК) расчитывали по формуле [23, 24]. Уровни изменчивости признаков оценивали по эмпирической шкале С.А Мамаева [7]. При сравнении одноимённых признаков и свойств использовали критерий F -Фишера и t -Стъюдента [11]. Дисперсионный анализ проводили с использованием пакета лицензионных программ Excel.
Экспериментальная часть, анализ, обобщение и разъяснение полученных данных. Содержание и состав фотосинтетических пигментов являются показателями основного метаболизма растений. Процессы ассимиляции в первую очередь связаны с фотосинтезом, а значит, и с пигментным аппаратом. Деревья разных форм могут сильно различаться по содержанию фотосинтетических пигментов из-за различного диапазона нормы их реакции на воздействие условий среды. Менее вариабельным является содержание в хвое хлорофилла а и каротиноидов (C.V.<20%). Более изменчивым является содержание хлорофилла b. Коэффициенты вариации этого показателя в определённые периоды могут достигать очень высокого уровня (C.V.>40%). В разные календарные периоды и годы индивидуальная вариабельность содержания хлорофиллов и каротиноидов может сильно различаться. Можно отметить повышение индивидуальной изменчивости содержания хлорофилла b осенью, особенно у краснопыльниковой формы (C.V. - до 54%), что, по-видимому, связано с более выраженными индивидуальными различиями в ответных реакциях деревьев этой формы на действие стрессовых факторов в связи с сезонными изменениями климатических и почвенногидрологических условий.
Уровни индивидуальной изменчивости относительных величин содержания пигментов: хлорофилла а и b (Х а /Х b ), хлорофиллов и каротиноидов (Х а + b /К) также выше осенью, особенно у краснопыльниковой формы. Причём относительный показатель Х а /Х b варьирует в большей степени ( C.V. =19-41% - у краснопыльниковой формы и C.V. =10-47% - у жёлтопыльниковой) по сравнению с показателем Х а + b /К ( C.V. =8-26% и C.V. =6-20%, соответственно). Это свидетельствует о больших различиях деревьев краснопыльниковой формы между собой в соотношении фотосинтетических пигментов.
То же самое было отмечено ранее при изучении индивидуальной изменчивости этих показателей в целом для этой популяции сосны в низовье Северной Двины. Изменение величины Ха+b/К рассматривается как приспособительная реакция свето-собирающего комплекса (ССК) фотосинтетических пигментов деревьев на условия внешней среды. Доля хлорофиллов, локализованных в ССК, у деревьев разных форм варьирует в пределах 20-70% от общего фонда зелёных пигментов. Причём величина коэффициента вариации показателя ССК также больше осенью (C.V.=22-32%) по сравнению с весенне-летним периодом (C.V.=13-17% - у краснопыльниковой формы и C.V.=7-8% - у жёлтопыльниковой). Соответственно, индивидуальная изменчивость этого показателя в весенне-летний период у краснопыльниковой формы характеризуется средним, а у жёлтопыльниковой формы - низким уровнем. Эти амплитуды связаны с адаптацией фотосинтетического аппарата сосны в сосняках кустарничково-сфагновых к световым условиям в высоких широтах. Осенью индивидуальные различия между деревьями по величине Ха/Хb становятся существенными, особенно у жёлтопыльниковой формы (F=14,5-79,5; F0,05=3,2; Fo,oi=5,5), что связано с широтой нормы реакции (реакцией генотипов на условия среды). Это же можно отметить для показателя ССК у жёлтопыльниковой формы, хотя и в меньшей степени (F=4,8-5,1; Fo,05=3,2; F0,01=5,5). Установлены достоверные различия дисперсий выборок деревьев краснопыльниковой и жёлтопыльниковой сосны по величине Ха/ХЬ в весеннелетний период (период линейного роста побегов) (F=4,1-5,1; F0,05=3,2; F0,01=5,5), ССК - в начале роста (F=4,0; F0,05=3,2). Однофакторный дисперсионный анализ подтверждает влияние фактора «форма» на изменчивость параметров фотосинтетического аппарата в конце мая, перед началом линейного роста побегов (F=7,10-20,45; F0,05=4,41).Двухфакторный дисперсионный анализ показывает совместное влияние факторов «форма» и «сезон» на количественное содержание фотосинтетических пигментов, а также величины Хa+b/К и ССК сосны (F=2,94-10,72; F0,05=2,73). Уровень индивидуальной изменчивости пероксидазной активности хвои у краснопыльниковой формы больше (очень высокий, C.V. >40%) по сравнению с жёлтопыльниковой (высокий, C.V.<40%) во все календарные периоды. Причём, осенью индивидуальная вариабельность активности пероксидаз повышается по сравнению с весенне-летним периодом у обеих форм (F=3,3-5,9; F0,05=3,2; F0,01=5,5). Это свидетельствует о более широкой норме реакции деревьев краснопыльниковой формы на влияние условий среды, по сравнению с жёлтопыльниковой, особенно в осенний период. Уровень индивидуальной изменчивости содержания свободного пролина средний (C.V.<20%) и повышенный (C.V.>20%) у обеих форм. Причём, осенью у краснопыльниковой сосны он снижается (C.V.=13-19%) по сравнению с весенне-летним периодом (C.V.=24-29%). Различия дисперсий этого показателя в выборках деревьев в ноябре и других календарных периодах достоверны на принятых условиях значимости у обеих форм (F=3,5-19,8; F0,05 = 3,2; F0,01 = 5,5). Более существенные различия наблюдаются у краснопыльниковой сосны (F=7,4-19,8), что также свидетельствует о более широкой норме реакции этой формы по данному показателю в ноябре, перед перезимовкой. Уровень индивидуальной изменчивости содержания в хвое стрессовых белков очень высокий у обеих форм (C.V. =40-107%), причём в выборках деревьев жёлтопыльниковой сосны различия дисперсий этого показателя между разными календарными периодами существенны (F=3,2-29,2; F0,05=3,2; F0,01=5,5). Уровень индивидуальной вариабельности содержания аскорбиновой кислоты в хвое у обеих форм больше в весеннелетний период (C.V. =31-59%) по сравнению с осенью (C.V.=10-16%). Это свидетельствует о более выраженных индивидуальных различиях реакций деревьев по этому показателю на действие внешних факторов в период активной вегетации.
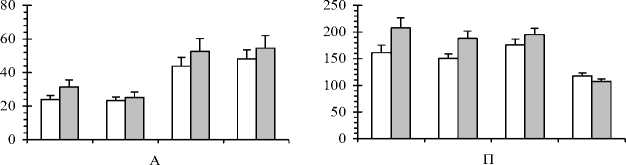
Ритмичность физиолого-биохимических процессов влияет на устойчивость сосны к воздействию внешних факторов и может иметь решающее значение при адаптации. Количественное содержание и качественный состав фотосинтетических пигментов, изменение их соотношения в листьях - важные и чувствительные показатели физиологического состояния растений, направленности адаптивных реакций при воздействии стрессовых факторов [3]. В период вегетации содержание фотосинтетических пигментов зависит от фенофаз развития деревьев. Характер сезонной изменчивости показателей фотосинтетического аппарата у форм с разным цветом микростробилов довольно сходный (рис. 1). Перед перезимовкой наблюдается сущест-венное снижение содержания хлорофиллов a и b у обеих форм по сравнению с весенне-летним периодом ( t =2,54-3,17; t 0,05 =2,26-3,25). Осенью происходит существенное повышение содержания каротиноидов у обеих форм t =12,26-14,03; 1 0,01 = 4,78. Однако наблюдаются и различия в сезонной динамике пигментов у этих форм. После завершения линейного роста (середина июля) у желтопыльниковой формы существенно больше концентрация хлорофилла b по сравнению с его содержанием в начале роста побегов (конец мая) и сентябре ( t =2,32-3,58; t 0,05 =2,26; t 0,01 =3,25).
Таким образом, у жёлтопыльниковой формы чётко выражен летний максимум этого показателя, в отличие от краснопыльниковой формы, у которой июльский максимум в период вегетации 2015 г. проявляется несущественно ( t < t 0,05 ).
Изменения соотношений пигментов в листьях -важные и чувствительные показатели физиологического состояния, направленности адаптивных реакций растений при воздействии стрессовых условий [3]. Известно [5], что водный стресс (в том числе и в случае насыщения почвы водой вокруг корней растений) наиболее сильно действует на свето-собирающий хлорофилл а / b - белковый комплекс. Осенью в листьях в условиях водного стресса хлорофилл а разрушается быстрее хлорофилла b . Действие плохой аэрации почвы является до некоторой степени косвенным: синтез хлорофилла задерживается вследствие общего нарушения метаболизма.
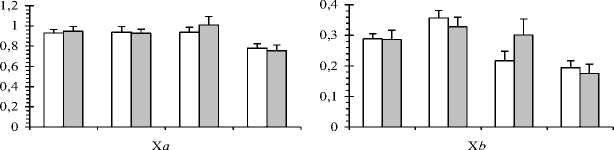
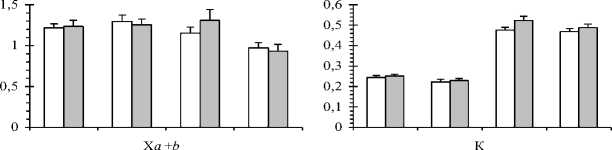
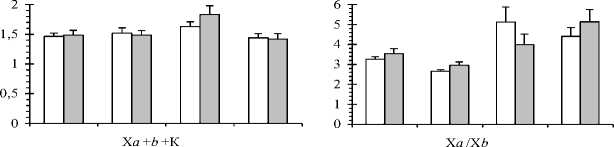
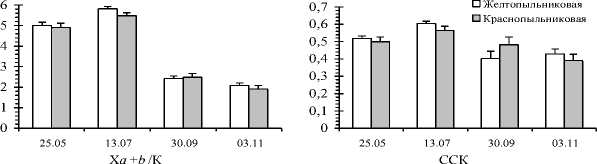
Рис. 1. Сезонная динамика (среднее значение с ошибкой) показателей фотосинтетического аппарата в 2015 г. у форм сосны с разным цветом микростробилов
Ха, ХЬ - содержание хлорофиллов а и b соответственно, мг/г воздушно сухой массы хвои; Ха+b - суммарное содержание хлорофиллов а и b соответственно, мг/г воздушно сухой массы хвои; К - содержание каротиноидов, мг/г воздушно сухой массы хвои; Ха+b+K - общая сумма пигментов, мг/г воздушно сухой массы хвои; Ха/Хb - отношение хлорофилла а к хлорофиллу b; Ха+b/K - отношение суммы хлорофиллов а и b к каротиноидам; ССК - доля хлорофиллов светособирающего комплекса
Установлено [15], что в сосняках сфагновой группы адаптация деревьев сосны к воздействию внешних факторов через увеличение емкости ССК у разных форм может проявляться по-разному. Показано [13], что увеличение величины соотношения хлорофиллов а и b свидетельствует о негативном влиянии внешней среды на ССК хлоропластов. По нашим данным, доля хлорофиллов, локализованных в ССК в разные сезонные периоды (конец мая - начало ноября 2015 г.) составляет в среднем 39-57% от общего фонда пигментов хвои в выборках деревьев с красной окраской микростробилов и 46-60% - с желтой окраской пыльников (рис. 1). Эта амплитуда связана с адаптацией фотосинтетического аппарата сосны в сосняках кустарничково-сфагновых к световым условиям в высоких широтах [14]. Различия этих форм по доле хлорофиллов, локализованных в ССК общего фонда пигментов, в другие периоды вегетации (конец мая - сентябрь) не достоверны по t -критерию на 5%-ном уровне значимости.
Каротиноиды играют важную роль в фотосинтезе, выполняя функции светосборников и фитопротекторов [21]. По величине отношения суммы хлорофиллов к каротиноидам, величине отношения содержания хлорофилла а к хлорофиллу b достоверные различия при критических значениях t-критерии между этими формами также отсутствует. Соответственно можно предполагать, что обе формы испытывают сходное негативное воздействие внешней среды. Видимо, в хвое, подверженной действию водного и гипоксического стресса, сосна с красной и желтой окраской микростробилов не различается по интенсивности разрушения хлорофиллов.
Нами установлены достоверные сезонные различия по величине отношения хлорофилла а к хлорофиллу b у желтопыльниковой формы (t=2,42-4,23; to,05=2,26; to,oi=3,25). Существенно меньше этот показатель в середине лета, а значительно больше - в конце мая (начало роста) и особенно, осенью (конец вегетации). У краснопыльниковой сосны этот показатель существенно увеличивается в ноябре, после завершения периода вегетации (t=2,37-3,36; t0,05=2,26; to,oi=3,25). Очевидно, отрицательное воздействие внешних факторов на деревья этой формы сильнее проявляется осенью. Из-за увеличения содержания каротиноидов осенью у обеих форм показатель Хa+b/К меньше по сравнению с летним (t=2,28-4,17) и весенним периодами (t=8,76-22,22; Zo,o5=2,26; 6o,oi=3,25; t0,001=4,78). Эти пигменты защищают хлорофилл от фотоокисления и являются более устойчивыми к действию неблагоприятных факторов в осенне-зимний период. Увеличение содержания каротиноидов рассматривается как адаптивная реакция, направленная на повышение устойчивости фотосинтетического аппарата, предотвращение фотодинамической деструкции в суровых северных условиях [3]. В весенний период (да начала линейного роста побегов) величина отношения хлорофиллов к каротиноидам значительно больше, чем осенью. Причем эти различия более выражены у желтопыльниковой формы (t=4,17; 6o,oi=3,25), чем у краснопыльниковой (t= 2,28; t0,05=2,26). Это можно рассматривать как при- способительную реакцию ССК фотосинтетических пигментов этих форм на изменение условий внешней среды, причем у желтопыльниковой сосны она имеет более выраженный характер.
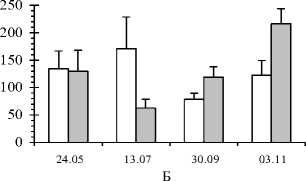
Увеличение активности пероксидаз можно рассматривать как защитную реакцию растений на воздействие стрессовых факторов, препятствующую развитию неконтролируемых окислительных процессов. По нашим данным, пероксидазная активность у обеих форм сосны почти в 2 раза выше осенью, по сравнению с весенне-летним периодом (рис. 2). В начале и при завершении линейного роста побегов и хвои этот показатель значительно меньше (t=2,45-4,32; t0,05=2,26; to,oi=3,25). Следовательно, осенью деревьям присущ более напряженный обмен веществ и для обеспечения нормального прохождения окислительных процессов пероксидазная активность хвои повышается.
Аккумуляция пролина считается типичным ответом растения на воздействие различных отрицательных факторов [4]. Наблюдалось достоверное снижение содержания свободного пролина у обеих форм сосны при критических значениях t -критерия в ноябре, после завершения вегетации. Причем, более значительно снижалось содержание пролина у краснопыльниковой сосны ( t =5,11-7,03; t 0,05 =2,26; t o,oi =3,25; t o,ooi =4,78) по сравнению с желтопыльниковой ( t =2,87-4,90; t o,o5 =2,26; t o,oi =3,25; t o,ooi =4,78). Можно предполагать, что его сезонная динамика связана с ритмикой ростовых процессов и оттоком энергетических и пластических веществ, в т.ч и свободных аминокислот, во вновь образующиеся побеги.
□ Желтопыльниковая
Краснопыльниковая

24.05 13.07 30.09 03.11
АК
Рис. 2. Сезонная динамика биохимических показателей (среднее значение с ошибкой) хвои в 2015 г. у форм сосны с разным цветом микростробилов
А - активность пероксидаз, усл. ед.; П - содержание пролина, мкг/г воздушно сухой массы хвои; Б - содержание стрессовых белков, мкг/г воздушно-сухой массы хвои; АК - содержание аскорбиновой кислоты, мкг/г воздушно-сухой массы хвои
В процессе адаптации к стрессовым условиям происходит формирование новых изоэнзимов или стрессовых белков. Согласно нашим данным, содержание стрессовых белков у краснопыльниковой сосны больше весной и повышается осенью, особенно в ноябре. Очевидно, эта форма более чувствительная к воздействию стрессовых факторов в отношении этого показателя весной и осенью по сравнению с желтопыльниковой. В начале ноября наблюдается существенное повышение содержания стрессовых белков у краснопыльниковой формы по сравнению с жёлтопыльниковой сосной ( t =2,46; t o,o5 =2,26). Влияние фактора «форма» на изменчивость этого показателя перед перезимовкой подтверждается однофакторным дисперсионным анализом ( F =6,06; F o,o5 =4,41). В середине июля, в период завершения линейного роста побегов хвоя деревьев краснопыльниковой формы накапливает больше свободного пролина по сравнению с жёлтопыльниковой ( t =2,26; t o,o5 =2,26) (рис. 2). Это свидетельствует в пользу того, что краснопыльниковая сосна более подвержена воздействию стрессовых факторов и в большей степени нуждается в развитии защитных функций. Дисперсионный анализ подтверждает влияние фактора «форма» на изменчивость содержания пролина в июле, после завершения линейного роста ( F =5,13-7,45; F 3,05 =3,97-4,41).
Аскорбиновая кислота (витамин С) наряду с другими соединениями участвует в регуляции окислительновосстановительного потенциала, с которым связана активность многих ферментов и физиолого-биохимических ре- акций, в т.ч. таких жизненно необходимых как фотосинтез и дыхание. Её содержание тесно связано с условиями произрастания и физиологическим состоянием растений [18]. Содержание аскорбиновой кислоты в хвое к осени у обеих форм резко увеличивается (t=8,75-20,50; to,oo1=4,78). Это может быть связано с повышением роли аскорбиновой кислоты как антиоксиданта растительных клеток при воздействии экстремальных факторов среды.
Выводы: в условиях постоянного избыточного увлажнения почвы выявлены существенные различия в сезонной динамике биохимических показателей у форм сосны, свидетельствующие о большей чувствительности и нуждаемости в развитии защитных функций краснопыльниковой сосны по сравнению с жёлтопыльниковой. Наблюдается снижение концентрации хлорофиллов перед перезимовкой и повышение содержания каротиноидов осенью, вследствие общего замедления метаболизма сосны. Установлено увеличение пероксидазной активности осенью (почти в 2 раза) у обеих форм и концентрации стрессовых белков перед перезимовкой у краснопыльниковой сосны, в связи с более экстремальными условиями. Наблюдается существенное снижение содержания свободного пролина после окончания периода вегетации, особенно у краснопыльниковой формы ( t > t o,oo1 ). Осенью происходит значительное увеличение содержания аскорбиновой кислоты у обеих форм сосны.
Исследования выполнялись в рамках государственного задания Федерального исследовательского центра комплексного изучения Арктики РАН (проект № 0410-2014-0025).
Список литературы Изменчивость биохимических показателей и адаптация краснопыльниковой и жёлтопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения
- Воскресенская, О.Л. Большой практикум по биоэкологии. Ч. 1: учеб. пособие//О.Л. Воскресенская, Е.А. Алябышева, М.Г. Половникова. -Йошкар-Ола.: Мар. гос. ун-т. 2006. 107 с.
- Бояркин, А.Н. Быстрый метод определения активности пероксидазы//Биохимия. 1951. Вып.1, № 4. С. 352-357.
- Головко, Т.К. Пигментный комплекс растений природной флоры Европейского Северо-Востока // Т.К. Головко, И.В. Далькэ, О.В. Дымова, И.Г. и др. // Известия Коми научного центра УрО РАН. 2010. Т. 1. С. 39-46.
- Климов, С.В. Адаптация растений к стрессам через изменение донорно-акцепторных отношений на разных уровнях структурной организации//Успехи современной биологии. 2008. Т. 128, № 3. С. 281-299.
- Крамер, П.Д. Физиология древесных растений: Пер с анг.//П.Д. Крамер, Т.Т. Козловский. -М.: Лесн. Пром-ть, 1983. 464 с.
- Лукаткин, А.С. Холодовое повреждение теплолюбивых растений и окислительный стресс. -Саранск: Изд-во Мордов. ун-та, 2002. 208 с.
- Мамаев, С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). -М.: Наука, 1972. 284 с.
- Методы биохимического исследования растений. -М.: Агропромиздат. 1987. 430 с.
- Третьяков, Н.Н. Практикум по физиологии растений//Н.Н. Третьяков, Т.В. Карнаухов, Л.А. Паничкин и др. -М.: Агропромиздат, 1990. 271 c.
- Пятыгин, С.С. Стресс у растений: физиологический подход//Журнал общей биологии. 2008. Т. 69, №. 4. С. 294-298.
- Свалов, В.В. Вариационная статистика. -М.: Лесная промышленность, 1977. 178 с.
- Судачкова, Н.Е. Состояние и перспективы изучения влияния стрессов на древесные растения//Лесоведение. 1998. № 2. С. 3-9.
- Тарханов, С.Н. Лесные экосистемы Северной Двины в условиях атмосферного загрязнения: диагностика состояния//С.Н. Тарханов, Н.А. Прожерина, В.Н. Коновалов. -Екатеринбург: УрО РАН, 2004. 333 с.
- Тарханов, С.Н. Морфоструктура и изменчивость биохимических признаков популяции сосны (Pinus sylvestris L.) в стрессовых условиях устья Северной Двины/С.Н. Тарханов, С.Ю. Бирюков//Сиб. экол. журн. 2014. №2. С. 319-327.
- Тарханов, С.Н. Формы внутрипопуляционной изменчивости хвойных в условиях атмосферного загрязнения (на примере Северо-Двинского бассейна). -Екатеринбург: Изд-во УрО РАН, 2010. 230 с.
- Тарчевский, И.А. Метаболизм растений при стрессе. -Казань: Фэн. 2001. 448 с.
- Чиркова, Т.В. Метаболизм этанола и лактата в тканях древесных растений, различающихся по устойчивости к недостатку кислорода//Физиология растений. 1975. Т. 22. Вып. 5. С. 952-958.
- Чупахина, Г.Н. Адаптация растений к нефтяному стрессу/Г.Н. Чупахина, П.В. Масленников//Экология. 2004. № 5. С. 330-335.
- Чупахина, Г.Н. Количественное определение аскорбиновой кислоты колориметрическим методом: специальный практикум по биохимии и физиологии растений. -Томск: Изд-во Томского ун-та, 1974. С. 27-31.
- Шлык, А.А. Определение хлорофиллов и каротиноидов в экстрактах зелёных листьев/Биологические методы в физиологии растений. -М., 1971. С. 154-170.
- Шуляковская, Т.А. Динамика содержания основных метаболитов в сеянцах сосны обыкновенной/Т.А. Шуляковская, Л.А. Чиненова, С.М. Шредерс, Г.К. Канючкова//Лесоведение. 2005. № 2. С. 53-61.
- Bates, L.S. Rapid determination of free proline for water-stress studies/L.S. Bates, R.P. Waldren, J.D. Teare//Plant and Soil. 1973. Vol. 39. № 1. P. 205-206.
- Lichenthaler, H.K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes//Methods in Enzymology, 1987. Vol. 148. P. 350-382.
- Lichenthaler, H.K. Vegetation stress: an introduction to the stress concept in plant//J. Plant Physiol. 1996. Vol. 148. P. 4-14.
