Изменение численности микроорганизмов в зависимости от величины агрегатов гумусового горизонта миграционно-мицелярного чернозема

Автор: Василенко Е.С., Кутовая О.В., Тхакахова А.К., Мартынов А.С.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Статья в выпуске: 73, 2014 года.
Бесплатный доступ
Показано, что количество микроорганизмов и интенсивность почвенно-биологических процессов в агрегатах миграционно-мицелярного постагрогенного чернозема связано с их размерами. Трансформация органических веществ, особенно разложение сложных органических соединений микроорганизмами активнее происходит в мелких агрегатах
Микробный пул, биологическая активность, почвенные агрегаты, почвенно-биологические процессы
Короткий адрес: https://sciup.org/14313580
IDR: 14313580 | УДК: 631.635
Changes in the intensity of soil-biological processes caused by different-sized aggregates of migrationary-mycelial chernozems
The intensity of soil-biological processes reveals changes in different-sized aggregates of post-agrogenic migrational-mycellary chernozems. Transformation of the organic matter and its decomposition by microorganisms in particular becomes more active in fine aggregates (
Текст научной статьи Изменение численности микроорганизмов в зависимости от величины агрегатов гумусового горизонта миграционно-мицелярного чернозема
Почвенные агрегаты являются уникальным продуктом почвообразования, в них протекают практически все микропроцессы, характерные для горизонта или почвы в целом (Хан и др., 2007). Агрегаты являются стабильными функционирующими образованиями, которые рассматриваются как самоорганизующиеся и саморегулирующиеся структурные единицы почвы (Трофимов, 1997). Агрегаты делят почву на многочисленные микрозоны, в которых могут создаваться резко отличающиеся условия (Звягинцев и др., 2005). Каждый почвенный агрегат является комплексом одновременно существующих микросред для разви- тия и жизнедеятельности почвенных микроорганизмов. Место расположения микроорганизмов в разных частях агрегата во многом определяет тип их метаболизма (Hattori, 1988).
Целью работы было определить биологическую активность агрегатов разной величины в миграционно-мицелярном поста-грогенном черноземе Каменной Степи, а также интенсивность и направленность почвенно-биологических процессов.
ОБЪЕКТЫ И МЕТОДЫ
Образец гумусового горизонта чернозема миграционно-мицелярного квазиглееватого глинистого (чернозема типичного) ненарушенного сложения размером 25×25×35 см был взят на территории НИИСХ ЦЧП им. В.В. Докучаева (Каменная Степь) на участке косимой степи рядом с лесополосой № 40, разр. Т-0066 (Хитров и др., 2007). Высушивание почвы при микробиологическом посеве на элективные среды дает искаженные, как правило, заниженные результаты численности микроорганизмов. Количество бактерий уменьшается в 5–10 раз, меняется качественный состав микроорганизмов (Звягинцев и др., 2005). Поэтому рассев на агрегаты производили из свежих образцов естественной влажности. Почва была рассеяна на ситах: 0,25 мм; 0,5 мм; 1 мм; 2 мм; 3 мм; 5 мм; 7 мм, предварительно стерилизованных 70%-ным этиловым спиртом.
В аналитическом центре Почвенного института им. В.В. Докучаева в каждой размерной фракции были определены следующие агрохимические показатели: рН водной и солевой почвенных суспензий (Воробьева, 1998); содержание обменных кальция и магния по Шолленбергу, концентрация Ca2+ и Mg2+ на атомно-абсорбционном спектрометре AAS-3 (Руководство по лабораторным …, 1990); содержание обменного калия по Масловой, концентрация калия на пламенном фотометре Flafo-4 (Воробьева, 1998); подвижные соединения фосфора по Чирикову спектрофотометрическим методом на спектрофотометре Specol-220 (Агрохимические методы …, 1975).
О биологической активности почв судили на основе определения численности эколого-трофических групп микроорганизмов, которое проводили методом посева на твердые питательные среды (мясопептонный агар – МПА , крахмалоаммиачный агар –
КАА , голодный агар – ГА , среда Чапека для культивирования микромицетов) (Теппер и др., 2005) и методом предельных разведений (Практикум по микробиологии …, 1976) на жидкие среды (среда Виноградского для культивирования анаэробных азот-фиксаторов, среда ГНД (Ильина, Фомина, 1983) – для микроорганизмов группы нитратного дыхания). Активность азотобактера оценивали по обрастанию почвенных комочков на среде Эшби (Теппер и др., 2005).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Количественное распределение агрегатов по размерам и их агрохимические показатели представлены в таблице. По ряду показателей не обнаружена зависимость между химическим составом и размером агрегатов: рН водной и солевой вытяжек, содержание Са и Mg во всех структурных отдельностях оставались примерно равными. Однако выявлена достоверная разница по содержанию водорастворимых форм фосфора и калия. Зафиксировано повышение их содержания от мелких агрегатов к крупным. От 5,72 до 7,72 мг/100 г почвы фосфора и от 19,86 до 24,64 мг/100 г почвы калия.
Некоторые агрохимические показатели агрегатов разных размеров гумусового горизонта миграционно-мицелярного постагрогенного чернозема Каменной Степи
|
Размер |
Содер- |
рН |
Са2+ |
Mg2+ |
P 2 O 5 |
K 2 O |
|
|
агрегатов, мм |
жание агрегатов, % |
водный |
солевой |
мг-экв/100 г |
мг/100 г |
||
|
< 0,25 |
01,9 |
6,29 |
5,56 |
33,53 |
6,76 |
5,70 |
Не опр. |
|
0,25–0,5 |
03,4 |
6,26 |
5,50 |
33,36 |
6,72 |
6,60 |
19,90 |
|
0,5–1 |
04,0 |
6,25 |
5,48 |
32,75 |
6,67 |
6,50 |
20,20 |
|
1–2 |
10,7 |
6,25 |
5,48 |
33,23 |
6,74 |
6,70 |
21,70 |
|
2–3 |
08,6 |
6,27 |
5,49 |
33,94 |
6,73 |
6,70 |
24,60 |
|
3–5 |
16,0 |
6,16 |
5,51 |
33,02 |
6,68 |
6,70 |
23,90 |
|
5–7 |
14,8 |
6,22 |
5,52 |
33,84 |
6,58 |
6,90 |
24,60 |
|
>7 |
40,6 |
6,23 |
5,51 |
33,96 |
6,67 |
7,72 |
23,83 |
Фосфор – биогенный элемент, имеет большое значение в энергетике клетки, входит в состав аденозинфосфорных кислот (АТФ, АДФ, АМФ), а также нуклеиновых кислот, фосфолипидов и коферментов. Содержание фосфора в микробной биомассе составляет примерно 30 мг Р/г клеток. От присутствия источников фосфора в среде зависит микробная активность (Современная микробиология, 2005).
Калий участвует в обменных процессах при синтезе аминокислот и белков. Контролируемое накопление К+ в клетках микроорганизмов является основным путем регуляции клеточного тургора. В результате поглощения клеткой калия образуется трансмембранный градиент рН (Современная микробиология, 2005).
Таким образом, увеличение содержания этих элементов на 25–35% в почве может стать причиной изменения не только численности и активности микроорганизмов, но и повлиять на структурные изменения в сообществе.
Для оценки направленности почвенно-биологических процессов, происходящих в различных агрегатах, была изучена структура микробного сообщества. Характеристика микробиологической активности агрегатов гумусового горизонта участка косимой степи чернозема типичного представлена на рис. 1–4.
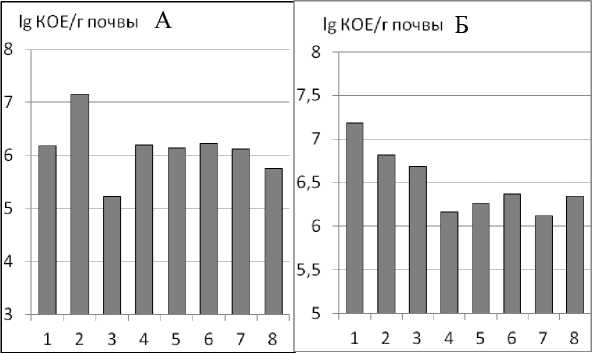
Рис. 1. Численность микроорганизмов-аммонификаторов (А) и общая численность амилолитиков (Б) в различных агрегатах. Здесь и далее: размер агрегатов: 1 – <0,25 мм; 2 – 0,25–0,5 мм; 3 – 0,5–1 мм; 4 – 1–2 мм; 5 – 2–3 мм; 6 – 3–5 мм; 7 – 5–7 мм; 8 – >7 мм.
Микроорганизмы-аммонификаторы (среда МПА) трансформируют органическое вещество, которое поступает в почву в виде растительных остатков и отмерших корней. Природное плодородие черноземов обусловлено аккумулирующей деятельностью естественной растительности и микроорганизмов (Королева и др., 2013), в основном, гетеротрофной группы.
Распределение аммонификаторов по агрегатам ровное (рис. 1А). Исключение составляют агрегаты размером 0,25–0,5 (максимум 7,6 lg КОЕ/г почвы) и 0,5–1,0 (минимум 5,22 lg КОЕ/г почвы). Возможно, это связано с микрозональным распределением экзогенного органического вещества. Процесс разложения белковых веществ происходит достаточно активно независимо от размера агрегатов, однако присутствие экстремальных локусов не исключено. Кроме того, микроорганизмы, разлагая свежее органическое вещество, способствуют образованию водопрочных агрегатов почвы (Хан, 1969).
Наибольшее количество микроорганизмов, использующих минеральные формы азота для построения собственных клеток (среда КАА), отмечено в мелких агрегатах (< 0,25 мм). По мере увеличения размера агрегатов от 0,25 до 2 мм их количество уменьшается, в более крупных агрегатах численность прокариот остается практически на одном уровне (рис. 1Б). Эти микроорганизмы, являясь активными иммобилизаторами легкодоступного углерода, могут участвовать в процессах разложения органических соединений растительного происхождения. Можно предположить, что в мелких агрегатах эти процессы протекают несколько активнее за счет большей их поверхности и, соответственно, за счет улучшенной возможности использования кислорода, необходимого для окисления органических веществ.
Актиномицеты, которые также учитывали на среде КАА, обладают набором разнообразных ферментов, благодаря которым могут разрушать и образовывать сложные органические вещества (Кучаева, 1974). Актиномицеты доминируют на поздних стадиях микробной сукцессии, когда создаются условия для использования труднодоступных субстратов (Зенова, 1992).
Численность актиномицетов максимальна в мелких агрегатах (от < 0,25 до 1 мм), далее, в агрегатах более крупных размеров, уменьшается и наблюдается варьирование их численности, не позволяющее установить зависимость (рис. 2А). Максимальная численность отмечена в агрегатах размером 0,5–1,0 мм (6,12 lg КОЕ/г почвы), минимальная – во фракции 3,0–5,0 мм (4,9 lg КОЕ/г почвы). Вероятно, это связано с тем, что актиномицеты являются прокариотами, требовательными к содержанию кислорода. В условиях хорошей аэрации (мелкие агрегаты) сложные органические вещества почвы минерализуются ими до простых соединений. Внутри более крупных агрегатов свободный кислород частично использован аэробными микроорганизмами, поэтому развитие актиномицетов в крупных агрегатах несколько снижено по сравнению с их численностью в мелких агрегатах размером от 1 мм и меньше. Неравномерное снижение численности актиномице-тов в более крупных агрегатах может быть объяснено микрозональностью почвы – качеством, которое характерно для почвы в целом.
Олиготрофы – микроорганизмы, не требовательные к элементам питания, учитывались на среде ГА. Наблюдается четкая зависимость снижения численности олиготрофов при увеличении размера агрегатов (рис. 2Б). Так как наличие олиготрофов являются показателем заключительного этапа разложения органических веществ, и их численность увеличивается в тех зонах, где минерализация растительных остатков практически завершена, и органи-
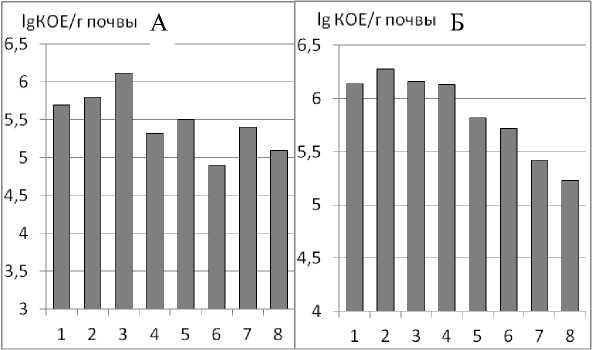
Рис. 2. Численность актиномицетов (А) и олиготрофных микроорганизмов (Б) в различных агрегатах.
ческие вещества находятся в низких концентрациях (Заварзин, Колотилова, 2001), можно предположить что, в мелких агрегатах ряд сукцессионных процессов завершен в бóльшей степени, чем в агрегатах крупных размеров.
Микроскопические грибы – основные деструкторы растительных остатков, учитывались на среде Чапека. Наибольшее количество грибных зачатков (рис. 3А) содержится в самых мелких агрегатах <0,25 мм, затем их численность постепенно уменьшается, в агрегатах средних размеров стабилизируется на уровне 3,5– 4,2 lg КОЕ/г почвы и, затем, снова резко сокращается на два порядка в крупных агрегатах (5,0–> 7 мм) до 2,25 lg КОЕ/г почвы. Микромицеты чрезвычайно чувствительны к аэрации, анаэробных форм среди них практически не встречается, поэтому развитие микроскопических грибов в крупных агрегатах снижено до минимума по причине уменьшения площади поверхности агрегата и создания внутри него анаэробных зон. Вероятно, на рост микро-мицетов оказывает влияние не только фактор аэрации, но также и более свободный доступ к экзогенным органическим веществам в мелких агрегатах.
Увеличенное содержание гумусовых веществ в наиболее мелких фракциях разных типов почв отмечено многими исследователями: в тонкодисперсных фракциях микроагрегатов дерново-
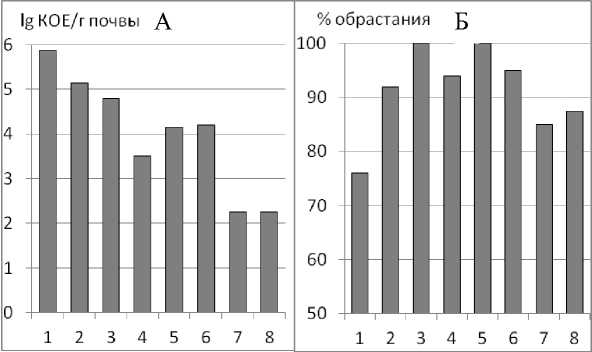
Рис. 3. Численность микромицетов (А) и активность бактерий рода Azotobacter (Б) в различных агрегатах.
подзолистой почвы (Куваева, Фрид, 2001), в илистой фракции черноземов (Когут с соавт., 1998; Милановский, 2009). Отмечено, что в микроагрегатах образуются зрелые гуматы гидрофобной природы, непосредственно участвующие в формировании водопрочных агрегатов (Куваева, 2012). Повышенное содержание гумуса может быть связано с деятельностью почвенных микроскопических грибов, численность которых в наших исследованиях, зарегистрированная в микроагрегатах < 0,25 мм, на 4 порядка больше, чем в агрегатах крупнее 5 мм. Накопление гумусовых веществ идет за счет темноокрашенного мицелия микромицетов, содержащего меланиновые пигменты, которые, как известно, имеют прогумусовый характер и могут участвовать в процессе гумусообразования (Beare et al., 1995).
Процессы, связанные с превращениями азота являются важными как для развития микрофлоры почвы, так и для растений, произрастающих на ней. Были рассмотрены процессы накопления и потери азота из почвы. Наличие аэробной фиксации азота оценивали по встречаемости в почве бактерий рода Azotobacter . Критерием их численности являлась доля обрастания почвенных частиц на среде Эшби.
Из мелких агрегатов (< 0,25 мм) азотобактер выделялся в меньшем количестве по сравнению с более крупными (рис. 6). Это может быть связано с содержанием питательных элементов в этих агрегатах, особенно фосфора, количество которого в них минимально (5,72 мг Р 2 О 5 /100 г). Численность, распространение и активность свободноживущего диазотрофа – бактерий рода Azotobacter , зависит от таких почвенных характеристик, как реакция среды, водно-воздушный режим, содержание фосфора и микроэлементов в почве.
Однако в крупных агрегатах (>5 мм) активность диазотро-фов снижается снова. Лимитирующим фактором в этом случае является, во-первых, возникновение анаэробных зон внутри почвенных агрегатов при потреблении кислорода микроорганизмами на поверхности агрегатов, превышающем скорость его диффузии (Smith, 1980); во-вторых, наличие в почве легкодоступных минеральных форм азота, азотфиксирующая способность аэробного диазотрофа в этом случае не проявляется.
Таким образом, максимальная активность Azotobacter зафиксирована в агрегатах средних размеров от 0,25 до 5 мм. Вероятно, именно за счет этих микроорганизмов формируются водопрочные свойства агрегатов черноземов (Белюченко, Славгород-ская, 2013; Зборищук с соавт., 2007; Чевердин, 2009), поскольку азотобактер, а также сопутствующие ему дрожжи из рода Lipomy-ces (выделяются также на среде Эшби) продуцируют в большом количестве полисахаридную слизь, которая является склеивающим агентом в почве и значительно повышает водопрочность почвенных агрегатов (Кононенко, 1959).
Активность анаэробных азотфиксаторов (рис. 4А) максимальна (5,13 lg КОЕ/г почвы) в агрегатах 0,5–2,0 мм. В почвенных агрегатах других размеров она снижена до 2,35 lg КОЕ/г почвы.
Денитрификация – последнее звено биохимического цикла азота, в котором связанный микроорганизмами азот вновь превращается в атмосферный резерв в виде N 2 .
Активность денитрификаторов оценивали на среде ГНД, используя интегральный показатель – эмиссию газа и помутнение среды (рис. 4Б). Количество микроорганизмов, восстанавливающих нитраты, увеличивается от мелких агрегатов к средним и становится максимальным в агрегатах размером 1,0–2,0 и 3,0–5,0 мм, затем, в более крупных, снова снижается.
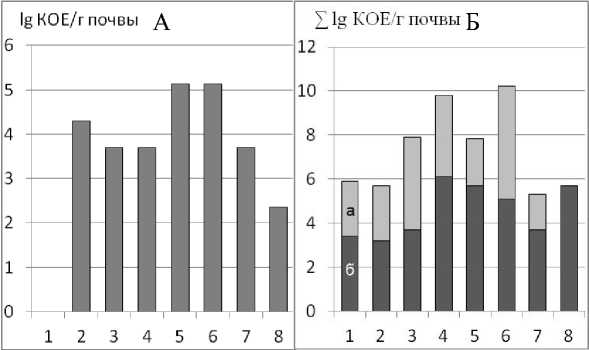
Рис. 4. Активность анаэробных азотфиксаторов (А) и активность денитрифика-торов (Б: а – эмиссия газа, б – изменение цвета) в различных агрегатах.
Денитрификация протекает при разложении органического вещества почвы в условиях недостатка атмосферного кислорода микроорганизмами, восстанавливающих окисные формы неорганических веществ – нитратов. Внутри почвенных агрегатов крупнее 1 мм складываются анаэробные условия, достаточные для протекания денитрификации (Степанов и др., 1997). Снижение процесса денитрификации в более крупных агрегатах, возможно, связано с меньшей доступностью органического вещества микроорганизмам группы нитратного дыхания.
Рассматривая почвенные агрегаты разных размеров как отдельно взятые биологически активные образования, можно сказать, что среди них выделяются агрегаты средних размеров, так называемые, агрономически ценные агрегаты, обеспечивающие плодородие почвы. Условно принято, что размер агрономически ценных агрегатов колеблется в пределах от 0,25 до 10 мм (Вершинин, 1958; Кульман и др., 1961; Edwards, 1967). Однако, за исключением незначительного увеличения объема крупных пор, физические свойства практически не меняются с изменением размера агрегатов крупнее 5 мм. Поэтому возможно принять за агрономически ценные агрегаты размерами от 0,25 до 5 мм (Воронин, 1986).
В агрономически ценных агрегатах (0,25–5 мм) большинство почвенно-биологических процессов происходит более или менее равномерно. Это следует из относительного постоянства численности аммонификаторов, амилолитиков, актиномицетов, микромицетов. Кроме того, в этих агрегатах наблюдается резкое увеличение численности микроорганизмов азотного цикла (азот-фиксация, денитрификация). Поэтому почвенные агрегаты этой размерной группы могут быть названы «биологически ценными».
ЗАКЛЮЧЕНИЕ
Размер агрегатов является одним из факторов, определяющих и численность различных эколого-трофических групп микроорганизмов, и интенсивность почвенно-биологических процессов, происходящих в миграционно-мицелярном постагрогенном черноземе.
Можно допустить, что трансформация органических веществ, особенно, разрушение сложных органических соединений, более интенсивно происходит в мелких агрегатах (< 1 мм), за счет наибольшей численности микроорганизмов углеродного цикла.
Одним из факторов, влияющим на накопление гумусовых веществ в микроагрегатах, отмеченное многими исследователями, может быть деятельность пигментированных микромицетов, численность которых в агрегатах мелких размеров значительно больше. Процессы азотного цикла, значимые для развития растений, наиболее интенсивно протекают в агрегатах средних размеров – агрономически ценных агрегатах (1,0–5,0 мм); за счет наибольшей численности диазотрофов может также повышаться водопрочность этих агрегатов.
Список литературы Изменение численности микроорганизмов в зависимости от величины агрегатов гумусового горизонта миграционно-мицелярного чернозема
- Агрохимические методы исследования почв. М.: Наука, 1975. 656 с.
- Белюченко И.С., Славгородская Д.А. Изменение агрегатного состава чернозема обыкновенного при внесении органо-минерального компоста//Доклады РАСХН. 2013. №4. С. 23-25.
- Вершинин П.В. Почвенная структура и условия ее формирования. М.-Л.: Изд-во АН СССР, 1958. 188 с.
- Воробьева Л.А. Химический анализ почв. М.: Изд-во Моск. ун-та, 1998. 272 с.
- Воронин А.Д. Основы физики почв. М.: Изд-во Моск. ун-та. 1986. 224 с.
- Заварзин Г.А., Колотилова H.H. Введение в природоведческую микробиологию. М.: Книжный дом Университет, 2001. 256 с. С. 192-210.
- Зборищук Ю.Н., Рымарь В.Т., Чевердин Ю.И. Состояние черноземов обыкновенных Каменной Степи. М., 2007. 160 с.
- Звягинцев Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. М.: Изд-во Моск. ун-та, 2005. 445 с.
- Зенова Г.М. Почвенные актиномицеты. М.: Изд-во Моск. ун-та, 1992. 76 с.
- Ильина Т.К., Фомина О.М. Авторское свидетельство № 113328а от 30.06.83. «Питательная среда для культивирования почвенных микроорганизмов».
- Когут Б.М., Травникова Л.С., Титова Н.А., Куваева Ю.В., Ярославцева Н.В. Влияние длительного применения удобрений на соединение органического вещества в легких и илистых фракциях черноземов//Агрохимия. 1998. № 5. С. 13-20.
- Кононенко Е.В. Роль дрожжей из рода Lipomyces в почвенных процессах//Почвоведение. 1959. № 6.
- Королева И.Е., Лебедева И.И., Гребенников А.М. Гумусное и азотное состояние целинных пахотных черноземов//Бюл. Почв. ин-та им. В.В. Докучаева. 2013. Вып. 71. С. 27-35.
- Куваева Ю.В. Групповой состав гумуса фракций тонкодисперсной фазы дерново-подзолистой среднесуглинистой почвы в длительном опыте//Бюл. Почв. ин-та им. В.В. Докучаева. 2012. Вып. 70. С. 18-42.
- Куваева Ю.В., Фрид А.С. Динамика органического вещества тонкодисперсных частиц дерново-подзолистых почв в длительных опытах//Почвоведение. 2002. № 1. С. 52-61.
- Кульман А., Климес-Чмик А. Исследования динамики водопрочности почвенных агрегатов//Почвоведение. 1961. № 3. С. 23-35.
- Кучаева А.Г. Лучистые грибки. Порядки актиномицеты (Actinomycetales) и актинопланы (Actinoplanales)//Жизнь растений. М.: Просвещение,1974. Т. 1. С. 273-288.
- Милановский Е.Ю. Гумусовые вещества почв как природные гидрофобно-гидрофильные соединения. М.: ГЕОС, 2009. 186 с.
- Практикум по микробиологии/Под ред. Н.С. Егорова. М.: Изд-во Моск. ун-та, 1976. 307 с.
- Руководство по лабораторным методам исследования ионно-солевого состава нейтральных и щелочных минеральных почв/Под ред. Н.Б. Хитрова и А.А. Понизовского. М.: ВАСХНИЛ, Почвенный институт им. В.В. Докучаева, 1990. 236 с.
- Современная микробиология. Прокариоты/Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля (в 2-х томах). М.: Мир, 2005. Т. 1. 656 с. Т. 2. 496 с.
- Степанов А.Л., Манучарова Н.А., Полянская Л.М. Продуцирование закиси азота бактериями в почвенных агрегатах//Почвоведение. 1997. № 8. С. 973-976.
- Теппер Е.З., Шильникова В.К., Переверзева Г.И. Практикум по микробиологии. М.: Дрофа, 2005. 256 с.
- Трофимов С.Я. О динамике органического вещества в почвах//Почвоведение. 1997. № 9. С. 1081-1086.
- Хан Д.В. Органо-минеральные соединения и структура почвы. М.: Наука, 1969. 142 с.
- Хан К.Ю., Поздняков А.И., Сон Б.К. Строение и устойчивость почвенных агрегатов//Почвоведение. 2007. № 4. С. 450-456.
- Хитров Н.Б., Лебедева И.И., Чевердин Ю.И., Чижикова Н.П., Ямнова И.А. Морфологические свойства почв Каменной Степи//Каменная Степь: проблемы изучения почвенного покрова. М.: Почв. ин-т им. В.В. Докучаева, 2007. С. 36-71.
- Чевердин Ю.И. Физическое состояние пахотных горизонтов черноземов Каменной Степи//Плодородие. 2009. №4. С. 44-46.
- Beare M.H., Coleman D.C., Crossley D.A., Hendrix P.F., Odum E.P. A hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling//Plant and Soil. 1995. V. 170. № 1. Р. 5-22.
- Edwards A.P., Bremncr J.M. Microaggregates in soil//J. Soil Sci. 1967. V. 18. P. 64-73.
- Hattori T. Soil aggregates as microhabitats for microorganisms//Rep. Inst. Agric. Res. Tohoku. Univ. 1988. V. 37. P. 23-26.
- Smith K.A. A model of the extent of anaerobic zones in aggregated soil, and its potential application to estimates of denitrification//J. Soil Sci. 1980. V. 31. P. 263-277.
