Изменчивость размеров, морфометрической структуры и половой диморфизм популяций жужелицы Carabuscoriaceus в Калужской области

Автор: Алексанов В.В., Белослудцев А.Д., Суходольская Р.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 1 т.32, 2023 года.
Бесплатный доступ
Характер изменчивости размеров и динамика полового диморфизма в популяциях животных считаются индикаторами среды обитания. Мы проанализировали эти параметры в четырех популяциях жужелицы Carabus coriaceus Linnaeus, 1758, обитающих в разных по степени антропогенной трансформации локалитетах Калужской области. Жуков отлавливали почвенными ловушками, оцифровывали и обмеряли с использованием самописной компьютерной программы по шести мерным признакам - длина и ширина надкрылий, переднеспинки, головы. Всего было проанализировано 483 особи. Размеры жуков в целом наибольшие в крупном лесном массиве, наименьшие в относительно небольших и изолированных местообитаниях селитебных ландшафтов. Морфометрическая структура популяций различается статистически значимо, противопоставляется крупный лесной массив на плакоре и небольшой лес в речной долине в городе. Самки крупнее самцов по всем признакам, наиболее четко половой диморфизм проявляется по ширине надкрылий. Наибольшие значения полового диморфизма отмечены в наименьшем по площади местообитании.
Изменчивость размеров, жужелицы, морфометрическая изменчивость, половой диморфизм, фрагментация, урбанизация, речная долина, лесной вид, carabus coriaceus
Короткий адрес: https://sciup.org/148327226
IDR: 148327226 | УДК: 595.762.12:574.24 | DOI: 10.24412/2073-1035-2022-10470
Текст научной статьи Изменчивость размеров, морфометрической структуры и половой диморфизм популяций жужелицы Carabuscoriaceus в Калужской области
Введение1
Известно, что внутривидовая изменчивость признаков играет ключевую роль в динамике сообществ и функционировании экосистем (Bolnick et al., 2011). Как в глобальном, так и в локальном масштабах внутривидовая изменчивость может быть существенной, достигая магнитуды межвидовых различий (Messier et al., 2010; Kichenin et al., 2013; Des Roches et al., 2018). Поэтому представляется актуальным интегрировать внутривидовую изменчивость в модели, объясняющую и прогнозирующую тренды ответа организмов на определенные условия среды (McGill et al., 2006; Körner, 2018).
Распространенным объектом для изучения внутривидовой морфометрической изменчивости в последние десятилетия стали жуки жужелицы (Coleoptera, Carabidae). Жужелицы – признанные индикаторы среды обитания (Kotze et al., 2011). Изменчивость структуры их сообществ отражает как глобальные процессы типа изменений климата, ландшафтных, гидрологических перестроек, так и локальные, например, прямую антропогенную трансформацию местообитаний. Огромное количество работ по экологии жужелиц выполнено на уровне сообщества. В то же время известно, что процессы дивергенции, адаптации и т. п. идут на популяционном уровне. В связи с этим создан проект по морфометрической изменчивости жужелиц, представленный на Research Gates, (\Sukhodolskaya Raisa). Для некоторых видов жужелиц относительно подробно изучен половой диморфизм по морфометрическим признакам, изменчивости размеров в широтно-долготно-высотном градиентах, градиентах антропогенного воздействия и в зависимости от особенностей растительности (Benítez et al., 2020; Sukhodolskaya et al., 2020,
2021; Luzyanin et al., 2022a, b). Показано, что в широкомасштабном аспекте размеры жуков зависят от климатических факторов (Luzyanin et al., 2022a), в географических градиентах изменчивость размеров родоспецифична (Sukhodolskaya, Saveliev, 2016; Luzyanin et al., 2022b).
Carabus coriaceus Linnaeus, 1758 – крупнейшая жужелица и один из крупнейших жуков в средней полосе России, распространенный в Европе, с оптимумом в зоне широколиственных лесов. Во многих европейских странах его численность сокращается в связи с урбанизацией и интенсивным ведением сельского хозяйства (Turin et al., 2003), в России он занесен в Красные книги некоторых регионов. В Калужской области это типичный обитатель широколиственных лесов, умеренно толерантный к рекреационной нагрузке, однако не обитающий внутри городской застройки (Алексеев, 2007; Алексанов, Алексеев, 2019). C. coriaceus не летает, однако при помощи пеших миграций может перемещаться на десятки и даже сотни метров (Riecken, Raths, 1996). Среди крупных жужелиц он интересен как вид, относительно специализированный к питанию раковинными брюхоногими моллюсками (хотя потребляет также слизней, дождевых червей и личинок насекомых). Развивается он в течение одного или двух лет, зимует на стадии личинки, а во многих случаях – также на стадии неполовозрелого имаго, размножается в Калужской области в августе – сентябре (Лежнева и др., 2018). Морфометрические особенности этого яркого представителя жужелиц детально не изучались, отдельные сведения опубликованы для Южной Италии (Talarico et al., 2007), где C. coriaceus представлен другим подвидом.
Предлагаемое исследование направлено на проверку следующих гипотез: (i) самки Carabus coriaceus крупнее самцов; (ii) величины полового диморфизма по разным признакам различаются и наибольшие будут по размерам надкрылий; (iii) в селитебных ландшафтов с местообитаниями малой площади размеры жуков меньше, чем в крупных лесных массивах; (iv) в селитебных ландшафтах половой диморфизм выше, чем в крупных лесных массивах.
Материал и методика Районы исследований.
Жуки были собраны в лесных биотопах в четырех локалитетах на востоке и юго-востоке Калужской области (рис. 1).
«Дмитровский» (N 53.9244, Е 35.8415), Козельский район, окрестности хутора Дмитровский (Березичское лесничество национального парка «Угра»), верхняя часть склона водоразде- ла, ясенник вязово-дубовый с преобладание клена остролистного во втором ярусе зеленчуково-хвощово-черемшовый. Запас валежника большой. Рекреационная нагрузка отсутствует. Общая площадь лесного массива около 600 га, от других массивов он отделен лесными дорогами и просеками под ЛЭП.
« Калуга-Бор » (N 54.5224, Е 36.1745), г. Калуга, памятник природы федерального значения «Городской бор», склон южной экспозиции долины р. Оки, дубо-липняк неморальнотравный (снытевый и снытево-волосистоосоковый). Запас валежника незначительный. Рекреационная нагрузка умеренная, напочвенный покров трансформирован только вдоль тропинок. Площадь широколиственного леса около 42 га, вместе с примыкающими к нему сложными сосняками он образует массив размером около 125 га, на севере отделенный от другого массива автомагистралью, а на юге ограниченный агроценозами, залежами и рекой.
« Брагино » (N 54.4108, Е 36.6678), Пере-мышльский и Ферзиковский районы, окрестности дер. Брагино и Ладыгино, долина р. Передут – малого правого притока р. Оки, сероольшаник с черемухой птичьей крапивно-снытевый, в устье реки ивняк крапивный, с участием вяза гладкого. Запас валежника небольшой, рекреационная нагрузка на большей части массива отсутствует. Общая площадь связанных облесенных участков более 320 га, однако форма облесенной территории крайне изрезанная. Лесные биотопы окружены лугами и залежами.
« Сивцево » (N 54.6316, Е 37.0491), Тарусский район, дер. Сивцево, пологий склон к ручью, бывший усадебный парк, старовозрастный липняк с дубом неморальнотравный (снытевый и снытево-волосистоосоковый). Запас валежника большой, рекреационная нагрузка отсутствует. Площадь липняка около 1,3 га, а вместе с примыкающими к нему более молодыми фрагментами мелколиственных лесов – около 10 га, в 200–250 м от данного леса начинаются более крупные фрагменты лесов (27 га), подверженные лесохозяйственной нагрузке, в целом окружающая территория заметно трансформирована лесохозяйственной и сельскохозяйственной деятельностью.
Все локалитеты расположены в пределах от 60 до 200 км друг от друга, однако разделены обширными безлесными пространствами, крупными автомагистралями и водотоками, что создает предпосылку для рассмотрения населяющих их группировок нелетающих лесных видов в качестве отдельных популяций.
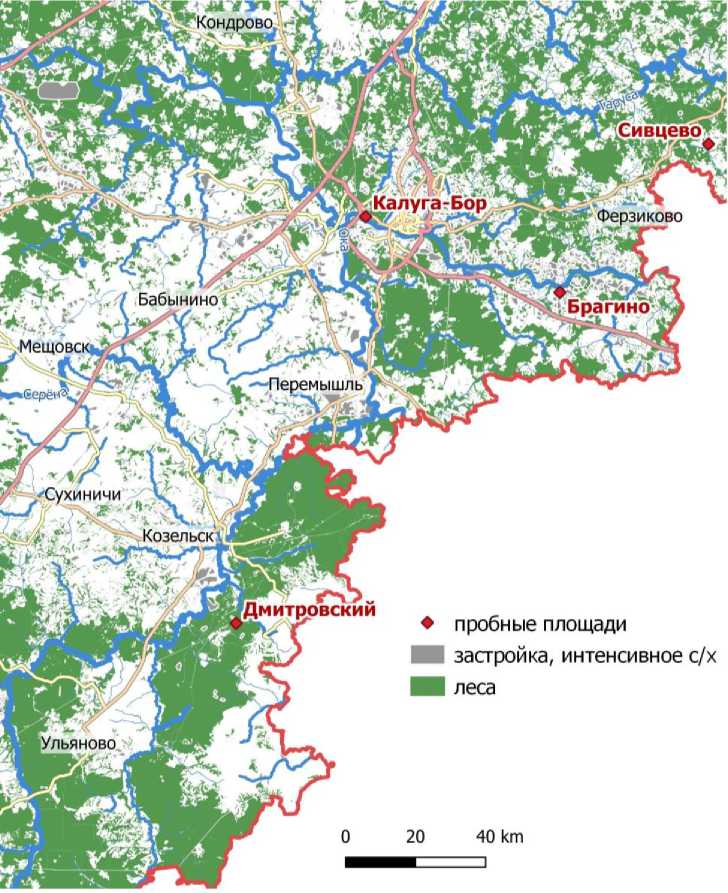
Рис. 1. Местоположение пробных площадей.
Fig. 1. Location of sample plots.
Техника исследований. Сбор жужелиц осуществлялся с помощью почвенных ловушек с раствором формалина в качестве фиксатора. Особей фотографировали по 3‒4 штуки; параметры измеряли при помощи авторской программы на Python 2.7 с использованием библиотек numpy и openCV (Мухаметнабиев, 2018), что обеспечивает автоматизацию и достижение большей точности измерений. Всего промерено 483 экземпляров насекомых по шести линейным признакам: А) длина надкрылий – расстояние по шву от середины бортика до вершины; Б) ширина надкрылий – расстояние между плечевым углом и швом, отдельно для левого и правого надкрылий; В) длина переднеспинки – расстояние по средней линии от основания до вершины; Г) ширина переднеспинки – ширина основания; Д) длина головы – расстояние от шеи до верхней губы; Е) ширина головы – расстояние между глазами.
Статистическая обработка проводилась в программах MS Excel и Statistica 10.0 с использованием параметрических методов.
Величина полового диморфизма оценивалась по формуле: (среднее самок/среднее самцов) - 1.
Результаты
Жуки Carabus coriaceus , обитающие в разных локалитетах, различаются по размерам.
По длине надкрылий для всех четырех популяций исследуемого вида жужелиц характерен половой диморфизм – самки больше самцов (рис. 2). Длина надкрылий у самок наибольшая в Калуге, на остальных трех участках она примерно одинакова. Самцы отличаются по размерам надкрылий в большей степени: они самые крупные в Брагино и Дмитровском, меньше – в Калу- ге и с самыми короткими надкрыльями – в Сивцево.
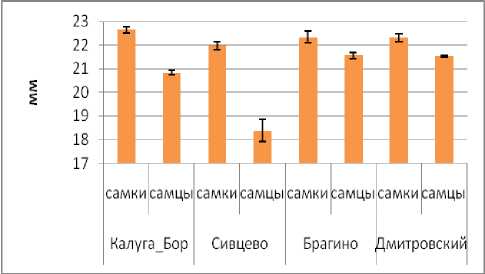
Рис. 2. Изменчивость длины надкрылий в популяциях Carabus coriaceus (обозначены средние арифметические и стандартные ошибки). Fig. 2. Variation of elytra length in Carabus coriaceus populations (means and standard errors are plotted).
Ширина надкрылий наименьшая в популяции Калуги, заметно выше в популяции Брагино и наибольшая в популяциях Сивцево и Дмитровского, в последних двух случаях она находится примерно на одном уровне (рис. 3). Обращает на себя внимание практически полное отсутствие полового диморфизма по этому признаку в первых двух популяциях, что не характерно для жужелиц, а также явно выраженная флуктуирующая асимметрия в популяции Дмитровского.
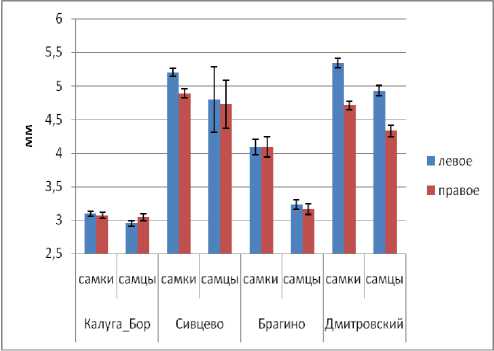
Рис. 3. Изменчивость ширины надкрылий в популяциях C. coriaceus (обозначения см. рис. 2).
Fig. 3. Variation of elytra width in C. coriaceus populations (designation see Fig. 2).
Длина и ширина переднеспинки у самок меньше в Сивцево по сравнению с тремя другими популяциями. У самцов переднеспинка значимо мельче в Дмитровском по сравнению с Калугой и Брагино (рис. 4). Обращает на себя внимание инвертированный половой диморфизм по параметрам переднеспинки (у самцов она крупнее) в популяции Сивцево.
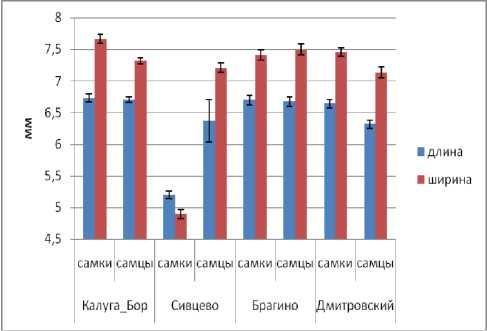
Рис. 4. Изменчивость размеров переднеспинки в популяциях C. coriaceus (обозначения см. рис. 2).
Fig. 4. Variation of pronotum size in C. coriaceus populations (designation see Fig. 2).
Длина головы у самок оказалась наибольшей в популяции Сивцево, несколько меньше она в популяции Дмитровского и значимо меньше в Калуге и Брагино. Среди самцов наибольшую длину головы демонстрируют жуки из Дмитровского, в остальных популяциях длина головы значимо меньше. Ширина головы у самок также наибольшая в Сивцево (рис. 5). У самцов же ширина головы примерно одинакова во всех популяциях.
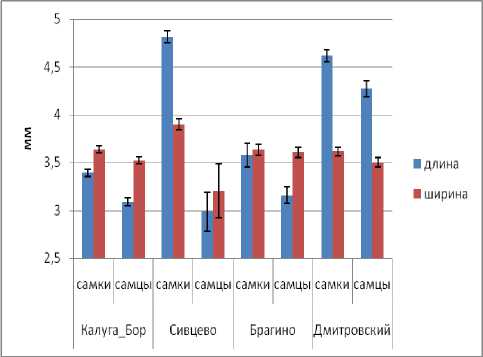
Рис. 5. Изменчивость размеров головы в популяциях C. coriaceus (обозначения см. рис. 2).
Fig. 3. Variation of head size in C. coriaceus populations (designation see Fig. 2).
Поскольку изложенные выше признаки представляют собой разные характеристики размеров жука, для анализа изменчивости размеров в целом мы упорядочили исследованные популяции по возрастанию размеров каждого признака (табл. 1). В левой части таблицы (наименьшая 45
величина признака и следующая за ним) популяции из Калуги и Сивцево упомянуты в 10 и 8 случаях соответственно. В правой части таблицы (наибольшая величина признака и следующая за ним) чаще всего - в 11 случаях - упоминается
Дмитровский. Брагино примерно одинаково распределяется по каждому из четырех мест по разным признакам. Следовательно, в целом размер жуков C. coriaceus меньше в Калуге (Бор) и Сивцево по сравнению с Дмитровским.
Таблица 1
Анализ характера изменчивости размеров жуков в популяциях Carabus coriaceus Patterns of variation of body size in Carabus coriaceus populations
|
Признак |
Место популяции по возрастанию величины признака |
|||
|
Самки |
1 |
2 |
3 |
4 |
|
Длина надкрылий |
Дмитровский< |
Сивцево= |
*Брагино< |
Калуга |
|
Ширина надкрылья левого |
*Калуга< |
*Брагино< |
*Сивцево< |
* Дмитровский |
|
------------------------- правого |
*Калуга< |
*Брагино< |
*Дмитровский< |
* Сивцево |
|
Длина переднеспинки |
Сивцево< |
Калуга= |
Брагино= |
Дмитровский |
|
Ширина переднеспинки |
Сивцево< |
Брагино= |
Дмитровский< |
Калуга |
|
Длина головы |
Брагино< |
Калуга< |
Дмитровский< |
Сивцево |
|
Ширина головы |
*Калуга= |
Брагино= |
*Дмитровский= |
Сивцево |
Самцы
|
Длина надкрылий |
Сивцево < |
Калуга < |
*Брагино= |
Дмитровский |
|
Ширина надкрылья левого |
*Калуга < |
*Брагино < |
*Сивцево= |
*Дмитровский |
|
------------------------- правого |
*Калуга= |
*Брагино < |
*Дмитровский= |
*Сивцево |
|
Длина переднеспинки |
Дмитровский < |
Сивцево= |
Калуга= |
Брагино |
|
Ширина переднеспинки |
Сивцево= |
Дмитровский < |
Калуга= |
Брагино |
|
Длина головы |
Калуга= |
Сивцево= |
Брагино < |
Дмитровский |
|
Ширина головы |
*Калуга= |
Сивцево= |
*Дмитровский < |
Брагино |
Полужирным шрифтом отмечены случаи сопряженной изменчивости длины и ширины того или иного отдела тела.
Знаком * обозначены случаи сопряженной изменчивости признака у самцов и самок (одно и то же место).
По морфометрической структуре исследованные популяции C. coriaceus различаются статистически значимо (табл. 2, рис. 6), с уровнем достоверности p < 0.001. Наиболее четко противопоставлены друг другу популяции Калуга-Бор и Дмитровский.
Таблица 2
Расстояние Махаланобиса между центроидами распределения признаков в изученных популяциях C. coriaceus
Mahalanobis distance between centroids of variable distribution in C. coriaceus populations
|
Локалитеты |
Брагино |
Калуга_Бор |
Сивцево |
|
Брагино |
- |
||
|
Калуга_Бор |
4.348 |
- |
|
|
Сивцево |
6.163 |
18.058 |
- |
|
Дмитровский |
10.369 |
24.465 |
0.874 |
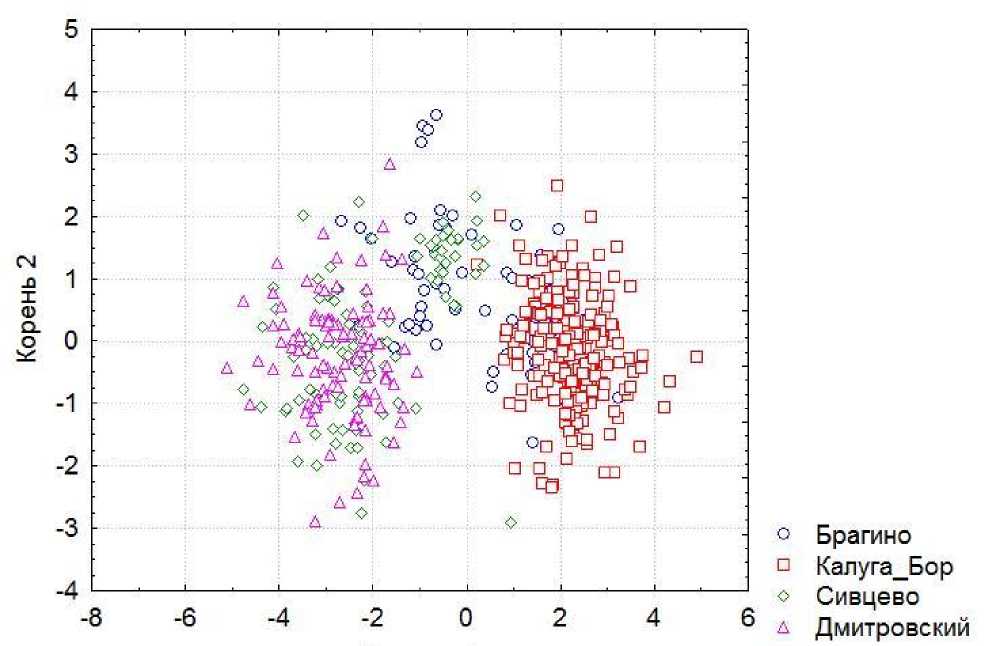
Корень 1
Рис. 6. Распределение популяций C. coriaceus в плоскости двух дискриминантных осей (Wilks' Lambda: 0.1556730 approx. F (21.1358) = 58.95957 p < 0.0001).
Fig. 6. Distribution of studied C. coriaceus populations along the first and second discriminant axes (Wilks' Lambda: 0.1556730 approx. F (21.1358) = 58.95957 p < 0.0001).
Как видно из табл. 1, в ряду исследованных популяций у самцов и самок полностью согласованно меняется только ширина левого и правого надкрылий и частично согласованно (совпадают 1-е и 3-е места, а 2-е и 4-е различаются) – ширина головы. Это свидетельствует о неодинаковом проявлении полового диморфизма в разных популяциях.
Половой диморфизм неодинаково выражен как по разным признакам, так и для разных популяций. При усреднении значений признаков по всем выборкам выявлено, что наиболее выражен половой диморфизм по ширине надкрылий и длине головы, наименее выражен по размерам переднеспинки (рис. 7).
Среди изученных популяций наибольший половой диморфизм по длине надкрылий выявлен в Сивцево, по ширине надкрылий – в Брагино, по длине и ширине головы – в Брагино (рис. 8). При усреднении значений полового диморфизма по всем шести измеренным признакам (рис. 9) более высокий уровень полового диморфизма обнаружен в Сивцево по сравнению с Дмитровским и Калугой (в Брагино в силу неодинаковой выраженности полового диморфизма по разным признакам его усредненная величина характеризуется очень большой стандартной ошибкой).
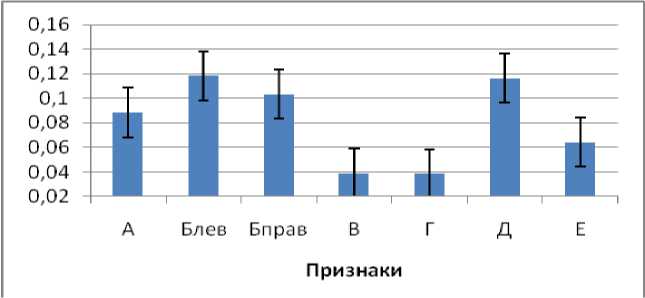
Рис. 7. Усредненные по всем выборкам значения полового диморфизма по разным признакам у C. coriaceus: А – длина надкрылий, Б – ширина надкрылий левого и правого, В – длина переднеспинки, Г – ширина переднеспинки, Д – длина головы, Е – ширина головы.
Fig. 7. Average values of sexual size dimorphism in C. coriaceus among variables: А – elytra length, Б – elytra width (left and right), В – pronotum length, Г – pronotum width, Д – head length, Е – head width.
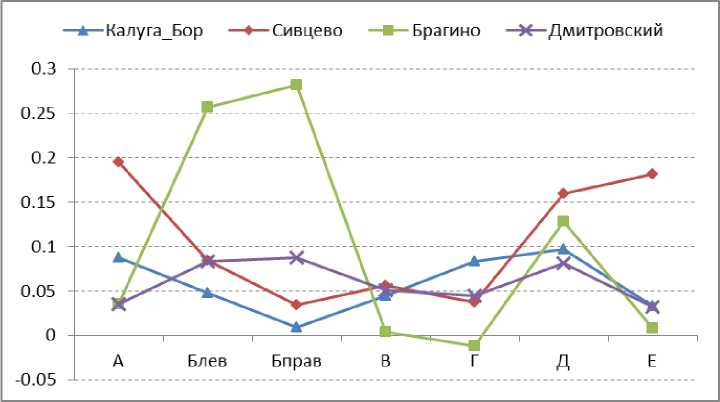
Рис. 8. Значения полового диморфизма по разным признакам в популяциях C. coriaceus. Обозначения см. рис. 7.
Fig. 8. Values of sexual size dimorphism in C. coriaceus for different population among variables. Designation see Fig. 7.
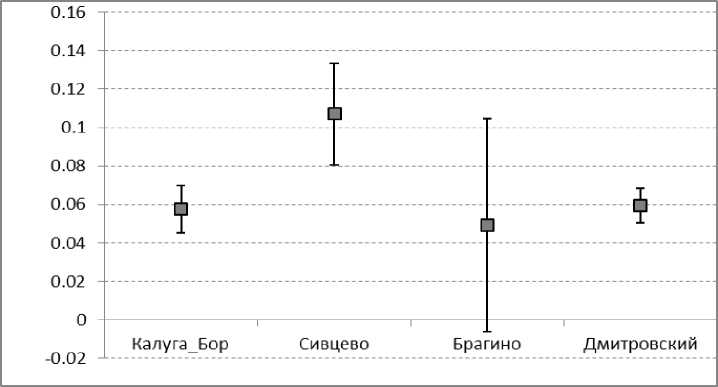
Рис. 9. Усредненные (по шести измеренным признакам) величины полового диморфизма в популяциях C. coriaceus.
Fig. 9. Average (resulting six variables) values of sexual sixe dimorphism for different population of C. coriaceus.
Обсуждение
Полученные результаты относительно полового диморфизма у C. coriaceus согласуются со сведениями о других изученных видах жужелиц — для этой группы характерен female-biased половой диморфизм, т. е., самки крупнее самцов (Sukhodolskaya et al., 2016), что имеет вполне понятный биологический смысл в силу разных ролей самцов и самок. Ширина надкрылий как наиболее подверженный половому диморфизму признак и размеры переднеспинки как наименее подверженные половому диморфизму показатели также известны для других видов жужелиц (Воробьева и др., 2022; Суходольская и др., 2022). Четкий половой диморфизм демонстрирует и длина надкрылий. Примечательно, что в Южной Италии половой диморфизм по длине тела у этого вида не оказался значимым (Talarico et al., 2007). Отчасти эти различия могут быть объяснены меньшей чувствительностью длины тела в целом по сравнению с промерами цельных склеротизированных структур в нашей работе, но более вероятно, что они вызваны таксономическими и экологическими различиями изученных жуков: в Южной Италии обитает другой подвид данного вида, населяющий открытые биотопы.
Размеры животных , как правило, уменьшаются в неблагоприятных условиях среды, складывающихся в нарушенных биотопах (Gray, 1989). В лесном поясе России для большинства видов жужелиц выявлено уменьшение размеров особей при антропогенном воздействии, в частности, при урбанизации, а также при пирогенных нарушениях, хотя для некоторых видов показано противоположное действие (Тимофеева, 2010; Sukhodolskaya, 2013; Sukhosolskaya et al., 2020; Ухова, Суходольская, 2021). В большинстве работ изучаются эвритопные виды жужелиц, гене-ралисты. В представляемой статье рассмотрен более стенобионтный вид, да еще крупных размеров. Полученные результаты достаточно четко демонстрируют снижение размеров C. coriaceus в биотопах селитебных ландшафтов по сравнению с крупным лесным массивом. Конкретные механизмы уменьшения размеров жуков в городах описываются по-разному (Magura et al., 2020; Lövei, Magura, 2022): город как остров тепла благоприятствует более мелким жужелицам, уменьшение размеров при недостатке пищевых объектов и т. д. В изученных нами популяциях C. coriaceus в Калужской области уменьшение размеров жуков, по-видимому, в первую очередь связано с фрагментацией местообитаний: малые размеры жужелиц в Сивцево и Калуга-Бор вызваны не столько качеством среды (напочвенный 49
покров, подстилка, пищевые объекты), сколько малыми размерами участка местообитания при затрудненной или невозможной миграции. Размер участка местообитания и качество окружающего его матрикса, по-видимому, в изученных популяциях отчасти компенсируют друг друга: в Сивцево площадь местообитания крайне мала, но окружающие луга, залежи и мелколесья не столь жестко затрудняют миграцию жуков из других лесных массивов, а широколиственный лес Калуга-Бор относительно крупнее, но более жестко изолирован от других лесных массивов.
По морфометрической структуре наиболее четко противопоставлены друг другу популяции C. coriaceus из Дмитровского и Калуга-Бор, а обитатели Сивцево и Брагино заняли промежуточную позицию, при этом для Калуга-Бор и Брагино общим являются более узкие надкрылья у обоих полов, а также короткая голова у самок. Нам представляется, что в морфометрической структуре проявляются различия между популяциями долины реки Оки с ее притоками и пла-корными популяциями. Механизм формирования наблюдаемых различий и их адаптивное значение требуют дальнейшего изучения. Известно, что в большинстве случаев внутривидовые морфометрические различия у жужелиц рода Cara-bus селективно нейтральны, но иногда они адаптивны по отношению к пищевым объектам (Turin et al., 2003). При морфометрических исследованиях жужелиц г. Казани показано, что различия между популяциями могут заключаться не только в размерах, но и в морфометрической структуре, и выдвинут тезис о том, что на популяционном уровне жуки приспосабливаются к условиям среды двумя способами – или изменением фактических размеров особей, или изменением морфометрической структуры (Тимофеева, 2010).
В случае C. coriaceus в Калужской области относительно давнее расхождение плакорных и долинных популяций представляется весьма вероятным в силу интенсивного сельскохозяйственного использования речных долин, в пределах которых лесные виды жуков могли сохраниться только на небольших лесных «островах». История расселения C. coriaceus в Калужской области не изучена. В начале ХХ века В.Н. Лучник (1910) характеризует C. coriaceus как очень редкий, «по-видимому, вымирающий» вид. Это утверждение наряду со сведениями об интенсивном сельскохозяйственном использовании Окской долины и ростом лесистости региона в 1940-е гг. (Люри и др., 2010) позволяет предполагать, что многие местообитания были освоены C. coriaceus менее 100 лет. К таковым относится, вероятно, местообитание Брагино, на что указывает относительная молодость древостоя. Однако значимые морфометрические различия в популяциях жужелиц могут формироваться даже за несколько десятилетий (Turin et al., 2003). В то же время леса Калужского городского бора и парк в Сивцево существуют гораздо больше столетия, и очень вероятно, что C. coriaceus обитает здесь очень длительное время, а его крайняя редкость в начале ХХ в. отчасти объясняется неполнотой территориального охвата региона энтомологическими исследованиями и преимущественно ручным сбором, который для данного вида малоэффективен (Алексанов, Алексеев, 2019).
Помимо исторических различий, долинные местообитания C. coriaceus очевидно более теплые и сухие по сравнению с плакорными. Обычно считается, что распространение данного вида лимитировано недостатком пищевых объектов (Turin et al., 2003), но в данном случае в локальном масштабе, вероятно, на первый план в долинных биотопах выходит недостаток убежищ для такого крупного жука. В связи с этим не исключена адаптивность более узкого тела к убежищам долинных биотопов.
Половой диморфизм C. coriaceus оказался неодинаково выраженным в разных популяциях. В суровых условиях среды его значение теоретически должно возрастать (Геодакян, 1991), что было доказано в ряде исследований для жужелиц (Суходольская, Савельев, 2017аб). Повышенное значение полового диморфизма в среднем по всем признакам в популяции C. coriaceus из Сивцево указывает на неблагоприятные условия для данного вида в этом небольшом биотопе.
Заключение
По результатам исследования можно сделать следующие выводы.
Для Carabus coriaceus , как и для большинства жужелиц, характерен female-biased половой диморфизм, т. е., самки крупнее самцов.
Наиболее выражен половой диморфизм по ширине надкрылий, наименее выражен по размерам переднеспинки.
Размеры жуков в местообитаниях малой площади в селитебных ландшафтах меньше, чем в крупных лесных массивах.
Половой диморфизм выше в местообитаниях малой площади в селитебных ландшафтах по сравнению с крупным лесным массивом.
По морфометрической структуре противопоставляются популяции крупного плакорного лесного массива и небольшого изолированного лесного массива в долине реки.
Представляемая работа является первым анализом морфометрической изменчивости у C coriaceus . Полученные результаты подтверждают данные, полученные раннее в этом направлении по другим видам жужелиц. Значимость работы заключается в анализе относительно стенотопного вида – типичного обитателя зональных широколиственных лесов, уязвимого в ряде регионов. Дальнейшее изучение этого интересного вида предполагает как расширение сети пунктов учета, так и интеграцию морфометрических исследований с изучением особенностей жизненного цикла, связей с другими видами и другими подходами и методами.
За помощь в сборе материала авторы признательны сотрудникам отдела мониторинга биоразнообразия ГБУ КО «Дирекция парков» С.К. Алексееву, М.И. Гаркунову, С.Е. Карпухину, Д.В. Хвалецкому, а также учащимся школ г. Калуги В.В. Беширу и Я.А. Кускову.
Список литературы Изменчивость размеров, морфометрической структуры и половой диморфизм популяций жужелицы Carabuscoriaceus в Калужской области
- Алексанов В.В., Алексеев С.К. Кадастр жуков жужелиц (Coleoptera, Carabidae) городского округа «Город Калуга». Ижевск: ООО «Принт», 2019. 276 с.
- Алексеев С.К. Пространственно-экологическая характеристика жужелиц (Coleoptera: Carabidae) лесов Калужской области: Автореф. дис. … канд. биол. наук. Калуга, 2007. 22 с.
- Воробьева И.Г., Алексанов В.В., Лузянин С.Л., Савельев А.А., Суходольская Р.А. Факторы, влияющие на половой диморфизм по размерам у жуков-жужелиц (Coleoptera, Carabidae) в крупных городах // Современные проблемы биологической эволюции: материалы IV Международной конференции к 875-летию Москвы и 115-летию со дня основания Государственного Дарвиновского музея. 17–20 октября 2022, г. Москва. М.: ГДМ, 2022. С. 434-436.
- Геодакян В.А. Эволюционная теория пола // Природа. 1991. № 8. С. 60-69.
- Лежнева В.В, Алексанов В.В., Алексеев С.К. К биологии жужелицы Carabus coriaceus L. в заповеднике «Калужские засеки» // Экологические и биологические основы повышения продуктивности и устойчивости природных и искусственно возобновленных лесных экосистем: материалы Междунар. науч.-практ. конф., посвящ. 100-летию высшего лесного образования в г. Воронеж и ЦЧР России 4–6 октября 2018 г. Т. 1. Воронеж, 2018. С. 158-165.
- Лучник В.Н. Заметки о скакунах и жужелицах русской фауны (Coleoptera, Cicindelidae et Carabidae). Русское энтомологическое обозрение. 1910. Т. 10, № 3. С. 140-143.
- Люри Д.И., Горячкин С.В., Караваева Н.А., Денисенко Е.А., Нефедова Т.Г. Динамика сельскохозяйственных земель России в XX веке и постагрогенное восстановление растительности и почв. Москва: ГЕОС, 2010. 415 с.
- Суходольская Р.А., Савельев А.А. Географическая изменчивость полового диморфизма у жужелицы Carabus granulatus L. (Coleoptera, Carabidae) // Рос. журн. прикладной экологии. 2017а. № 4. С. 3-10.
- Суходольская Р.А., Савельев А.А. Половой диморфизм по размерам жужелицы Carabus cancellatus Ill. (Coleoptera, Carabidae) // Вестн. Оренбург. пед. ун-та. 2017b. Т. 1. С. 49-64. Суходольская Р.А., Лузянин С.Л., Воробьева И.Г., Алексанов В.В., Минязова В.Б. Изменчивость размеров и половой диморфизм жужелицы Pterostichus melanarius Ill. в городах // Рос. журн. прикладной экологии. 2022. № 1. С. 15-22.
- Тимофеева Г.А. Морфометрическая структура популяций жужелиц (Coleoptera, Carabidae) в антропогенных ландшафтах. Дис. …. канд. биол. наук. Казань, 2010. 167 с.
- Ухова Н.Л., Суходольская Р.А. Изменчивость размеров и морфометрической структуры жужелиц (Coleoptera, Carabidae) при разной степени нарушенности естественного ландшафта // Проблемы трансформации естественных ландшафтов в результате антропогенной деятельности и пути их решения: сб. науч. тр. по материалам Междунар. науч. экол. конф. Краснодар: КубГАУ, 2021. С. 702-705.
