Изменчивость размеров корневой системы и интенсивности микоризации у всходов сосны обыкновенной (Pinus sylvestris L.) при адаптации к условиям обитания
 при адаптации к условиям обитания")
Автор: Веселкин Д.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Почвенная микробиология
Статья в выпуске: 1 т.49, 2014 года.
Бесплатный доступ
Для выявления закономерностей изменчивости по признакам строения подземных органов, характеризующих выраженность реализации автономного и симбиотического способов адаптации эктомикоризных растений в подземной сфере, изучили всходы сосны обыкновенной ( Pinus sylvestris L.) из 45 естественных и искусственных местообитаний. На территории Уральского региона растения (всего 1496 образцов) отбирали в ненарушенных лесах, нарушенных (горелых) лесах, естественных и искусственных (питомники) нелесных местообитаниях с глинистыми, суглинистыми и торфянистыми почвами, а также выращивали в вегетационном опыте (в нестерильных условиях в теплице в вегетационных сосудах с торфяно-песчаной смесью и лесной дерново-подзолистой почвой в качестве инокулюма). Оценивали длину недетерминированных корней (индикатор автономной адаптации) и степень микоризации корневой системы (индикатор симбиотической адаптации). Индивидуальная вариабельность (изменчивость между особями в однородных условиях) по интенсивности микоризации оказалась выше, чем по длине недетерминированных корней. В наибольшей мере оба признака определяются экологическими условиями, складывающимися в зависимости от типа местообитания. На компоненту дисперсии, связанную с различием между группами по местообитанию, у признаков «длина корней» и «интенсивность микоризации» приходится соответственно 52 и 75 % общей дисперсии. Локальными модификациями экологических условий между местообитаниями в пределах групп местообитаний объясняется 14 % (длина корней) и 7 % (интенсивность микоризации) общей дисперсии. Соответственно только 34 и 18 % общей дисперсии указанных признаков объясняется индивидуальной изменчивостью. Представленные данные характеризуют эктомикоризный симбиоз как ценотический инструмент для увеличения разнообразия состояний подземных органов у эктомикоризных растений, действие которого выражается в качественных изменениях этих органов и определяется обилием, активностью, межвидовыми и внутривидовыми различиями микобионтов.
Корневая система, симбиоз, эктомикоризы, изменчивость, сосна обыкновенная pinus sylvestris
Короткий адрес: https://sciup.org/142133480
IDR: 142133480 | УДК: 631.466.12+582.475]:630*181.351
Variability of roots size and mycorrhization in seedlings of pine (Pinus sylvestris L.) under adaptation to the conditions of inhabitance
To find out the peculiarities of underground parts' architectonics variability which characterize an extent of autonomic and symbiotic adaptation of ectomycorrhizal plants in root zone, the seedlings of Pinus sylvestris L. from 45 natural and artificial localities were studied. At the territory of the Ural region the plants, 1496 samples in total, were picked out from intact forests, burnt forests, natural (arboreta) and non-forest habitats on clay, loam, and peat soils. In the experiments, the tested seedlings were also grown in greenhouse under unsterile conditions using pots filled with the mix of peat and sand to which a sample of woodland sod-podzole soil was added as an inoculum. The length of undetermined roots as an index of autonomic adaptation, and the rate of root mycorrhization as an index of symbiotic adaptation were estimated. Under similar conditions, the individual's variability on mycorrhization was shown to be higher comparing to the variability on undetermined roots length. Both these traits were mostly determined by environmental conditions depending on locality. For root length and mycorrhization rate, the component, related with differences between groups due to locality made 52 and 75 % of the total variance, respectively. Local environment modifications between the habitats within a group of habitat resulted in 14 % (root length) and 7 % (mycorrhization rate) of the total variance. So, from the total variance on these traits 34 и 18 % at most are due to individual variability. These data characterize the ectomycorrhial symbiosis as a coenotic facility to increase a diversity of ectomycorrhizal plants in root zone. Its effect results in qualitative changes in these organs and depends on plenty, activity, and inter- and intraspecial differences of mycobionts.
Текст научной статьи Изменчивость размеров корневой системы и интенсивности микоризации у всходов сосны обыкновенной (Pinus sylvestris L.) при адаптации к условиям обитания
Основной способ реализации модификационных адаптации у многолетних растений, то есть приведения морфологического строения в соответствие с условиями окружающей среды, заключается в изменении активности роста и образования новых органов (1, 2). В подземной сфере у древесных растений, помимо этого автономного механизма, действие которого выражается в дополнительном развитии корней, происходит также симбиотическая адаптация, которая морфологически проявляется в заселении детерминированных корней эктомикоризными грибами и формировании симбиотических поглощающих органов (эктомикориза). Разнообразные эффекты эктомикоризного симбиоза давно и плодотворно исследуются. В целом признано, что формирование эктомикориз приводит к изменению физиолого-биохимических параметров состояния растений и, как правило, — к их лучшему росту, что с успехом используется в практических целях (3-8).
В настоящее время преобладают исследования микоризообразова-ния, посвященные преимущественно изучению процессов их разнообразия и организации на молекулярном и биохимическом уровнях. Но для понимания экофизиологических закономерностей приспособления эктомикоризных растений к почвенным условиям необходимо знать характер изменчивости признаков, индуцирующих соответствующие механизмы их адаптации — автономные и симбиотические. Анализ варьирования признаков — не только необходимый предварительный этап в изучении био- логических объектов, но и задача, имеющая самостоятельное значение, поскольку по характеру изменчивости можно судить об адаптивной значимости и регуляции экспрессии признака (9, 10).
Цель выполненной работы заключалась в том, чтобы на примере всходов хозяйственно важного вида древесных растений (сосна обыкновенная Pinus sylvestris L.) сопоставить степень изменчивости по признакам строения подземных органов, характеризующим выраженность автономного и симбиотического способов адаптации эктомикоризных растений.
Методика . Изучали состояние корневой системы у 4-12-месячных всходов сосны обыкновенной из 45 естественных и искусственных местообитаний (выборок) ( n ) на территории Уральского региона (2004-2007 годы). Местообитания объединялись в пять групп (I-V): I — ненарушенные леса (возраст всходов 5-12 мес, n = 7) в Пригородном районе Свердловской области (сосняк наскальный, 2006 год) и Кетовском районе Курганской области (сосняк бруснично-мелкотравно-зеленомошный, 2005 год); II — нарушенные (горелые) леса (12 мес, n = 7) в Кетовском районе Курганской области (сосняк бруснично-мелкотравно-зеленомошный, 2005 год) (11); III — естественные и искусственные (питомники) нелесные местообитания с глинистыми и суглинистыми почвами (5-6 мес, n = 9), лесные питомники в Березовском, Сысертском, Ревдинском и Первоуральском районах Свердловской области (2004 год) и в Кетовском районе Курганской области (2005 год) (12), естественные местообитания в Пригородном районе Свердловской области (2005 год); IV — естественные и искусственные нелесные местообитания c торфянистыми почвами (4-6 мес, n = 5), лесные питомники в Пригородном и Сысертском районах Свердловской области (2004 год) и естественные местообитания в Пригородном районе Свердловской области (2005 год); V — вегетационный опыт (5 мес, n = 17), сеянцы сосны выращивали в 2007 году в вегетационных сосудах в теплице на территории Ботанического сада УрО РАН (г. Екатеринбург) в смеси верхового торфа и песка при разных нормах добавочного внесения азота (в форме мочевины) и фосфора (двойной суперфосфат) при оптимальной водообеспеченности; в качестве инокулюма эктомикоризных грибов использовали гумусово-аккумулятивный горизонт дерново-подзолистой почвы сосняка черничного (13). Всего проанализировано 1496 особей (по 3050 в выборке).
В качестве легко интерпретируемого признака автономной адаптивной активности использовали общий линейный размер корневой системы, для характеристики проявления симбиотических адаптаций учитывали долю детерминированных корней, трансформированных в эктомикоризы (показатель интенсивности микоризации) (14). После фиксации в 4 % растворе формалина у каждого образца определяли суммарную длину (мм) главного и боковых недетерминированных корней 1-го и 2-го порядков, а также долю (%) корней, трансформированных в эктомикоризы. Оценивали индивидуальную (между особями) и межгрупповую изменчивость признаков.
При описании индивидуального варьирования использовали показатель относительного размаха ( R ) — отношение интерлимитного размаха для выборки (местообитания) к интерлимитному размаху, наблюдаемому во всех выборках (15): R = ( X max - X mn )/( X maxABS - X minABs ) x 100, где X max и X min — соответственно максимальное и минимальное значение в выборке, X maxABS и X minABS — абсолютное максимальное и минимальное значение, зарегистрированное во всех выборках.
Для характеристики вклада в общую изменчивость признаков ис- пользовали одно- и двухфакторный дисперсионный анализ, градациями факторов в которых считали принадлежность особи к выборке и принадлежность выборки к группе местообитаний (оба фактора рассматривали как случайные); при этом разделение дисперсии выполняли по Снедекору (16). В работе также применяли непараметрический аналог двухфакторного дисперсионного анализа — расчет рангового критерия Шейрера-Рея-Хара (17). Значимость отклонения распределения признаков от нормального оценивали по критерию Шапиро-Уилка (18).
Результаты . Подход, использованный при обработке полученных данных, позволяет разделить общую дисперсию на две или три (в зависимости от схемы анализа) компоненты: долю дисперсии, обусловленную различиями между группами местообитаний; долю, которая определяется различиями между местообитаниями (выборками) в пределах групп местообитаний (межвыборочная изменчивость) и остаточную дисперсию (ее интерпретировали как индивидуальную изменчивость).
В 58 % выборок распределение по длине корней соответствовало нормальному, в 25 % — отклонения от нормального распределения были высокозначимыми (P < 0,01), что связано, как правило, с положительными значениями асимметрии и эксцесса (отличия от нуля при P < 0,05 соответственно в 36 и 28 % выборок) (табл. 1). В большей части выборок преобладали всходы с малыми размерами корневых систем.
Распределение интенсивности микоризации соответствовало нормальному только в 1 /3 выборок; отклонения от нормальности на уровне значимости 0,01 < P < 0,05 установили в 13 % выборок, на больших уровнях — в 54 % выборок. Наблюдалась как положительная (31 % выборок), так и отрицательная (24 % выборок) асимметрия распределения по интенсивности микоризации. Преобладание особей с высоким показателем обилия эктомикоризы было характерно для лесных территорий, с низким — для нелесных местообитаний.
1. Параметры индивидуального варьирования длины корней и интенсивности микоризации у всходов сосны обыкновенной Pinus sylvestris L. в зависимости от условий местообитания (Уральский регион, 2004-2007 годы)
|
Параметр |
Группа по местообитанию |
Во всех местообитаниях |
||||
|
I |
1 II |
1 III |
1 IV | |
V |
||
|
Длина корней |
||||||
|
m , мм |
97-208 |
153-253 |
244-516 |
423-752 |
499-3214 |
97-3214 |
|
SD , мм |
30-122 |
73-121 |
92-219 |
117-276 |
225-1314 |
30-1314 |
|
As |
0,94±0,18 |
0,80±0,16 |
0,41±0,17 |
0,38±0,15 |
0,61±0,15 |
0,63±0,08 |
|
Ex |
1,35±0,61 |
0,56±0,52 |
0,36±0,42 |
- 0,30±0,14 |
0,94±0,38 |
0,69±0,21 |
|
Cv , % |
44,7±4,8 |
45,1±1,3 |
35,6±2,2 |
32,4±2,3 |
44,5±2,9 |
41,5±1,6 |
|
R , % |
5,8±1,1 |
7,5±0,8 |
11,9±1,5 |
15,0±2,2 |
52,7±5,9 |
26,0±3,9 |
|
Интенсивность микоризации |
||||||
|
m , % |
73-87 |
43-63 |
11-43 |
6-9 |
1-35 |
1-87 |
|
SD , % |
8-20 |
21-32 |
9-23 |
6-12 |
2-11 |
2-32 |
|
As |
- 1,49±0,30 |
- 0,71±0,20 |
0,27±0,23 |
2,12±0,27 |
0,93±0,19 |
0,30±0,19 |
|
Ex |
3,09±1,23 |
- 0,14±0,30 |
0,12±0,36 |
5,60±1,27 |
1,15±0,57 |
1,54±0,41 |
|
Cv , % |
18,9±2,5 |
47,4±4,6 |
59,0±7,1 |
131,2±16,6 |
79,8±10,3 |
66,8±6,4 |
|
R , % |
65,3±9,5 |
91,0±2,6 |
55,5±6,8 |
38,0±4,5 |
28,9±2,5 |
50,5±4,0 |
Примечание. Описание групп по местообитанию см. в разделе «Методика»; m — среднее арифметическое, SD — среднеквадратическое отклонение (приведены пределы варьирования значений для место-обитания/группы), As — асимметрия, Ex — эксцесс, Cv — коэффициент вариации, R — относительный размах (среднее арифметическое±стандартная ошибка среднего арифметического; учетная единица — значение параметра в выборке).
Для приближения статистических распределений к нормальному оптимальными преобразованиями служили извлечение квадратного корня (длина корней) и арксинус-преобразование (интенсивность микоризации).
Анализируя данные по индивидуальной изменчивости, следует от- метить, что на основании значений коэффициентов вариации (Cv) по шкале, предложенной Г.Н. Зайцевым (19), в 93 % выборок варьирование длины корней у особей характеризовалось как нормальное или большое, в 7 % выборок — как очень большое. Нормальное и большое варьирование интенсивности микоризации наблюдали в 58 % выборок, очень большое, сверхбольшое или аномально большое — в 42 %. Судя по значениям R, в 30 из 45 выборок выявленный размах по признаку «длина корней» не превышал 25 % его общего размаха. В отношении интенсивности микоризации имелся противоположный эффект: в 22 выборках размах составлял 25-50 % от общего, в 18 — оказался выше 50 % (51-99 %) от потенциального диапазона изменчивости признака.
Индивидуальная вариабельность признаков особи различалась в зависимости от местообитания (см. табл. 1; табл. 2, строки 1-4-я). Наибольшую изменчивость размеров корневой системы отмечали в вегетационном опыте. По интенсивности микоризации максимальную величину R выявили в условиях произрастания на лесных участках, наибольшие значения Cv оказались характерными для нелесных местообитаний. Это иллюстрировали различия, оцениваемые параметрами Cv и R . При высокой активности микоризообразования при произрастании в лесу величина отношения дисперсии признака «интенсивность микоризации» к ее средним значениям ( Cv ) меньше, при низких значениях активности микоризообразования на нелесных участках и в вегетационном опыте реализуемый диапазон изменчивости уже потенциального. В среднем индивидуальная вариабельность по интенсивности микоризации, оцениваемая двумя показателями, была выше, чем по длине недетерминированных корней (см. табл. 1 и табл. 2, строки 5-6-я).
2. Сравнение величин индивидуального варьирования длины корней и интенсивности микоризации у всходов сосны обыкновенной Pinus sylvestris L. в зависимости от условий местообитания с использованием рангового критерия Шейрера-Рея-Хара (Уральский регион, 2004-2007 годы)
|
Параметр, признак |
N |
Фактор |
Взаимодействие факторов 1 х 2 ( dF = 4) |
|
|
1 (группа по местообитанию, dF = 4) |
2 (признак, dF = 1) |
|||
|
Cv , длина корней |
45 |
12,35 (0,015) |
- |
— |
|
Cv , интенсивность микоризации |
45 |
25,70 (< 0,001) |
— |
— |
|
R , длина корней |
45 |
35,15 (< 0,001) |
- |
— |
|
R , интенсивность микоризации |
45 |
28,09 (< 0,001) |
— |
— |
|
Cv , оба признака |
90 |
14,81 (0,005) |
7,19 (0,007) |
27,45 (< 0,001) |
|
R , оба признака |
90 |
2,93 (0,570) |
33,33 (< 0,001) |
37,03 (< 0,001) |
Примечание. Приведены рассчитанные значения критерия Шейрера-Рея-Хара и их уровни значимости (в скобках); N — общее число наблюдений (учетная единица — значение параметра в выборке), dF — число степеней свободы для фактора; прочерки означают, что анализировались только однофакторные планы.
Отметим, что большая индивидуальная вариабельность размеров корневых систем у всходов сосны и степени их микоризации обусловливает необходимость значительного размера выборок при проведении анализа. В частности, если для оценки длины корней с точностью 10 % требуется измерить их у 30-190 растений, то для той же точности определения интенсивности микоризации следует сравнить 10-1300 образцов. Для точности 20 % при изучении первого из указанных признаков достаточно проанализировать 15-40 особей. Такой же объем выборки дает ту же точность при оценке интенсивности микоризации в условиях леса, но в нелесных местообитаниях необходимый размер выборки уже существенно больше — 40-300 особей.
В группах по местообитанию параметры варьирования признаков между выборками были близкими (табл. 3). Сравнение значений Cv из таблиц 1 и 3 показывает, что между особями признаки варьировали сильнее, чем между выборками. Это подтвердило определение относительной величины дисперсии, обусловливаемой изменчивостью между выборками. Так, по длине корней доля межвыборочной дисперсии от общей в разных группах составляла от 10 до 49 % (в среднем 27 %), по интенсивности микоризации — от 1 до 57 % (в среднем 20 %) (см. табл. 3). Иными словами, изменчивость обоих признаков в пределах группы по местообитанию преимущественно определяется различиями между особями, но не между выборками.
3. Изменчивость длины корней и интенсивности микоризации между выборками у всходов сосны обыкновенной Pinus sylvestris L. в зависимости от условий местообитания (Уральский регион, 2004-2007 годы)
|
Признак |
Группа по местообитанию |
||
|
I I II I |
III 1 IV |
1 V |
|
|
Длина корней |
Cv ±ошибка Cv , % 29,7±8,6 17,7±4,9 |
22,9±5,7 20,8±6,9 |
51,5±10,9 |
|
Интенсивность микоризации |
6,3±1,7 12,4±3,4 |
38,5±10,3 21,8±7,2 |
70,4±17,0 |
|
М е ж в ы б |
о р о ч н ы й компонент |
дисперсии |
|
|
(доля от полной дисперсии внутри групп), % |
|||
|
Длина корней |
21,2 10,3 |
28,5 23,3 |
49,2 |
|
Интенсивность микоризации |
7,0 3,4 |
32,6 1,2 |
56,5 |
|
Примечание. Описание групп |
по местообитанию см. в разделе «Методика». |
||
Соотношение межвыборочной и индивидуальной изменчивости в разных местообитаниях специфично. По обоим признакам межвыборочная составляющая общей изменчивости была значительной на нелесных участках с глинистыми почвами и в вегетационном опыте (35-50 %), тогда как при произрастании на лесных территориях и на нелесных участках с торфянистыми почвами она заметно меньше, особенно для признака «интенсивность микоризации» (1-7 %).
Для двух изученных признаков компонента изменчивости между группами по местообитанию оказалась основной. Размахи средних значений длины корней и интенсивности микоризации в разных группах практически не перекрывались (см. табл. 1). Значения коэффициентов вариации, характеризующих изменчивость признаков между группами местообитания (по длине корней — 95,7±50,9 %; по интенсивности микоризации — 85,8±42,7 %), свидетельствуют о «сверхбольшом» варьировании согласно упоминавшейся выше классификации.
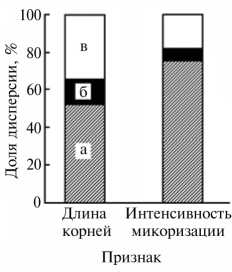
Соотношение компонент дисперсии по двум изученным признакам у всходов сосны обыкновенной Pinus sylvestris L. : а — межгрупповая, б — межвыборочная, в — индивидуальная (описание групп по местообитанию см. в разделе «Методика»; Уральский регион, 2004-2007 годы).
Компонента дисперсии, связанная с различием между группами местообитания, составляла по длине корней 52 % от общей дисперсии, по интенсивность микоризации — 75 % (рис.). Локальными различиями экологических условий между местообитаниями в пределах групп объяснялось
14 % (длина корней) и 7 % (интенсивность микоризации) общей дисперсии. Соответственно только 34 и 18 % общей дисперсии признаков определялось индивидуальной изменчивостью.
Варьирование размера корневой системы всходов и интенсивно- сти микоризации корней на любом уровне, то есть при анализе изменчивости индивидуальной, межвыборочной и между разными группами местообитаний, может быть обусловлено действием как экологических, так и генетических механизмов. Полученные в наших наблюдениях и экспериментах данные не позволяют оценить значение этих механизмов раздельно — для этого необходимы специально спланированные исследования (20, 21). В то же время для понимания закономерностей и причин изменчивости анализируемых признаков важно учитывать особенности регуляции их проявления. Образование недетерминированных (проводящих) корней осуществляется фитобионтом автономно, однако вступление в эктомикоризный симбиоз может приводить к изменению степени развития корней (22, 23). Формирование эктомикоризы определяется активностью грибов, хотя физиологическая и генетическая регуляция ми-коризообразования со стороны растения также имеет место (20, 21, 24). Факторы внешней среды способны влиять на экспрессию обоих признаков через регуляцию количества доступных энергетических и пластических ресурсов (25-30), а на микоризообразование — также посредством изменения видового состава и активности эктомикоризных грибов (3133). Именно такие значительные различия условий местообитания, связанные со степенью сформированности ценозов, в наибольшей степени детерминируют и размер корневой системы всходов, и интенсивность ее микоризации. Также вероятны генетические (популяционные) различия между растениями из разных групп местообитания по скорости роста и активности микоризообразования.
Реализованный потенциал индивидуальных симбиотических адаптаций растения в подземной сфере, определяемый внешними по отношению к особи факторами, изменяется в широком диапазоне. Индикаторный признак автономной адаптивной активности всходов менее изменчив. Из этого следует, что, независимо от внешних условий, распределение ресурсов растения в ювенильном состоянии происходит так, чтобы максимизировать эксплуатируемый объем почвы, а эктомикориза (в целом во всем рассмотренном диапазоне условий) представляет собой факультативную структуру. Однако в лесных ценозах соотношение показателей индивидуальной изменчивости по изучаемым признакам меняется на обратное. Подобный факт свидетельствует о том, что здесь эктомикоризы становятся облигатными, то есть необходимыми, конститутивными органами. Это, в свою очередь, можно расценивать как подтверждение высокой физиологической значимости эктомикоризного симбиоза в лесных местообитаниях. Представленные данные характеризуют эктомикоризный симбиоз как ценотический инструмент увеличения разнообразия состояний подземных органов у эктомикоризных растений, действие которого выражается в качественных изменениях подземных органов и определяется обилием, активностью, межвидовыми и внутривидовыми различиями микобионтов (34-36).
Таким образом, у всходов сосны обыкновенной индивидуальная вариабельность, то есть изменчивость между особями в однородных условиях, по интенсивности микоризации оказалась выше, чем по длине недетерминированных корней. В наибольшей степени значения обоих признаков определяются экологическими условиями, складывающимися в зависимости от типа местообитания. На компоненту дисперсии, связанную с различием между группами по местообитанию, у признаков «длина корней» и «интенсивность микоризации» приходится соответственно 52 и 75 % об- щей дисперсии. Локальными модификациями экологических условий между местообитаниями в пределах групп местообитаний объясняется 14 % (длина корней) и 7 % (интенсивность микоризации) общей дисперсии. Следовательно, только 34 и 18 % общей дисперсии этих признаков объясняется индивидуальной изменчивостью. В целом, на основании обсуждаемых данных эктомикоризный симбиоз следует рассматривать как ценоти-ческую адаптацию, обеспечивающую увеличение структурного разнообразия в отношении состояния подземных органов у эктомикоризных растений и приобретение лучшего соответствия между строением их поглощающего аппарата и почвенными условиями.
