Изменение активности пероксидазной системы при действии гипертермии и засоления NaCl

Автор: Суворов В.Й., Чудинова Л.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология. Охрана природы
Статья в выпуске: 2, 2004 года.
Бесплатный доступ
На проростках гороха исследовали изменения активности и изоферментных составов пероксидазы при действии закаливающих (35°С) и повреждающих температур (40 и 45 °С) с последующим засолением NaCl. При действии закаливающих температур активность фермента в листьях и корнях уменьшалась, что сопровождалось сокращением числа анионных изопероксидаз. При повреждающих температурах активность пероксидазы значительно возрастала, что связано с увеличением числа анионных и изменением активности имеющихся катионных изоформ.
Короткий адрес: https://sciup.org/147204324
IDR: 147204324 | УДК: 581.1
The alteration of activity of the peroxidazal system affected by hyperthermal treatment and salting with NaCl
We have examined the alterations of the activity and isofermental compositions of peroxidasa on the peas shoots affecting them by the low and high temperatures followed by salting with NaCl. The activity of the ferment in the leaves and roots is reduced at low temperatures, where as its activity increases considerably at higher temperatures which is dealt with the mcreasing number of anion isoforms and the alteration of the activity of cation ones.
Текст научной статьи Изменение активности пероксидазной системы при действии гипертермии и засоления NaCl
Пермский государственный университет, 614990, Пермь, ул. Букирева, 15
На проростках гороха исследовали изменения активности и изоферментных составов пероксидазы при действии закаливающих (35°С) и повреждающих температур (40 и 45 °C) с последующим засолением NaCI. При действии закаливающих температур активность фермента в листьях и корнях уменьшалась, что сопровождалось сокращением числа анионных изопероксидаз. При повреждающих температурах активность пероксидазы значительно возрастала, что связано с увеличением числа анионных и изменением активности имеющихся катионных изоформ.
У растений имеется широкий набор изоформ пероксидазы, причем изопероксидазы представляют собой мономерные белки, которые кодируются разными генами (Hiraga et al., 2001). В то же время некоторые изоформы могут быть продуктами одного гена, возникающими в результате модификации на посттранскрипционном / посттрансляционном уровнях (Лебедева и др., 2003).
Пероксидазная система служит уникальным индикатором стрессового состояния растений, она может дать достаточную информацию о их физиологическом состоянии и быть критерием их устойчивости к различным факторам среды (Андреева, 1988; Савич, 1989; Quiroda et al, 2000).
В ответ на действие определенного стрессора устойчивость клеток повышается не только по отношению к нему, но и по отношению к повреждающим агентам другой природы. Данные о существование общих систем устойчивости к избыточному засолению и высокой температуре были получены у про- и эукариотических организмов. Было показано, что на осмотический шок клетки Е. coli отвечали повышением термотолерантности, а на действие повышенных температур реагировали повышением солеустойчивости (Шерман, 1987).
Цель работы заключалась в выяснении влияния комбинированного действия гипертермии и засоления на активность и перестройки изоферментных составов пероксидазы, выступающей в качестве адаптационного фактора повышения устойчивости растений.
Объекты и методы исследований
Исследования проводились на проростках гороха (Pisum sativum L.) сорта Сахарный.
Растения выращивали в водной культуре на смеси Прянишникова (0,5 нормы). Проростки подвергали тепловому воздействию и засолению по следующей схеме: вариант 1 - контроль (смесь Прянишникова), варианты 2, 3, 4 - тепловое воздействие температурами 35, 40, 45°С в течение Зч с последующим засолением 0,4% раствором NaCI.
Для анализа растения брали на 3-й и 10-й день после обработки в начале светлого периода суток (от 9 до 10 ч).
Изоферментный состав пероксидазы анализировали в цитоплазматической фракции листьев и корней гороха с ее предварительной очисткой путем фильтрации через сефадекс G-50 (грубый).
Разделение белков осуществлялось методом диск-электрофореза в 7,5% полиакриламидном геле в щелочной (pH 8,9) и кислой (pH 4,3) буферных системах (Маурер, 1971; Сафоновы, 1971). Локализацию изоферментов устанавливали по относительной электрофоретической подвижности (ОЭП) с использованием в качестве индикаторов для анионных форм красителя Sunset yellow и для катионных - метиленового зеленого.
Проявление изопероксидаз проводили с использованием в качестве субстрата бензидина по прописи Сафоновых (1971), общей пероксидазной активности с тем же субстратом по методу А.Н. Бояркина (1951).
Результаты и их обсуждение
Анализ ростовых параметров проростков гороха показал, что при действии гипертермии (35°С) и последующего засоления снижался ростингиби-рующий эффект по сравнению с засолением NaCI.
При действии повышенных температур (40, 45°С) наблюдалось снижение ростовых параметров корневой системы, хотя размеры вегетативной массы при действии 40°С не уменьшались. На основании изменений ростовых параметров температуру 35°С условно можно принять как закаливающую, а 40 и 45°С - как повреждающие.
Анализ активности пероксидазы показал, что при действии закаливающей и повреждающих температур и последующего влияния хлоридного засоления можно наблюдать различные ответные реакции (таблица).
Изменение активности пероксидазы при гипертермии и засолении (в условных единицах)
|
Орган растения |
Вариант |
Время после обработки |
|
|
3 дня |
10 дней |
||
|
1 |
31,6 ±2,2 |
38,6 ±2,7 |
|
|
Листья |
2 3 |
26,0 ± 1,7 48,2 ± 1,6 |
34,3 ±3,1 56,3 ± 4,0 |
|
4 |
35,9 ± 6,2 |
45,4 ± 1,4 |
|
|
1 |
118,0 ±3,8 |
129,4 ± 5,9 |
|
|
Корни |
2 |
88,5 ± 3,2 |
117,2 ± 3,5 |
|
3 |
148,1 ± 12,1 |
165,8 ±4,7 |
|
|
4 |
192,3 ±5,0 |
245,6 ± 5,3 |
|
Примечание. Варианты: 1 - контроль, 2, 3, 4 - гипертермия (35, 40, 45°С) 3 ч и засоление.
При действии гипертермии 35 °C и засоления наблюдалось значительное снижение активности фермента в листьях и корнях проростков на 3-й день экспозиции и уменьшение ингибирующего эффекта у более возрастных растений. Результаты позволяют допустить, что изменение активности пероксидазы в сторону снижения в первые дни при действии закаливающей температуры отражают адаптивную перестройку обмена веществ и зависят, по-видимому, от синтеза белков de novo и, прежде всего, белков теплового шока. При образовании стресс-белков наблюдается уменьшение синтеза других полипептидов, которые образуются в норме, а в данном случае можно говорить о снижении индуцированного синтеза пероксидазных белков.
При переходе в зону повреждающих температур (40-50°С) активность пероксидазы значительно возрастала, что связано, по-видимому, с индуцированным синтезом этого защитного фермента.
Анализ ' перестроек изоферментных составов согласуется с результатами, представленными в таблице. У контрольных растений, растущих на смеси Прянишникова, на 10-й день в корнях изоформы были представлены 7 компонентами, в листьях - 9. Полиморфизм катионной группы изоферментов был выражен слабее, в корнях обнаруживалось 5 форм, в листьях - 7 (рисунок). При действии закаливающих температур и засоления перестройки наборов изоферментов были связаны с упрощением составов и интенсивностью проявления отдельных форм обеих фракций изоферментов. Гипертермия (40°С) и засоление, наоборот, стимулировали увеличение числа анионных компонентов, в спектре которых можно выделить вновь появляющиеся изоформы и перераспределе-
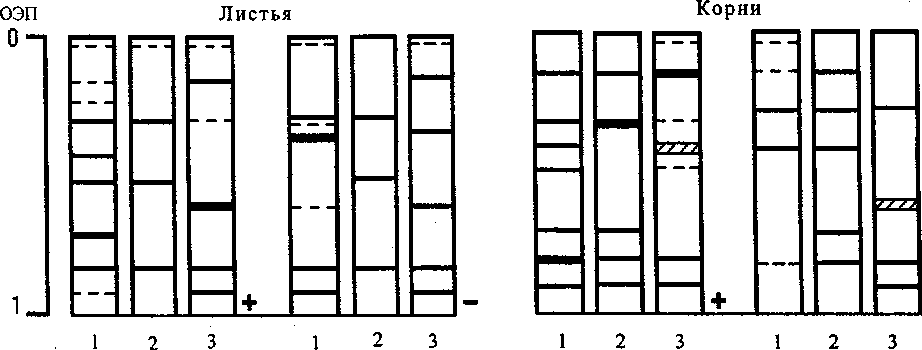
Изменение составов изопероксидаз листьев и корней проростков гороха на 10-й день после последовательного действия гипертермии и засоления NaCl:
1 - контроль; 2 - гипертермия 35°С и засоление; 3 - гипертермия 40°С и засоление ние активности у существующих изоферментов. Ответная реакция катионных изоформ была связана с изменением активности, которая выражалась в различной интенсивности проявления зон в гелях.
Таким образом, из полученных данных можно заключить, что реакция растений на засоление при предварительной обработке их закаливающими и повреждающими температурами имеет качественно различный характер. Следовательно, механизмы обеспечения их устойчивости, которые выражаются в изменении активности пероксидазы, неодинаковы. Первый может быть связан с синтезом стресс-белков или с фосфорилированием полипептидов протеинкиназами (Кузнецов, Рощупкин, 1994), а второй - с индуцированным синтезом ферментов, выполняющих защитные функции.
Список литературы Изменение активности пероксидазной системы при действии гипертермии и засоления NaCl
- Андреева В.А. Фермент пероксидаза: участие в защитном механизме. М.: Наука, 1988. 127 с.
- Бояркин А.Н. Быстрый метод определения активности пероксидазы//Биохимия. Т. 16, № 4.
- Кузнецов Вл.В., Рощупкин Б. В. Стрессорный ответ клеток Nicotiana sylvestris L. на засоление и высокую температуру. 2. Синтез белков теплового шока и фосфорилирование полипептидов//Физиол. раст. 1994. Т. 41, № 4. С. 566-571.
- Лебедева О.В., Ежова Т.А., Мусин СМ., Радюкина Н.Л., Шестаков СВ. Ген PXD контролирует образование трех изоформ анионных перокси-дца Arabidopsis thaliana II Известия АН. Сер. биол. 2003. № 2. С. 159-168.
- МаурерГ. Диск-электрофорез. М.: Мир, 1971.274 с.
- Савич И.М. Пероксидазы -стрессовые белки растений//Успехи совр. биол. 1989. Т. 107, № 3. С. 406-417.
- Сафонов В.И., Сафонова М.П. Исследование белков и ферментов растений методом электрофореза в полиакриламидном геле//Биохимические методы в физиологии растений. М., 1971. С. 113-136.
- Титов А.Ф. Молекулярно-генетический подход к проблеме терморезистентности растений. М.: Наука, 1973. 128 с.
- Шерман М.Ю. Участие белков теплового шока в осморегулящш Е. соН II Молек. биол. 1987. Т. 21,№ I.e. 189-194.
- Hiraga S., Sasa К. По К, Ohashi Y., Matsui Н. A large family of class 111 plant peroxidases II Plant cell Physiol. 2001. Vol. 42. P. 462-468.
- Quiroga M., Guerrero C, Botella M.A., Barcelo A., Amaja I., Medina M.I., Alonso F.J., de Forchetti S.M., Tigier H., Valpuesta V. A tomato peroxidase involved in the synthesis of lignin and suberm II Plant physiol. 2000. Vol. 122, № 4. P. 1119-1127.
