Изменение активности каталазы и пероксидаз в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды

Автор: Четина Оксана Александровна, Боталова К.И., Мошева Виктория Андреевна, Лучникова Ксения Игоревна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 4, 2018 года.
Бесплатный доступ
Изучены некоторые физиолого-биохимические показатели в листьях овса посевного Avena sativa L. при совместном воздействии засоления и разных рН-уровней корневой среды. Через 30 мин. отмечено увеличение содержания перекиси водорода на всех опытных вариантах. Активность каталазы и пероксидаз усилилась при отдельном действии исследуемых факторов корневой среды. На фоне комбинированного стресса в листьях растений не выявлено существенного увеличения активности этих антиоксидантных ферментов. Возможно, разбалансировка между АФК-генерирующими системами и системами антиоксидантной защиты при сочетании неблагоприятных условий корневой среды связана с недостаточным количеством времени для формирования адаптивного ответа. Наибольшее негативное влияние на ростовые показатели овса оказывало сочетание засоления и кислой реакции корневой среды.
Активные формы кислорода, каталаза, пероксидазы, окислительный стресс, адаптация
Короткий адрес: https://sciup.org/147227051
IDR: 147227051 | УДК: 581.1 | DOI: 10.17072/1994-9952-2018-4-423-429
Change of the catalase and peroxidases activity in leaves of cultivated oats under the influence of the separate and combined impact of salinization and pH-level of the root environment
Some physiological-biochemical indicators in leaves of cultivated oat Avena sativa L are studied. at joint impact of salinization and different рН-levels of the root environment. After 30 min. increase in content of peroxide of hydrogen on all skilled options was indicated. The activity of a catalase and peroxidases has amplified at separate action of the studied factors of the root environment. Against the background of the combined stress in leaves of plants essential increase in activity of these antioxidant enzymes isn't revealed. Perhaps, disbalance between AFK-generate systems and the systems of antioxidant protection at a combination of adverse conditions of the root environment is connected with insufficient amount of time for formation of the adaptive answer. The greatest negative impact on growth indicators of oats was exerted by a combination of salinization and sour reaction of the root environment.
Текст научной статьи Изменение активности каталазы и пероксидаз в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды
Среди природных стрессоров засоление занимает одно из ведущих мест. По оценке FAO (Food and Agriculture Organization) засоленными являются 22% мировых земель, находящихся в сельскохозяйственном использовании [Гарифзянов, Жуков, 2013]. На протяжении долгого времени адаптацию растений к засолению рассматривали через призму регуляторных механизмов поддержания ионного и осмотического гомеостаза. Однако, как известно, повышенная концентрация NaCl приводит к окислительному стрессу, сопряженному с избыточной генерацией ак- тивных форм кислорода (АФК), таких как супероксидный анион радикал (О2•-), перекись водорода (Н2О2) и гидроксильный радикал (•ОH) [Meneguzzo, Navani-Izzo, Izzo, 1999; Mittler, 2002; Полесская, Каширина, Алехина, 2006; Гарифзянов, Жуков, Ивани-щев, 2011]. Наряду с выполнением необходимых сигнальных или иных функций, АФК, как высокореактивные вещества, способны наносить вред биологическим структурам клетки и физиологобиохимическим процессам, протекающим в ней.
Антиоксидантная защитная система клетки растения – это множество взаимосвязанных окислительно-восстановительных реакций, в которых участвуют антиоксидантные ферменты и низкомо-
лекулярные метаболиты. В нормальных условиях и при окислительном стрессе супероксиддисмутазы, пероксидазы, каталаза и ферменты аскорбат-глутатионового цикла, играют важную роль в поддержании определенного безопасного уровня АФК. В ответ на усиление генерации АФК, как правило, наблюдается активация элементов антиоксидантной защитной системы. [Sies, 1997; Колупаев, 2007; Jones, 2008 и др.].
В почвах накопление солей может сопровождаться увеличением щелочности, техногенное засоление развивается и на фоне кислой реакции среды [Еремченко, Митракова, Шестаков, 2017]. В свою очередь, для большинства сельскохозяйственных культур, в особенности для молодых растений, требуется благоприятная среда, близкая к pH 6.5. Следовательно, актуальность изучения механизмов адаптации растений к совместному воздействию засоления и кислотности/щелочности обусловлена тем, что эти факторы сочетаются в природных и техногенных почвах. В современной физиологии растений недостаточно изучена ответная реакция растений на комбинированное воздействие этих факторов на состояние корневой среды, в том числе, участие ферментов антиоксидантной защиты в устойчивости растений.
Цель работы – изучить изменение активности каталазы и пероксидаз в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды.
Объекты и методы исследования
В качестве объекта исследования использовали овес посевной Avena sativa L. Предварительно замоченные семена высаживали в вермикулит. На 7й день после посадки на корневую среду воздействовали стресс-факторами: 100 мМ NaCl, 200 мМ NaCl, pH 3, pH 10, 100 мМ NaCl + рН 3, 100 мМ NaCl + рН 10, 200 мМ NaCl + рН 3, 200 мМ NaCl + рН 10. Щелочная среда создавалась буфером глицин-KOH (pH10), кислая – уксусной кислотой. Устойчивость реакции среды в вермикулите под воздействием щелочных и кислых растворов контролировали путем измерения на рН-метре в течение 48 ч. Контрольный полив производили дистиллированной водой. Через 30 мин. после воздействия стресс-фактором определяли содержание пероксида – по ферротиоцианатному методу [Sagisaka, 1976], пероксидазную функцию в растительном материале по методу А.Н. Бояркина [Ермаков и др., 1987], активность каталазы – титрометрическим методом по А.Н. Баху и А.И. Опарину [Воскресенская, Алябышева, По-ловникова, 2006]. Биологическая и аналитическая повторность определений – трехкратная. Ростовые показатели определяли через 24 и 48 ч. в 30- кратной повторности. Полученные данные обработаны методами статистики, регрессионного и дисперсионного анализов.
Результаты и их обсуждение
Общим интегральным процессом, характеризующим негативное действие стрессоров различной природы, является усиление генерации активных форм кислорода. Пероксид водорода, обладающий наибольшим временем жизни (около 1 мс) и способный диффундировать от места образования, является одним из важнейших кислородных редуктантов, который в клеточных компартментах приводит к окислению липидов, углеводов, белков, повреждению ДНК и РНК, дезорганизации цитоскелета [Foyer, Noctor, 2005; Полесская, 2007; Pol-jsak, Milisav, 2012; Miura, Tada, 2014; Радюкина 2015].
Результаты нашего эксперимента показали, что на всех опытных вариантах после воздействия стресс-факторов наблюдалось повышенное содержание пероксида водорода в листьях овса в сравнении с контролем (рис. 1). Наибольшее количество пероксида зафиксировано при действии отдельных факторов: растворов с рН3 и рН10 и засоления NaCl 100мМ, содержание его в растениях было в 8–10 раз выше контрольного уровня. При сочетании засоления и кислотности, засоления и щелочности корневой среды, особенно растворов с NaCl 200мМ, количество перекиси было ниже опытных вариантов с действием отдельных факторов, что может быть обусловлено функционированием антиоксидантной системы.
В последнее время активно обсуждается вопрос о способности АФК выступать в качестве сигнальных молекул и регуляторов экспрессии генов, детерминирующих защитный ответ растения. Определенный уровень АФК необходим для протекания ряда метаболических реакций в клетке и не вызывает повреждений биомолекул. Образование повышенного количества АФК опасно в случае нарушения баланса их образования и разрушения. Наиболее распространенными ферментами, способными детоксифицировать пероксид, являются каталаза и большое семейство пероксидаз [Blokhina, Virolainen, Fagerstedt, 2003; Mittova et all., 2003; Apel, Hirt, 2004; Mittler et al., 2004; Gechev et al., 2006; Полесская, 2007; Abogadallah, 2010, Poljsak, Milisav, 2012; Радюкина 2015; Foyer, Noctor, 2015].
В ходе эксперимента в надземной части овса посевного при разном уровне засоления корневой среды происходило достоверное увеличение активности каталазы (на 24–33%) по сравнению с контролем (рис. 2). Увеличение активности этого фермента (на 21%) отмечено и после воздействия кислого раствора без засоления. На остальных ва- риантах опыта, сочетающих оба стрессовых фактора корневой среды, не было значимых различий с контролем по активности каталазы.
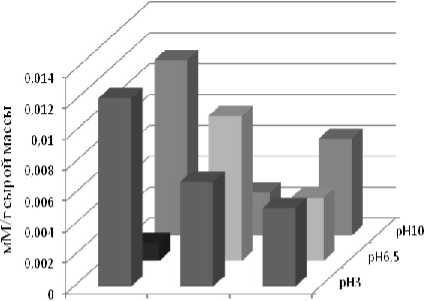
ОмМ ЮОглМ ZOOmM
концешрадй NaCl и pH кнесимеи' рас спора
Рис. 1 . Содержание перекиси в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды (черным цветом выделен контрольный вариант опыта)
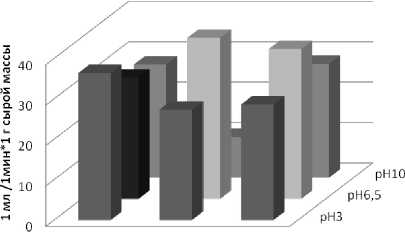
О ЮОмМ 200мМ
Рис. 2 . Активность каталазы в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды (черным цветом выделен контрольный вариант опыта)
Сущность каталитического действия каталазы состоит в разложении перекиси водорода с выделением молекулярного кислорода. Данная реакция протекает с очень высокой скоростью, одна молекула фермента способна вызвать распад 6 x 10 6 молекул перекиси водорода в секунду. В то же время каталаза имеет низкое сродство к субстрату (H 2 O 2 ) и начинает работать только при достаточно высоком содержании перекиси. В связи с этим, а также, поскольку каталаза практически отсутствует в ряде компартментов клетки, существует необходимость функционирования других ферментов, задействованных в детоксикации перекиси водорода [Рогожин, 2004; Гарифзянов, Жуков, Иванищев, 2011].
Активность пероксидаз в листьях овса относительно контроля возрастала на 50–60% при действии отдельного засоления корневой среды (рис. 3). Близкое к этому усиление пероксидазной функции наблюдалось и при отдельном воздействии рН-уровня корневой среды: на 52 – на кислой и на 57% – на щелочной среде. При сочетании факторов засоления и кислотности/щелочности корневой среды существенных различий по активности пероксидаз в надземных органах овса посевного относительно контроля через 30 мин. после воздействия не обнаружено.
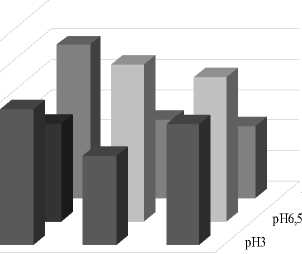
100мМ 200мМ
Рис. 3 . Пероксидазная функция в надземной части овса под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды (черным цветом выделен контрольный вариант опыта)
Наш эксперимент подтверждает сбалансированность окислительно-восстановительных реакций в растительных клетках, т. к. были выявлены прямые зависимости между содержанием перекиси (y 1 ) и активностью каталазы (х 1 ), а также между содержанием перекиси и активностью пероксидаз (y 2 ) в листьях овса посевного (х 2 ): y 1 = 23.91+924.7х 1 ; коэффициент корреляции R=0.37; уровень значимости нулевой гипотезы P=0.0005; y 2 = 27.26 + 1565x 2 ; R=0.5; Р=1.2 х 10-6.
Наиболее общим проявлением и первым симптомом действия стрессоров является подавление роста и развития растений. В природе степень устойчивости к действию стрессора часто характеризуется обратной корреляцией со скоростью роста. Медленный рост позволяет растениям выживать при стрессе, так как освобождает множество ресурсов (строительных блоков и энергии), необходимых для реализации защитной программы [Алехина и др., 2005; Deinlein et al., 2014].
Данные нашего исследования показали, что высота надземной части растений овса посевного через 24 ч. после воздействия растворов с разным рН-уровнем (без засоления) достоверно ниже контрольного варианта. На кислой среде прирост длины надземной части тормозится на 75% в сравнении с контролем, на щелочной – на 38% (рис. 4). Через 48 ч. после воздействия стресс-фактора существенные различия по приросту высоты надземной части наблюдаются только между контрольным вариантом и рН 3 (меньше контроля на 37%).
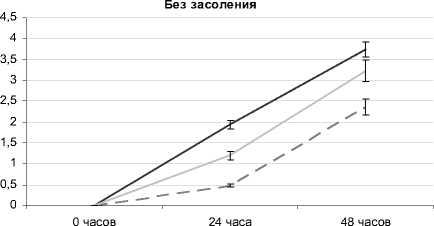
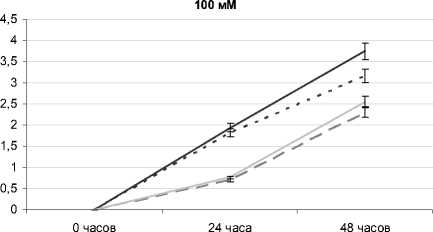
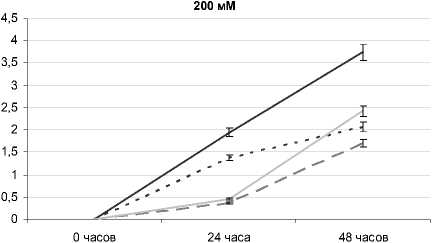
контроль 200мМ 200мМ+рН3 200мМ+рН10
Рис. 4 . Высота надземной части растений овса посевного при отдельном и комбинированном воздействии засоления и разных рН-уровней корневой среды
При отдельном воздействии засоления 100 мМ через 24 ч. значимых различий по приросту с контролем не обнаружено, через 48 ч. отмечено торможение на 16%. При комбинированном действии засоления 100 мМ NaCl с разными рН-уровнями корневой среды надземная часть растений так же замедляет свой рост по сравнению с контролем. Через 24 ч. после воздействия стрессового фактора наиболее сильное угнетение наблюдается на вариантах NaCl 100 мМ + рН 3 и NaCl 100 мМ + рН 10 (угнетение прироста на 64 и 61% соответственно). Через 48 ч. прирост растений на варианте NaCl 100 мМ + рН 10 тормозился на 32% относительно контроля, а на варианте NaCl 100 мМ + рН 3 – на 39%.
После комбинированного воздействия засоления корневой среды NaCl 200 мМ и разных уровней рН существенные различия отмечаются между всеми вариантами опыта и контролем. Через 24 ч. наибольшее угнетение происходило на вариантах
NaCl + рН 3 и NaCl + рН 10 (прирост надземной части меньше на 81 и 77% по сравнению с контролем соответственно). Через 48 ч. угнетение наблюдалось на всех вариантах относительно контроля: при отдельном воздействии засоления – на 45, при рН 10 – на 35, при рН 3 – на 55%.
Аналогичные результаты получены и при изучении других ростовых показателей (площади листовой пластинки и массы надземной части растений овса посевного), где через 24 ч. произошло торможение роста на всех опытных вариантах, а через 48 ч. существенные различия с контролем отмечались у вариантов с кислой корневой средой (как отдельного фактора, так и в сочетании с засолением 100 мМ). При комбинации факторов засоления NaCl 200 мМ и разных рН корневой среды значимые различия высоты растений относительно контроля наблюдались в течение 2 сут. По мнению В.И. Полонского и Н.А. Сурина [2003], чувствительность зерновых культур к кислым почвам наиболее высока в фазе проростков, что согласуется с нашими данными. Резкий сдвиг рН в ту или иную сторону оказывает вредное, а иногда и губительное действие на растение, при этом менее вреден сдвиг рН почвы в щелочную сторону. Н.А. Гаевский, Т.И. Голованова, В.М. Гольд [2012] объясняют это тем, что клетки корня растения выделяют СО 2 , а иногда и органические кислоты, которые нейтрализуют избыточную щелочность.
Заключение
Через 30 мин. после отдельного и комбинированного воздействия засоления NaCl и рН-уровня корневой среды было отмечено усиление окислительно-восстановительных реакций в листьях овса посевного. На всех опытных вариантах повысилось содержание пероксида водорода в сравнении с контролем, наибольшее его количество было одновременно зафиксировано при действии отдельных факторов: кислой (рН 3) и щелочной реакции (рН 10), а также засоления NaCl 100 мМ. Активность каталазы и пероксидаз усилились при отдельном действии всех исследуемых факторов корневой среды. На фоне комбинированного стресса в листьях растений не выявлено существенного увеличения активности этих антиоксидантных ферментов. Возможно, разбалансировка между АФК-генерирующими системами и системами антиоксидантной защиты при сочетании неблагоприятных условий корневой среды связана с недостаточным количеством времени для формирования адаптивного ответа. Не исключено и угнетение защитных механизмов растений, т.к. прирост за 24-48 ч. на этих вариантах минимален. Наибольшее негативное влияние на рост овса оказывало сочетание засоления NaCl и кислой реакции (рН 3) корневой среды.
Список литературы Изменение активности каталазы и пероксидаз в листьях овса посевного под влиянием отдельного и комбинированного воздействия засоления и рН-уровня корневой среды
- Алехина Н.Д. и др. Физиология растений / под ред. И.П. Ермакова. М.: Академия, 2005. 640 с.
- Воскресенская О.Л., Алябышева Е.А., Половникова М.Г. Большой практикум по биоэкологии: учеб. пособие. Йошкар-Ола: Изд-во Мар. ун-та, 2006. Ч. 1. 107 с.
- Гаевский Н.А., Голованова Т.И., Гольд В.М. Избранные главы экологической физиологии растений: конспект лекций. Красноярск, 2012. 91 с.
- Гарифзянов А.Р., Жуков Н.Н. АФК-индуцированные процессы в клетках xTriticosecale в условиях натрий-хлоридного засоления // Известия ТулГУ. Естеств. науки. 2013. № 1. С. 241-250.
- Гарифзянов А.Р., Жуков Н.Н., Иванищев В.В. Образование и физиологические реакции активных форм кислорода в клетках растений // Современные проблемы науки и образования. 2011. № 2. С. 161-183.
- Еремченко О.З., Митракова Н.В., Шестаков И.Е. Природно-техногенная организация почвенного покрова территории воздействия солеотвалов и шламохранилищ в Соликамско-Березниковском экономическом районе // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 311-320.
- Ермаков А.И. и др. Методы биохимического исследования растений. Л.: Агропромиздат, 1987. С.41-43.
- Колупаев Ю.Е. Активные формы кислорода в растениях при действии стрессоров: образование и возможные функции // Вестник Харьковского нац. аграр. ун-та. Сер. Биология. 2007. С. 6-26.
- Полесская О.Г., Каширина Е.К., Алехина Н.Д. Влияние солевого стресса на антиоксидантную систему растений в зависимости от условий азотного питания // Физиология растений. 2006. № 2. С. 207-214.
- Полесская О.Г. Растительная клетка и активные формы кислорода / под ред. И.П. Ермакова. М., 2007. 140 с.
- Полонский В.И., Сурин Н.А. Оценка зерновых злаков на устойчивость к неблагоприятным экологическим факторам. Новосибирск: Изд-во СО РАСХН, 2003. 125 с.
- Радюкина Н.Л. Функционирование компонентов антиоксидантной системы дикорастущих видов растений при кратковременном действии стрессоров: автореф. дис.. д-ра биол. наук. М., 2015. 48 с.
- Рогожин В.В. Пероксидаза как компонент антиоксидантной системы живых организмов. СПб: ГИОРД, 2004. 240 с.
- Abogadallah G.M. Antioxidative defense under salt stress // Plant Signal Behav. 2010. Vol. 5. P. 369-374.
- Apel K., Hirt H. Reactive oxygen species: metabolism, oxidative stress and signal transduction // Annu. Rev. Plant Biol. 2004. Vol. 55. P. 373-399.
- Blokhina O., Virolainen E., Fagerstedt K.V. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: a Review // Ann. Botan. 2003. Vol. 91. P. 179-194.
- Deilein U. et al. Plant salt-tolerance mechanisms // Trend. In Plant Sci. 2014. Vol. 19. P. 371-377.
- Foyer C.H., Noctor G. Defining robust redox signalling within the context of the plant cell // Plant, Cell and Environment. 2015. Vol. 38. P. 239-239.
- Foyer C.H., Noctor G. Oxidant and antioxidant signaling in plants: a reevaluation of the concept of oxidative stress in a physiological context // Plant Cell Environ. 2005. Vol. 29. P. 1056-1071.
- Gechev T.S. et al. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death // BioEssays/ Wiley Periodicals Inc. 2006. Vol. 28. P. 1091-1101.
- Jones D.P. Radical-free biology of oxidative stress // Am. J. Physiol. Cell physiol. 2008. Vol. 295. P. 849-868.
- Meneguzzo S., Navari-Izzo F., Izzo R. Antioxidative responses of shoots and roots of wheat to increasing NaCl concentrations // J. Plant. Physiol. 1999. № 155. P. 274-280.
- Mittler R. Oxidative stress, antioxidants and stress tolerance // Trends in Plant Science. 2002. № 7. P. 405-410.
- Mittler R. et al. Reactive oxygen gene network of plants // Trends in Plant Science. 2004. Vol. 9. P. 490-498.
- Mittova V. et al. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii // Plant, Cell and Environ. 2003. Vol. 26. P. 845-856.
- Miura K., Tada Ya. Regulation of water, salinity and cold stress responses by salicylic acid // Frontiers in plant science. 2014. Vol. 5. P. 1-12.
- Poljsak B., Milisav I. The Neglected Significance of "Antioxidative Stress" // Oxidative Medicine and Cellular Longevity. 2012.
- DOI: 10.1155/2012/480895
- Sagisaka S. The occurrence of peroxide in a perennial plant, Populus gelrica // Plant Physiol. 1976. Vol. 57. P. 308-309.
- Sies H. Physiological society symposium: impaired endothelial and smooth muscle cell function in oxidative stress: oxidants and antioxidants // Experimental Physiology. 1997. Vol. 82. P. 291-295.
