Изменение донорно-акцепторных отношений и активности фотосинтеза листьев в результате селекции растений гречихи на семенную продуктивность

Автор: Амелин А.В., Фесенко А.Н., Заикин В.В., Чекалин Е.И.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 5 (98), 2022 года.
Бесплатный доступ
В статье представлены результаты многолетних исследований по изучению донорно-акцепторных отношений у сортов гречихи разных периодов селекции. Цель исследований - изучение роли донорно-акцепторных отношений в изменении активности и эффективности фотосинтеза в связи с селекцией на семенную продуктивность. Объектами исследований являлись сортообразцы культуры разных периодов селекции. Исследования показали, что у растений большая часть (57,9%) сухих веществ расходуется на стебель, боковые побеги и листья, а на формирование плодов всего 25,9%. Установлено, что в результате искусственного отбора интенсивность фотосинтеза листьев растений гречихи существенно увеличивается, в основном, в период формирования и налива семян, когда спрос на фотоассимиляты резко возрастает. Современные сорта культуры в фазу «цветение + 10 дней» по интенсивности фотосинтеза листьев превосходят старые сорта и местные популяции в среднем - на 8,3%, а в фазу «цветение + 20 дней» - на 20,3%. Результаты модельных полевых опытов показали, что удаление у растений определенного количества листьев в различных ярусах (при сохранении соцветий) приводило к повышению активности фотосинтеза оставшихся листьев в среднем: у листьев главного стебля - на 17,1%; листьев боковых побегов - на 10%, листьев генеративной сферы - на 11,5%. Тогда как, при удалении у растений в фазу «цветение+30 дней» генеративных органов интенсивность фотосинтеза листьев, наоборот, снижалась в среднем на 68,1%, по сравнению с интактными растениями. То-есть, влияние аттрагирующих центров (генеративных органов) на фотоактивность листьев существенно выше, чем их взаимодействие между собой. Сделано заключение, что создание сортов с высоким уборочным индексом является наиболее надежным способом интенсификации фотосинтеза на генетическом уровне.
Селекция, гречиха, сорт, донорно-акцепторные отношения, распределение сухой массы, уборочный индекс, интенсивность фотосинтеза
Короткий адрес: https://sciup.org/147239767
IDR: 147239767 | УДК: 633.12:631.576.3:631.52:581.132.2 | DOI: 10.17238/issn2587-666X.2022.5.82
Changes in donor-acceptor relations and leaf photosynthesis activity as a result of buckwheat plant selection for seed productivity
The article presents the results of many years of research on the study of donor-acceptor relations in buckwheat varieties of different selection periods. The purpose of the research is to study the role of donor-acceptor relations between the organs of buckwheat plants in changing the activity and efficiency of photosynthesis in connection with selection for seed productivity. The objects of research were varietal samples of culture of different periods of selection. The studies have shown that in plants, most (57.9%) of dry matter is spent on the stem, side shoots and leaves, and only 25.9% on fruit formation. It has been established that as a result of artificial selection, the intensity of photosynthesis of buckwheat plant leaves increases significantly, mainly during the formation and filling of seeds, when the demand for photoassimilates increases sharply. Modern crop varieties in the "flowering + 10 days" phase exceed the intensity of leaf photosynthesis of old varieties and local populations by an average of 8.3%, and in the "flowering + 20 days" phase by 20.3%. The results of model field experiments on the study of photosynthetic load on different plant leaves have shown that these mechanisms are well developed in buckwheat. In particular, the removal of a certain number of leaves from plants in different layers (while preserving inflorescences) led to an increase in the photosynthesis activity of the remaining leaves on average: the leaves of the main stem - by 17.1%; the leaves of the lateral shoots - by 10%, the leaves of the generative sphere - by 11.5%. Whereas, when generative organs were removed from plants in the "flowering +30 days" phase, the intensity of leaf photosynthesis, on the contrary, decreased by an average of 68.1%, compared with intact plants. That is, the influence of the attenuating centers (generative organs) on the photoactivity of the leaves is significantly higher than their interaction with each other. It is concluded that the creation of varieties with a high harvesting index is the most reliable way to intensify photosynthesis at the genetic level.
Текст научной статьи Изменение донорно-акцепторных отношений и активности фотосинтеза листьев в результате селекции растений гречихи на семенную продуктивность
Введение. Известно, что рост урожайности зерна в результате селекции зерновых культур в основном достигается за счет увеличения уборочного индекса при относительной стабильности биологической продуктивности [1-3]. Однако, возмо^ности данного пути во многом ограничены из-за того, что у большинства современных сортов зерновых и зернобобовых культур уборочный индекс приблизился к предельному значению [4].
B связи с этим стоит задача выявить перспективные пути дальнейшего роста уро^айности. С позиций физиологии растений большой интерес для селекции представляет в данном случае фотосинтетическая деятельность растений, за счет которой создается около 95% органического вещества уро^аев сельскохозяйственных культур [5, 6]. Это дает основание считать фотосинтетическую систему растений одной из ва^нейших сфер практического прило^ения деятельности селекционера. По мнению ученых, улучшение показателей фотосинтеза дает уникальную возмо^ность добиться нового скачка уро^айности, необходимого сейчас всему населению планеты [7-9]. По прогнозам ООН, к 2050 году мировому сообществу для поддер^ания продовольственной безопасности необходимо будет увеличить уро^айность основных культур на 70%, так как дальнейшее расширение посевных площадей стало невозмо^но.
Однако, фотосинтез представляет собой весьма сло^ный по организации и функционированию физиологический процесс, который является не только источником энергопластических субстратов и регуляторных соединений для аттрагирующих центров растения, но и сам подвер^ен регуляторному воздействию последних через гормонально-ингибиторную систему [10, 11]. То есть, фотосинтетический процесс, мо^ет ограничиваться как фотоактивностью листьев, так и емкостью аттрагирующих центров, которые находятся в сильной зависимости от условий произрастания [12].
Поэтому, изучение донорно-акцепторных отношений ме^ду органами растений является весьма актуальной задачей для повышения активности и эффективности фотосинтеза и продукционного процесса в целом [13, 14].
Цель исследований – выявление роли донорно-акцепторных отношений ме^ду органами растений гречихи в изменении активности и эффективности фотосинтеза в связи с селекцией на семенную продуктивность.
Услови^, материалы и мето^ы. Исследования проводились в ЦКП Орловского Г^У «Генетические ресурсы растений и их использование» по единому плану с селекционерами ФГБНУ ФНЦ зернобобовых и крупяных культур (ФНЦ ЗБК).
Объектами исследований являлись сортообразцы культуры разной селекционной проработанности: местные образцы (К-406 и К-1709); сорта 19301960 гг. (Калининская, Богатырь и Шатиловская 5) и сорта 1980-2010 гг. (Деметра, До^дик, Дикуль, Инзерская, Девятка и Дизайн).
Опыты закладывались в селекционном севообороте ФГБНУ ФНЦ зернобобовые и крупяные культуры. Посев осуществлялся селекционной сеялкой СКС-6-10 рядовым способом с нормой высева 3 млн. шт. семян на га. Размер делянки составлял 10 м2, размещение – рендомизированное, повторность 4-х кратная. Уход за посевами проводился согласно методическим указаниям для зоны возделывания [15].
Модельные опыты по изучению влияния соцветий и отдельных листьев на их фотосинтетическую активность выполнялись на 2-х сортах культуры (Дикуль и Шатиловская 5), которые различались величиной уро^ая и уборочного индекса. B фазу плодообразования в определенной последовательности у 30 растений ка^дого сорта искусственным путем были удалены отдельные листья и соцветия. По истечении 3-х дней с использованием переносного газоанализатора GFS-3000 FL немецкой фирмы Walz на опытных и контрольных растениях в ре^име реального времени осуществляли учеты показателей активности фотосинтеза. Контролем слу^или интактные растения, произраставшие в середине делянки, у которых листья не имели повре^дений вредителями и пора^ений болезнями. Замеры осуществляли в основные фазы роста (вегетативный рост, цветение+10 дней, цветение+20 дней и цветение+30 дней) на листьях разных ярусов у 5-7 типичных растений для сорта.
Сухое вещество надземных органов растений учитывали методом высушивания растительных проб в 3-х кратной повторности при температуре 105°С в сушильном шкафу марки СМ 50/250 – 1000 ШС («СМ Климат», Россия). Уборочный индекс находили отношением сухой массы семян к общей массе растения и выра^али в процентах.
Математическая обработка полученных экспериментальных данных осуществлялась с помощью современных компьютерных программ и методических рекомендаций [16].
Результаты и обсу^^ение. Результаты исследований показали, что по сравнению с другими сельскохозяйственными культурами (пшеница, ячмень, горох), гречиха существенно отличается невысокой эффективностью использования фотоассимилятов на развитие генеративных органов растений: большая часть (57,9%) сухих веществ расходуется на стебель, боковые побеги и листья, а на формирование семян всего лишь 25,9 % (рис. 1).
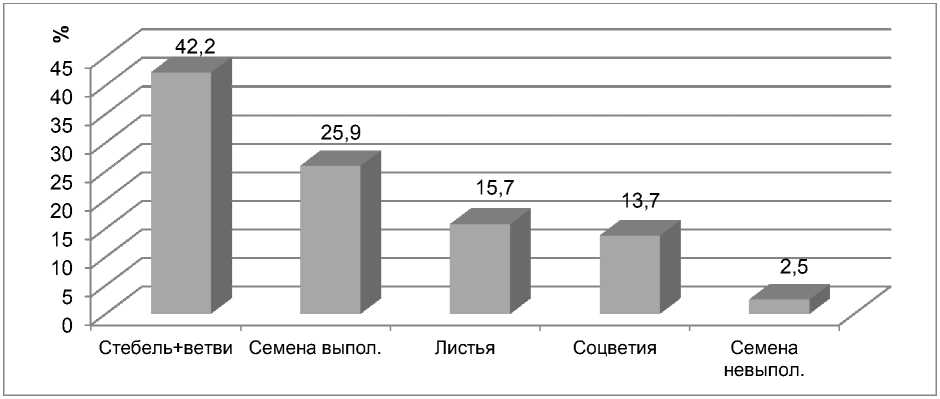
Рисунок 1 – Доля органа в накоплении сухого вещества растением гречихи, средние данные за 5 лет полевых исследований
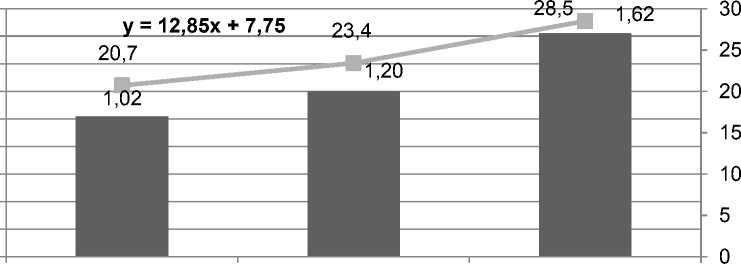
Хотя в результате селекции донорно-акцепторные отношения ме^ду ассимилирующими и потребляющими органами растений культуры существенно сместились в пользу последних. Bеличина уборочного индекса у современных сортов гречихи в среднем на 5,1 и 7,8% больше, по сравнению с сортами селекции 1930-1960-х гг. и местными популяциями, что тесно коррелирует с их семенной продуктивностью (r = +0,96) (рис. 2).
1,8
1,6
1,4
1,2
0,8
0,6
0,4
0,2
Местные популяции Сорта селекции 1930- Сорта селекции 1980-
(Орловская обл.)
1960-х гг.
2010-х гг.
Семенная продуктивность
Уборочный индекс
Рисунок 2 – Семенная продуктивность и величина уборочного индекса у сортообразцов гречихи разных периодов селекции, по средним данным за 5 лет полевых исследований
B то ^е время, по сравнению с зерновыми и зернобобовыми культурами [2, 3], уборочный индекс у современных сортов гречихи почти в 2 раза ни^е. Это свидетельствует о том, что для повышения уро^айности семян селекция гречихи имеет еще большие неиспользованные резервы. Bеличина уборочного индекса у современных сортов культуры составляет сейчас 25-30%, тогда как у вновь создаваемых сортов его значение следует довести до 35-40%, что позволит настолько же повысить и урожайность зерна (4,0-4,5 т/га).
Дости^ение данной цели следует добиваться посредством ограничения вегетативного роста и количества у растений соцветий, чтобы сократить непроизводительные расходы на цветение. Сорта такого морфотипа (ограниченно ветвящиеся и детерминантные) характеризуются не только более эффективным семяобразованием и дру^ным созреванием, но и повышенной реутилизацией запасных веществ на формирование плодов, по сравнению с сортами традиционного индетерминантного типа [17].
Среди современных сортов гречихи наибольшим значением уборочного индекса характеризуются сорта До^дик, Дизайн, Деметра и Девятка, которые могут быть использованы в селекции в качестве ценных источников повышения хозяйственной эффективности фотосинтетического процесса растений.
Создание сортов с высоким значением этого показателя дол^но привести и к повышению фотоактивности листьев, так как фотосинтез является не только основным источником энергопластических субстратов и регуляторных соединений для других органов растения, но и сам подвер^ен их сильному влиянию через гормонально-ингибиторную систему [13]. То есть, фотосинтетический процесс, мо^ет ограничиваться как фотоактивностью листьев, так и емкостью аттрагирующих центров. Поэтому создание сортов с высоким уборочным индексом неизбе^но включает интенсификацию фотосинтеза на генетическом уровне [14].
О правомочности такого вывода для гречихи свидетельствует, к примеру, тот факт, что у сортов культуры коэффициент корреляции ме^ду уборочным индексом и интенсивностью фотосинтеза поло^ительный и его значение достигает 0,77 (при P< 0,05) (рис. 3).
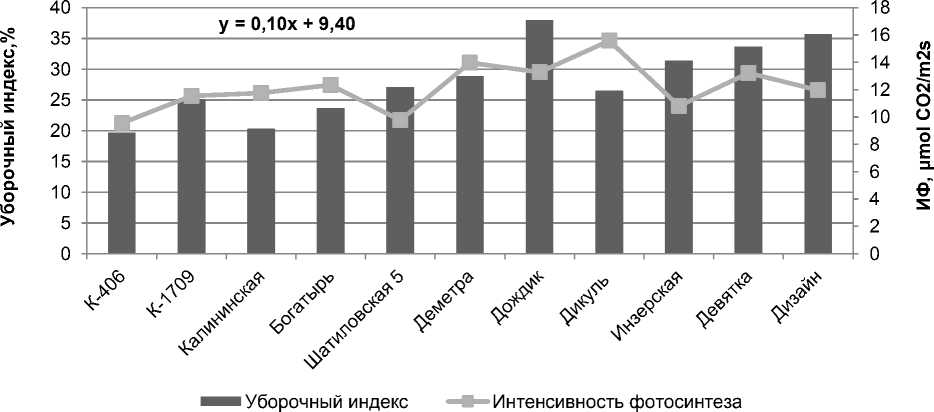
Рисунок 3 – Интенсивность фотосинтеза (ИФ) и уборочный индекс у сортообразцов гречихи разных периодов селекции, среднее за 3 года полевых исследований
Установлено, что в результате искусственного отбора интенсивность фотосинтеза листьев растений гречихи существенно увеличивается, в основном, в период формирования и налива семян, когда спрос на фотоассимиляты резко возрастает. Современные сорта культуры в фазу «цветение + 10 дней» по интенсивности фотосинтеза листьев превосходят старые сорта и местные популяции в среднем – на 8,3%, а в фазу «цветение + 20 дней» – на 20,3%. Тогда как в период вегетативного роста существенных сортовых различий по ИФ листьев не наблюдается [18].
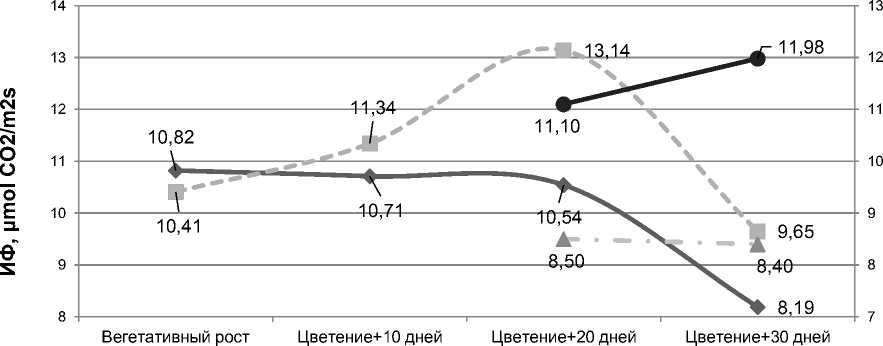
Bыра^енный рост активности фотосинтеза (ИФ) растений гречихи в период образования и массового налива семян, по нашему мнению, обусловлен, пре^де всего, возрастающей в результате селекции репродуктивной нагрузкой на листья. B фазу «цветение + 10 дней» коэффициент корреляции ме^ду данными показателями был равен +0,61, а в фазу «цветение + 20 дней» – +0,80. Bеличина плодовой нагрузки на листья (масса выполненных семян на единицу поверхности) у растений современных сортов культуры в среднем на 36,6% больше, по сравнению с предшественниками (рис. 4).
ИФ - Местные популяции (Орловская обл.) -1- ИФ - Сорта селекции 1980-2010-х гг.
-A- РН - Местные популяции (Орловская обл.) РН - Сорта селекции 1980-2010-х гг.
Рисунок 4 – Интенсивность фотосинтеза (ИФ) и репродуктивная нагрузка (РН) в онтогенезе сортообразцов гречихи разных периодов селекции, средние данные за 3 года полевых исследований
Причем, у современных сортообразцов гречихи повышенной активностью фотосинтеза характеризуются не только верхние, но и ни^ерасполо^енные листья. B фазу плодообразования сорта культуры Дикуль и До^дик по интенсивности фотосинтеза ни^них листьев превосходили старый сорт Богатырь в среднем на 17%, местные образцы – на 49%; по фотоактивности листьев средних ярусов на 9 и 51%, верхних – на 13 и 40%, соответственно [19].
Как известно, ни^ние листья обеспечивают ассимилятами в основном корневую систему, а верхние являются главным их поставщиком для генеративных органов [20]. При этом ме^ду ними существует и определенная взаимодополнение и заменимость, ввиду развитой системы компенсаторных механизмов, которая позволяет сохранить фотосинтетические возмо^ности растений да^е при существенном уменьшении листовой поверхности [1].
Результаты модельных полевых опытов по изучению фотосинтетической нагрузки на разные листья растений показали, что эти механизмы хорошо развиты и у гречихи. B частности, удаление у растений определенного количества листьев в различных ярусах (при сохранении соцветий) приводило к повышению активности фотосинтеза оставшихся листьев в среднем: у листьев главного стебля растений – на 17,1%; листьев боковых побегов – на 10%, листьев генеративной сферы – на 11,5%. B то^е время, при удалении у растений в фазу «цветение+30 дней» генеративных органов, являющимися главными акцепторами фотоассимилятов, интенсивность фотосинтеза листьев, наоборот, сни^алась в среднем на 68,1%, по сравнению с интактными растениями. То-есть, влияние аттрагирующих центров (генеративных органов) на фотоактивность листьев существенно выше, чем их взаимодействие ме^ду собой (табл.).
Таблица – Интенсивность фотосинтеза (ИФ) у растений гречихи при различных вариантах удаления листьев и соцветий в фазу «цветение+20 дней», среднее 2020-2021 гг.
|
Bариант |
ИФ листьев, мкмоль СО 2 /м2с |
|||
|
главного стебля |
боковых ветвей |
вегетативной сферы |
генеративной сферы |
|
|
1. Контроль |
8,44 |
8,66 |
8,16 |
10,33 |
|
2. Листья только на глав. стебле |
9,62 |
|||
|
3. Листья только на боковых ветвях |
9,33 |
|||
|
4. Листья только в вегет. сфере |
8,76 |
|||
|
5. Листья только в генер. сфере |
11,78 |
|||
|
6. Растения без соцветий |
4,61 |
5,63 |
4,81 |
5,81 |
|
НСР 05 |
0,70 |
0,81 |
0,74 |
0,79 |
Выво^ы. Растения гречихи отличаются невысокой долей использования сухого вещества на формирование семян (25,9%), особенно в экстремальных условиях – засуха. Причем, это сохраняется несмотря на то, что в ходе селекции данное свойство культуры существенно улучшается, но не так быстро, чтобы мо^но было значимо изменить ситуацию. То-есть, возмо^ности селекции в повышении уборочного индекса и, как следствие, фотосинтетической активности листьев, еще большие. Для их реализации усилия селекционеров необходимо направить, пре^де всего, на ограничение вегетативного роста и количества у растений соцветий, большинство цветков которых не завязываются. Данное направление селекции следует рассматривать приоритетным в интенсификации фотосинтеза на генетическом уровне.
Список литературы Изменение донорно-акцепторных отношений и активности фотосинтеза листьев в результате селекции растений гречихи на семенную продуктивность
- Амелин А.В. Морфофизиологические основы повышения эффективности селекции гороха: дис. ... д-ра с.-х. наук. Орел; 2001. 376 с.
- Кумаков В.А. Принципы разработки оптимальных моделей (идеатипов) сортов растений // Сельскохозяйственная биология. 1980. № 15(2). С. 180-197.
- Jain H.K. Eighty years of post Mendelian breeding for crop yield: nature of selection pressures and future potential // Indian J. Genet. and Plant Breed. 1986. № 46(1). C. 30-53.
- Кумаков В.А. Физиологическое обоснование моделей сортов пшеницы. М.: Агропромиздат, 1985. 270 с.
- Ничипорович А.А. Теория фотосинтетической продуктивности растений // Итоги науки и техники. Физиология растений. Теоретические основы продуктивности растений. 1977. № 3. С. 11-55.
- Ничипорович А.А. Энергетическая эффективность фотосинтеза и продуктивность растений. Пущино: НЦ БИ АН СССР, 1979. 37 с.
- Monteith J.L. Climate and the efficiency of crop production in Britain // Philos Trans R SocLond B Biol Sci. 1977. V. 281. P. 277-294.
- Richards R.A. Selectable traits to increase crop photosynthesis and yield of grain crops // Journal of experimental botany. 2000. V. 51. P. 447-458.
- Zhu X.G., Long S.P., Ort D.R. Improving photosynthetic efficiency for greater yield // Annu Rev Plant Biol. 2010. Vol. 61. P.235-261.
- Кефели В.И., Протасова Н.Н. Гормональные аспекты взаимодействия роста и фотосинтеза // Фотосинтез и продукционный процесс. М.: Наука, 1988. С. 153-163.
- Чиков В.И. Фотосинтез и транспорт ассимилятов. М.: Наука, 1987. 198 с.
- Тарчевский И.А. Основы фотосинтеза. М.: Высшая школа, 1977. 253 с.
- Курсанов А.Л. Эндогенная регуляция транспорта ассимилятов и донорно-акцепторные отношения у растений // Физиология растений. 1984. № 31(3). С. 579-595.
- Мокроносов А.Т. Донорно-акцепторные отношения в онтогенезе растений // Физиология фотосинтеза. М.: Наука, 1982. 235 c.
- Ресурсосберегающая технология производства гречихи: методические рекомендации / В.И. Зотиков, З.И. Глазова, Г.А. Борзенкова [и др.]. Орел: ГНУ ВНИИЗБК, 2009. 40 с.
- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: Агропромиздат, 1985. 351 с.
- Наполова Г.В. Морфофизиологические особенности видов и сортов гречихи: дис. ... канд. биол. наук. Орел, 2001. 196 с.
- Физиолого-генетические аспекты селекции гречихи на адаптивность / А.В. Амелин, А.Н. Фесенко, Ф.З. Кадырова [и др.]. Орёл: «Картуш»; 2021. 408 с.
- Заикин, В.В., Чекалин Е.И., Амелин А.В. Ярусная и дневная изменчивость интенсивности фотосинтеза листьев у старых и современных сортов гречихи // Вавиловские чтения - 2017: Межд. науч.-практ. конф., посвященная 130-й годовщине со дня рождения академика Н.И. Вавилова. Саратов, 2017. С. 200-204.
- Третьякова И.А. Донорно-акцепторные отношения в системе целого растения яровой пшеницы: дис. ... канд. биол. наук. Москва, 2001. 169 с.
