Изменчивость основных систематических признаков гибридной формы елей Picea abies и P. obovata (Pinaceae)
")
Автор: Казанцева М.Н., Попов П.П., Арефьев С.П.
Журнал: Вестник Нижневартовского государственного университета @vestnik-nvsu
Рубрика: Экология растений
Статья в выпуске: 4 (72), 2025 года.
Бесплатный доступ
Работа посвящена изучению индивидуальной и географической изменчивости основных систематических признаков гибридной (промежуточной) формы елей европейской (Picea abies (L.) Karst.) и сибирской (P. obovata Ledeb.) – длины шишек и формы семенных чешуй. Ее актуальность определяется тем¸ что эти признаки у гибридных популяций ели изучены недостаточно и главным образом визуально-описательным путем. Основная методическая особенность данного исследования заключаются в использовании метрических и приемов обработки математико-статистических большого исходного материала. Использован материал с 24 ключевых участков в районе распространения гибридной формы ели на территории России. Выборка шишек с каждого участка включала не менее 100 штук (по одной с дерева); общее число деревьев составило 4581 экземпляр. Анализ полученного материала показал, что, в популяциях гибридной формы елей P. abies и P. obovata сложились промежуточные параметры основных систематических признаков. Средняя длина шишек гибридной формы ели составляет 76(63–91) мм. Коэффициенты сужения (coefficient of narrowing – Cn) и вытянутости (coefficient of projection – Cp) верхней части семенных чешуй равны соответственно 45(36–54) и 52(47–71)%, их разность (Cn–Cp) находится в пределах от -16 до +23%. Индивидуальная (внутрипопуляционная) изменчивость этих признаков значительно выше географической. В межпопуляционной изменчивости показателей формы семенных чешуй и длины шишек имеется корреляция среднего уровня. Результаты исследования позволяют обосновать морфологогеографическую дифференциацию промежуточной формы ели на три группы в пределах общего района ее распространения, а также могут быть использованы для выделения более мелких таксонов гибридной формы елей P. abies и P. obovata. Учитывая наличие связи рассматриваемых параметров с количеством и качеством семян в шишках, полученные результаты могут быть полезны для практического лесоводства при отборе наиболее перспективных лесосеменных участков.
Picea abies, P. obovata, гибридная форма ели, длина шишек, показатели формы семенных чешуй, изменчивость, корреляция
Короткий адрес: https://sciup.org/14134399
IDR: 14134399 | УДК: 574.3:575.21 | DOI: 10.36906/2311-4444/25-4/02
Variability of the Main Systematic Features of the Hybrid Shape of Firs Picea abies and P. obovata (Pinaceae)
The article is devoted to the study of individual and geographical variability of the main systematic traits of the hybrid (intermediate) form of European spruce (Picea abies (L.) Karst.) and Siberian (P. obovata Ledeb.) - the length of the cones and the shape of the seed scales. The relevance of the study is determined by the fact that these traits in hybrid populations of spruce have been insufficiently studied and mainly by visual-descriptive means. The main methods of this study are the use of metric and mathematicalstatistical techniques for processing large source material. The material was collected at 24 sites in the area of distribution of the hybrid form of spruce on the territory of Russia. The sample of cones from each plot included at least 100 pieces (one from a tree); the total number of trees was 4581 specimens. The analysis of the obtained material showed that intermediate parameters of the main systematic features were formed in the populations of the hybrid form of European and Siberian spruce. The average length of cones of a hybrid form of spruce is 76 (63–91) mm. The coefficients of narrowing (Cn) and projection (Cp) of the upper part of the seed scales are 45(36–54) and 52(47–71)%, respectively, their difference (Cn–Cp) is in the range from -16 to +23%. Individual (intrapopulation) variability of these traits is much higher than the geographical one. In the interpopulation variability of the parameters of the shape of the seed scales and the length of the cones, there is a correlation of the average level. The results of the study make it possible to substantiate the morphological and geographical differentiation of the intermediate form of spruce into three groups within the general area of its distribution, and can also be used to identify smaller taxa of the hybrid form of European and Siberian spruces. Given the relationship between the parameters under consideration and the quantity and quality of seeds in cones, the results obtained can be useful for practical forestry in the selection of the most promising forest-seed plots.
Текст научной статьи Изменчивость основных систематических признаков гибридной формы елей Picea abies и P. obovata (Pinaceae)
Казанцева М.Н., Попов П.П., Арефьев С.П. Изменчивость основных систематических признаков гибридной формы елей Picea abies и P. obovata (Pinaceae) // Вестник Нижневартовского государственного университета. 2025. № 4(72). С. 16-28.
Феномен интрогрессивной гибридизации видов древесных растений широко распространен и давно вызывает научный и практический интерес, определяемый их особой экологической и хозяйственной значимостью. В последние годы внимание к этому вопросу возросло в связи задачами сохранения популяционно-генетического разнообразия, в выявлении которого наряду с традиционными морфологическими методами большое значение приобрели методы молекулярно-генетические. Значительное внимание вызывает также оценка устойчивости разных популяционно-генетических форм деревьев к современным изменениям климата, особенно ощутимым в северных регионах, где основными эдификаторами лесной растительности являются хвойные виды деревьев. Современные исследования интрогрессивной гибридизации хвойных [34; 27; 28; 30; 41], в частности видов елей ( Picea ) [29; 32; 35; 36; 42; 33; 40], показали генетическую специфику популяционно-географических форм хвойных, определяемых интрогрессией, нередко имеющую сложную эволюционно-историческую природу. Показана также значительная роль интрогрессии в сохранении генетического разнообразия хвойных в меняющихся условиях, в частности, повышенная устойчивость «промежуточных» межвидовых форм к наблюдаемым ныне изменениям климата. Вместе с тем, молекулярно-генетические исследования елей на территории России пока не имеют сколь-нибудь масштабного характера, актуальна дифференциация гибридных форм по генетически обусловленным морфологическим признакам.
Популяции гибридной (промежуточной или переходной) формы елей европейской ( Picea abies (Linnaeus) H. Karsten, 1881) и сибирской ( Picea obovata Ledebour, 1833) располагаются, главным образом, в российской части ареала [5; 15; 21; 31]. Впервые эта ель была описана на территории Финляндии в 1863 г. по заметно отличающейся форме верхней

части семенных (шишечных) чешуй отдельных особей под названием fennica (Regel) и medioxima (Nylander) [13; 22]. В.Л. Комаров [14] охарактеризовал ее в ранге вида – ель финская ( Picea fennica Regel). В дальнейшем она так и называлась ( Picea fennica (Regel) Kom.) [4; 16; 21], хотя не все систематики были согласны с ее видовым статусом. Наиболее полно и достаточно конкретно условные границы распространения гибридной формы ели в пределах России указал Л.Ф. Правдин [39]. По его данным юго-западная граница ее проходит примерно от Санкт-Петербурга, западнее Москвы к южной границе ареала, северо-восточная – от восточного берега Чешской губы к Сыктывкару, Перми и далее к южной границе ареала. Район распространения гибридных популяций ели, выделенный с использованием метрических приемов изучения морфологии семенных чешуй, в основном тот же, но границы проходят несколько иначе [19]. Леса, образованные гибридной формой ели, распространены в центральной части Европейской России, они имеют большое хозяйственное и экологическое значение [1; 6; 24; 26]. Гибридные, формы-особи, характеризуются многими авторами давно (см. обзор Л.В Орловой, А.А. Егорова [16]), но параметры признаков, их изменчивость до сих пор мало изучены и в основном визуальноописательным путем в отдельных регионах. Целью настоящей работы является изучение индивидуальной и географической изменчивости линейных параметров шишек и семенных чешуй гибридной формы елей европейской и сибирской на территории России на основе математико-статистического анализа.
Материал и методы
Исследования проведены в районе распространения гибридной формы елей P. abies и P. obovata [19; 21]. Здесь были выделены 3 группы (фенотипы) популяций ели, географическое расположение которых определено по преобладанию особей промежуточных фенотипов, оцениваемых на основе биометрических параметров семенных чешуй [19]: I – фенотип P.emm. ( Picea europaea-medioxima-medioxima ), II – фенотип P.m. ( Picea medioxima ), III – фенотип P.mms. ( Picea medioxima-medioxima-sibirica (рис. 1).
При сборе и обработке материалов для данной статьи использована апробированная методика [12] с использованием элементов многомерного математического анализа [7].
В ходе работ 2017–2020 гг. материал собран на 56 участках ельников. На каждом участке формировали выборку по 100 и более шишек средней длины (по одной с дерева). Для настоящего анализа взято 24 ключевых участка, наиболее равномерно распределенных по району исследований, всего на них собран 4581 экземпляр шишек. У шишек определяли длину ( L c – Length of cones ), показатели формы семенных чешуй в виде коэффициентов сужения ( C n – coefficient of narrowing ) и вытянутости ( C p – coefficient of projection ) верхней части, их разность ( C n –C p ) [18], а также корреляционные зависимости между этими признаками с использованием коэффициента корреляции Пирсона. Показатели определяли для каждой особи, выборки, по всей совокупности выборок, а в ее пределах по фенотипам (группам) популяций промежуточной формы ели.

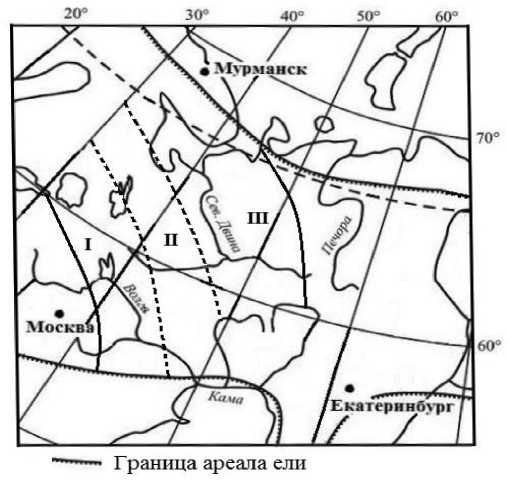
Рис. 1. Географическое распространение групп (фенотипов) популяций гибридной формы елей Picea abies и P. obovata в России. I – P emm ., II – P.m. , III – P.mms .
Результаты и обсуждение
В индивидуальной (внутрипопуляционной) изменчивости показателей L c , C n и C p наблюдается определенная закономерность (табл. 1). Диапазон показателя длины шишек составляет от 45–50 до 100–120 мм. Максимальное среднее для особи значение этого показателя превышает минимальное примерно в 2 раза, при коэффициенте вариации 13– 14%. Таким же уровнем изменчивости характеризуется показатель C p . Изменчивость показателя C n несколько выше, различие определяется в 2,4 (2,33–2,45) раза, вариация его составляет 17–19%.
Таблица 1
Индивидуальная изменчивость показателей длины шишек и формы семенных чешуй в гибридных популяциях елей Picea abies и P. obovata в ключевых пунктах на территории России
|
№ пункта |
n |
L c |
C n |
C p |
|||
|
X( min–max ) |
C v |
X( min–max ) |
C v |
X( min–max ) |
C v |
||
|
I фенотип популяций |
|||||||
|
1 |
208 |
85(55–120) |
14 |
38(22–57) |
19 |
60(42–83) |
15 |
|
2 |
120 |
80(55–110) |
14 |
36(33–50) |
19 |
59(47–75) |
11 |
|
3 |
164 |
81(50–105) |
13 |
40(23–59) |
19 |
57(40–85) |
14 |
|
4 |
175 |
88(55–115) |
16 |
39(26–60) |
18 |
57(40–85) |
14 |
|
5 |
140 |
80(60–120) |
14 |
39(24–63) |
22 |
61(45–84) |
14 |
|
6 |
150 |
91(53–122) |
15 |
39(23–57) |
18 |
57(40–73) |
11 |
|
7 |
210 |
79(55–110) |
14 |
39(20–60) |
21 |
57(39–78) |
14 |
|
8 |
200 |
78(50–110) |
15 |
40(24–62) |
20 |
56(37–81) |
14 |
|
II фенотип популяций |
|||||||
|
9 |
252 |
78(55–115) |
14 |
42(23–67) |
19 |
55(39–74) |
12 |
|
10 |
167 |
73(50–105) |
15 |
45(26–62) |
19 |
57(39–72) |
13 |
|
11 |
220 |
64(45–95) |
15 |
47(30–68) |
17 |
53(38–74) |
12 |
|
12 |
130 |
80(55–110) |
14 |
44(29–64) |
19 |
54(40–70) |
12 |

|
13 |
130 |
82(55–115) |
14 |
44(27–63) |
18 |
53(40–84) |
14 |
|
14 |
135 |
82(60–115) |
13 |
41(27–57) |
16 |
53(35–71) |
14 |
|
15 |
200 |
79(55–110) |
13 |
44(25–62) |
21 |
51(35–77) |
14 |
|
16 |
150 |
76(50–106) |
15 |
46(29–64) |
16 |
52(38–72) |
13 |
|
III фенотип популяций |
|||||||
|
17 |
270 |
77(55–110) |
12 |
47(31–69) |
17 |
49(35–64) |
14 |
|
18 |
160 |
67(45–90) |
12 |
48(21–75) |
21 |
50(36–69) |
14 |
|
19 |
250 |
63(45–95) |
16 |
50(23–73) |
20 |
48(34–68) |
14 |
|
20 |
200 |
73(45–105) |
13 |
51(28–69) |
17 |
50(36–67) |
13 |
|
21 |
350 |
82(60–115) |
14 |
46(20–70) |
22 |
50(34–67) |
13 |
|
22 |
300 |
82(60–115) |
13 |
50(31–69) |
15 |
50(37–69) |
12 |
|
23 |
200 |
76(55–107) |
13 |
52(27–75) |
17 |
49(35–69) |
11 |
|
24 |
100 |
70(48–95) |
13 |
51(31–69) |
17 |
50(37–67) |
12 |
Примечани е: здесь и в табл. 2–4: Пункты: 1 – Выборг, 2 – Рощино (Линдуловская роща), 3 – Сортавала, 4 – Ладва, 5 – Сонково, 6 – Данилов, 7 – Иваново, 8 – Арзамас, 9 – Реболы, 10 – Костомукша, 11 – Обозерский, 12 – Коноша, 13 – Никольск, 14 – Солигалич, 15 – Шарья, 16 – Киров, 17 – Казань, 18 – Кемь, 19 – Карпогоры, 20 – Зайцево, 21 – Верхняя Тойма, 22 – Афанасьево, 23 – Кудымкар, 24 – Оса; n – число особей в выборке, шт; L c – длина шишек, мм; C n – коэффициент сужения, C p – коэффициент вытянутости верхней части семенных чешуй; X – среднее значение признака, min–max – крайние значения; C v – коэффициент вариации, %.
Показатель C n в популяциях фенотипов I и II меньше C p , а в популяциях фенотипа III они близки по своей величине.
Коэффициент вариации C n – C p составляет 39–41% при заметно большей его величине (33–50%) по отдельным популяциям. В индивидуальной изменчивости показателей C n и C p наблюдается отрицательная корреляция среднего уровня (табл. 2).
Таблица 2
Средние показатели формы семенных чешуй и их внутрипопуляционная корреляция в гибридных популяциях елей Picea abies и P. obovate
|
№ пункта |
C n –C p |
η C n и C p |
R C n и C p |
|
|
X(min–max) |
C v |
|||
|
I фенотип популяций |
||||
|
1 |
–22(–55…+11) |
44 |
0,645 |
–0,611 |
|
2 |
–23(–50…0) |
50 |
0,580 |
–0,516 |
|
3 |
–17(–57…+24) |
34 |
0,627 |
–0,616 |
|
4 |
–18(–45…+23) |
48 |
0,520 |
–0,505 |
|
5 |
–22(–60…+20) |
41 |
0,684 |
–0,679 |
|
6 |
–18(–41…+16) |
49 |
0,523 |
–0,456 |
|
7 |
–18(–51…+15) |
39 |
0,652 |
–0,647 |
|
8 |
–16(–54…+21) |
38 |
0,702 |
–0,682 |
|
II фенотип популяций |
||||
|
9 |
–13(–51…+24) |
34 |
0,575 |
–0,563 |
|
10 |
–10(–41…+22) |
45 |
0,575 |
–0,563 |
|
11 |
–6(–44…+27) |
33 |
0,551 |
–0,543 |
|
12 |
–10(–39…+18) |
45 |
0,630 |
–0,623 |
|
13 |
–9(–40…+18) |
45 |
0,652 |
–0,647 |
|
14 |
–12(–40…+21) |
40 |
0,580 |
–0,514 |
|
15 |
–7(–49…+28) |
37 |
0,715 |
–0,706 |
|
16 |
–6(–36…+23) |
31 |
0,611 |
–0,580 |
|
III фенотип популяций |
||||
|
17 |
–2(–30…+27) |
45 |
0,774 |
–0,755 |
|
18 |
–2(–42…+33) |
38 |
0,652 |
–0,627 |

|
19 |
2(–40…+40) |
35 |
0,719 |
–0,712 |
|
20 |
1(–39…+33) |
34 |
0,681 |
–0,665 |
|
21 |
–4(–46…+33) |
38 |
0,713 |
–0,702 |
|
22 |
0(–30…+30) |
40 |
0,560 |
–0,548 |
|
23 |
3(–32…+38) |
36 |
0,577 |
–0,516 |
|
24 |
1(–25…+32) |
47 |
0,514 |
–0,431 |
Примечани е: η – корреляционное отношение показателей C n и C p , R – коэффициент их корреляции.
По всем популяциям фенотипов (I, II, III) коэффициент отрицательной корреляции равен: –0,572(–0,422 … –0,734); –0,573(–0,303 … –0,706); –0,641(–0,431 … –0,779) соответственно. Во всех случаях корреляция существенно не отличается от прямолинейной (η2–R2 ˂ 0,1). Здесь, как видно, корреляция показателей C n и C p в популяциях фенотипа III в целом несколько выше. Возможно, это обусловлено более интенсивными процессами интрогрессии P. abies и P. obovata в восточной части зоны гибридизации. Внутрипопуляционная корреляция показателей C n и C p здесь в 1,25–1,40 выше, чем, например, у P. obovata в сибирской части ареала [12].
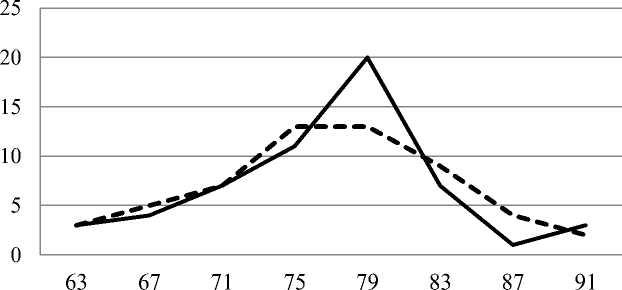
Географическая (межпопуляционная) изменчивость анализируемых показателей существенно отличается от индивидуальной (табл. 3). Во всей совокупности популяций средняя длина шишек равна 76 (63–91) мм, большее популяционное значение превосходит меньшее в 1,44 раза. В этом случае коэффициент вариации составляет 8 %. Распределение популяций по длине шишек не отличается от нормального (рис. 2).
Таблица 3 Географическая изменчивость средних показателей длины шишек и формы семенных чешуй в популяциях разных фенотипов гибридной формы елей Picea abies и P. obovata на территории России
|
Фенотип популяций |
N |
X( min–max ) |
|||
|
L c |
C n |
C p |
C n –C p |
||
|
I–III |
56 |
76(63–91) |
45(36–54) |
52(47–61) |
–7(–23…+5) |
|
I |
13 |
82(77–91) |
38(36–40) |
58(56–61) |
–20(–16…–23) |
|
II |
15 |
77(64–82) |
43(40–47) |
53(51–56) |
–10(–6…–14 |
|
III |
28 |
73(63–81) |
50(46–54) |
49(47–51) |
1(–5…+5) |
Примечание : N – число выборок.
Классы длины шишек, мм
Рис. 2. Распределение 56 популяций гибридной формы елей Picea abies и P. obovata по классам средней длины шишек. Сплошная линия – эмпирические показатели, пунктирная – выравненные
По данным Л. Ф. Правдина [21] длина шишек гибридной формы ели составляет 85±6,5 (то есть 78,5–91,5) мм. Как видно, среднее и нижнее значения имеют большие различия с нашими данными.
В популяциях фенотипа I средняя длина шишек составляет 82 мм, в популяциях фенотипа III она примерно на 12% меньше. Промежуточное значение длины шишек (77 мм) характерно для популяций фенотипа II . Размах изменчивости длины шишек в пределах фенотипа популяций примерно одинаков (63–82(91) мм). То есть, в пределах всех фенотипов встречаются популяции с довольно мелкими шишками (63–65 мм) и с относительно крупными (80–90 мм). Но во всех случаях географическая изменчивость признака в 2–3 раза ниже индивидуальной. Изменчивость показателей C n и C p между популяциями существенно меньше индивидуальной. При этом оказывается несколько больше вариация показателя C n , чем вариация показателя C p . Величина показателя C n –C p в популяциях разных фенотипов варьирует от –20 до +1 %. Этот показатель, естественно, является наиболее вариабельным.
В географической изменчивости показателей имеется большое сходство, подтверждаемое высоким уровнем корреляции (табл. 4). Коэффициент отрицательной корреляции показателей C n и C p (R= –0,90; C n = –1,2466 C p +110,93; C p = –0,649 C n +82,01; ошибка уравнений равна 0,442) значительно выше внутрипопуляционного.
Таблица 4
Сходство (корреляция) в географической изменчивости показателей формы семенных чешуй и длины шишек в популяциях гибридной формы елей Picea abies и P. obovata
|
Признаки |
η |
R |
K |
||
|
η±S η |
t |
R±S r |
t |
||
|
C n и C p |
0,934±0,0488 |
19,14 |
–0,900±0,0593 |
18,18 |
0,062 |
|
C n и L c |
0,701±0,0971 |
7,22 |
–0,654±0,1029 |
6,36 |
0,063 |
|
C p и L c |
0,629±0,1058 |
5,94 |
+0,598±0,1091 |
5,48 |
0,038 |
|
C n –C p и L c |
0,677±0,1001 |
6,76 |
–0,674±0,1006 |
6,70 |
0,040 |
Примечание. Число пар 56, η±S η – корреляционное отношение и его ошибка, R±S r – коэффициент корреляции и его ошибка, t – показатель достоверности, K(η2–R2) – показатель криволинейности связи. K 0.05 = 0,1.
Межпопуляционная (географическая) корреляция ( C n , C p и C n –C p ) и длины шишек ( Lc ) среднего уровня (R = 0,6–0,7): межу Lc и C n , C n –C p она отрицательного направления, а между C p и Lc положительного. Отсюда можно сделать вывод относительно географического изменения анализируемых признаков. Корреляции показателей длины шишек и формы семенных чешуй внутри популяций нет [25].
Наличие межпопуляционной (а по мнению некоторых авторов – и внутрипопуляционной) «связи» формы семенных чешуй с длиной шишек имеет практическое лесоводческое значение, поскольку с длиной шишек тесно связано количество и качество семян, которое влияет на рост деревьев в начальный период [2; 3; 9; 11; 17; 20; 23].
Гибридная форма елей P. abies и P. obovata характеризуется и рядом других признаков в основном промежуточного характера. Средняя масса 1000 полных семян составляет 4,5– 5,5 г, число семядолей у всходов 7–7,5 шт. [18], относительная высота семенного потомства – 125(113–138) % в популяциях фенотипов I и II, а в популяциях фенотипа III – 100(88–
113) %. Показатель внутрипопуляционного разнообразия фенотипов при их 9-классной градации характеризуется повышенным (5,1–6) и высоким (6,1–7) уровнем [38]. На территории северо-запада России, где распространена промежуточная форма ели (главным образом в Карелии), оказывается повышенной частота аллеля Gdh0.75 по сравнению с елями P. abies (Беловежская пуща) и P. obovata (Красноярск), а частота аллеля Gpi0.80 повышенной относительно первой и существенно ниже относительно второй [10]. По мнению П.А. Волковой [8], ель в Карелии по морфологическим признакам не отличается от P. obovata , а по органельной ДНК – от P. abies . Повышенная изменчивость ели здесь, как она предполагает, обусловлена естественной гибридизацией P. abies и P. obovata .
Положение популяций гибридной формы ели относительно популяций P. abies из Украинского Закарпатья и P. obovata из Восточной Сибири, используемых в качестве «стандартных» [37], определяли по C n и C p через квадрат дистанции Махаланобиса (Squared Mahalanobis Distances) [7]. Для всей их совокупности он составляет 29,44 и 18,75, то есть в целом они ближе к P. obovata ., чем к P. abies. По фенотипам популяций (I, II, III) соотношение этих показателей иное: 15,18 и 34,08; 25,02 и 22,54; 38,13 и 9,61 соответственно. Отсюда видно, что популяции фенотипа I ближе к Picea abies , а популяции фенотипа III к P. obovata . Популяции фенотипа II занимают примерно одинаковое положение относительно «стандартных» популяций елей P. obovata и P. abies . Потомство гибридной формы ели в испытательных (географических) посадках, судя по многочисленным данным, по крайней мере, для центральных районов европейской части России, отличается рядом преимуществ перед потомствами исходных елей, что весьма важно для лесоводства.
Заключение
В гибридных (переходных) популяциях елей P. abies и P. obovata на территории России наблюдаются промежуточные параметры основных систематических признаков – длины шишек и показателей формы семенных чешуй. Индивидуальная (внутрипопуляционная) изменчивость их значительно выше географической. В межпопуляционной изменчивости изученных показателей имеется значительное сходство, которое характеризуется средним уровнем корреляции. На всем протяжении полосы гибридных популяций, особенно в ее центральной части, интрогрессия прослеживается на градиенте «юго-запад – северо-восток», к северу полосы несколько усиливается широтный градиент интрогрессии, к югу – ее долготный градиент. Результаты исследования могут быть полезны для практического лесоводства, а также для решения вопросов систематики гибридной формы елей европейской и сибирской.
Работа выполнена в рамках государственного задания: проект № 121041600045 НИР ТюмНЦ СО РАН.
