Изменение экспрессии генов у цыплят-бройлеров (Gallus gallus) кросса Смена 9 разного пола при введении бетаина в рацион
 кросса Смена 9 разного пола при введении бетаина в рацион")
Автор: Йылдырым Е. А., Ильина Л. А., Лаптев Г. Ю., Филиппова В. А., Тюрина Д. Г., Соколова К. А., Заикин В. А., Пономарева Е. С., Фисинин В. И., Егоров И. А., Егорова Т. А., Манукян В. А., Ленкова Т. Н., Дегтярева О. Н., Тишенкова М. С., Демидова Е. С., Кашпоров Л. М., Пащенко В. Е.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Кормовые добавки, кормовые культуры
Статья в выпуске: 6 т.60, 2025 года.
Бесплатный доступ
Оптимизация состава рационов остается актуальной задачей современного птицеводства, учитывая стремление повысить рентабельность производства при минимизации затрат на дорогостоящие компоненты рациона. В настоящей работе впервые получены результаты, демонстрирующие влияние введения в рацион бетаина на экспрессию ключевых генов, связанных с иммунитетом, воспалительными процессами, барьерной функцией кишечника, антиоксидантной активностью и транспортом питательных веществ у цыплят-бройлеров кросса Смена 9 на фоне рационов с пониженным содержанием обменной энергии (на 5 %), лизина (на 10 %) и метионина (на 10 %). Целью работы было изучение влияния различных дозировок (200-400 г/т корма) бетаина на экспрессию ряда генов в тканях слепых отростков кишечника у цыплят-бройлеров кросса Смена 9, а также оценка его влияния на мясную продуктивность птицы, получавшей рацион с пониженным содержанием обменной энергии, лизина и метионина. Исследование проводили на цыплятах-бройлерах (Gallus gallus) кросса Смена 9 на базе ФГБУ СГЦ «Загорское ЭПХ» ВНИТИП (г. Сергиев Посад) в 2025 году. Были сформированы четыре группы птицы (n = 36 в каждой группе с равным разделением по полу): I группа (контрольная) получала основной рацион (ОР) с пониженным содержанием обменной энергии (на 5 %), лизина (на 10 %) и метионина (на 10 %) в соответствии с рекомендациями ВНИТИП (17); II группа - ОР с добавлением 200 г/т комбикорма бетаина в форме кристаллического порошка с содержанием 95 % триметилглицина («Taian Havay Chemicals Co., Ltd», Китай), III группа - ОР с добавлением 300 г/т бетаина, IV группа - ОР с добавлением 400 г/т бетаина. Бетаин вносили посредством тщательного смешивания с комбикормом вручную. Кормление осуществляли вволю с использованием рассыпных комбикормов. Цикл кормления был трехэтапным: первый этап (до 14-х сут жизни) - использование стартерного комбикорма Старт (СГЦ «Загорское ЭПХ», Россия); второй этап (15-21-е сут) - применение ростоформирующего комбикорма Рост (СГЦ «Загорское ЭПХ», Россия; третий этап (22-35-е сут) - завершающая фаза с применением финишного комбикорма Финиш (СГЦ «Загорское ЭПХ», Россия). В конце эксперимента убивали птицу посредством декапитации и проводили отбор тканей слепых отростков кишечника для последующего анализа экспрессии генов. Анализ экспрессии генов был выполнен методом количественной полимеразной цепной реакции в реальном времени. Общую РНК выделяли из образцов тканей с использованием коммерческого набора Aurum™ Total RNA («Bio-Rad», США) в соответствии с протоколом производителя. qRT-PCR выполняли с использованием набора SsoAdvanced™ Universal SYBR® Green Supermix («Bio-Rad», США) на детектирующем амплификаторе DTlite 4S1 (НПК «ДНК-технология», Россия). Использовали праймеры для генов IL6, IL8L2, PTGS2, кодирующих провоспалительные цитокины, AvBD1, AvBD2, AvBD9, AvBD10, AvBD11, кодирующих антимикробные пептиды, Casps6, кодирующего каспазу-6, MUC2 , кодирующего синтез муцина, SGLT2, кодирующего натрий-глюкозный котранспортер 2, SOD1, кодирующего супероксиддисмутазу 1. Относительную экспрессию генов рассчитывали с использованием метода 2-ΔΔCT. В дополнение к изучению экспрессии генов, фиксировали живую массу цыплят индивидуально, среднесуточный прирост живой массы, убойный выход и содержание абдоминального жира в тушах. Представляют интерес выявленные половые и дозозависимые различия в ответе на бетаин. Настоящая работа впервые демонстрирует сложный, дифференцированный эффект бетаина на экспрессию генов, связанных с иммунитетом, воспалением и метаболизмом глюкозы, в зависимости от пола и дозировки. Так, наиболее значительное увеличение живой массы петушков (на 7,2 и 7,3 %, p ≤ 0,05) наблюдалось при повышенных дозировках бетаина (300 и 400 г/т), что демонстрирует ранее известные свойства бетаина по улучшению продуктивности цыплят-бройлеров кросса Смена 9. При этом было выявлено существенное снижение отложения абдоминального жира (p ≤ 0,05). Бетаин вызывал дифференцированную реакцию, характеризующуюся повышением экспрессии SOD1 -ключевого гена антиоксидантной защиты. Влияние бетаина на экспрессию многих генов иммунитета зависело от пола птицы. Так, у курочек произошло более чем 8-кратное увеличение экспрессии гена AvBD1 при дозировке 200 г/т бетаина (p ≤ 0,01). В то же время при более высоких дозах отмечалось снижение экспрессии других генов семейства AvBD (p ≤ 0,05). Отмечено снижение уровня мРНК генов провоспалительных факторов (IL6, IL8 и PTGS2) (p ≤ 0,05). Количество мРНК гена Casp6 повышалось у курочек при скармливании 200 г/т бетаина в 13,9 раза по сравнению с контролем (p ≤ 0,01). У петушков в группе с дозировкой 200 г/т произошел резкий скачок экспрессии SGLT2 (натрий-глюкозный транспортер второго типа) - в 362 раза (p ≤ 0,001), а при увеличении дозировки до 300 и 400 г/т - соответственно в 39,4 и 13,5 раза (p ≤ 0,01).
Бетаин, экспрессия генов, мясная продуктивность, цыплята-бройлеры, иммунитет, воспаление, рацион
Короткий адрес: https://sciup.org/142247723
IDR: 142247723 | УДК: 619:636.52/.58:636.084:577.2 | DOI: 10.15389/agrobiology.2025.6.1097rus
Changes in gene expression in male and female Smena 9 broiler chickens (Gallus gallus) fed dietary betaine
Optimizing poultry diets remains a pressing issue in modern poultry farming, given the desire to increase production profitability while minimizing the cost of expensive diet components. This study provides the first results demonstrating the effect of betaine supplementation on the expression of key genes associated with immunity, inflammation, intestinal barrier function, antioxidant activity, and nutrient transport in Smena 9 broiler chickens fed diets with reduced metabolizable energy (by 5%), lysine (by 10 %), and methionine (by 10 %). The work aimed at studying effects of 200-400 g/t betaine on the expression of a number of genes in the caecum tissues in Smena 9 cross broiler chickens/ In addition, we aimed to assess its effect on the meat productivity of poultry fed a diet with a reduced content of metabolizable energy, lysine and methionine. The study was conducted on broiler chickens ( Gallus gallus ) of the Smena 9 cross (Zagorskoye EPH, the All-Russian Research and Technological Poultry Institute RAS, Sergiev Posad, 2025). Four groups of birds were formed (n = 36 each), equal in males and females. Group I (control) received the basic diet (BD) with a reduced content of metabolizable energy (by 5 %), lysine (by 10 %) and 200 g/t of betaine crystalline powder containing 95 % trimethylglycine (Taian Havay Chemicals Co., Ltd, China). OR in group III was added with 300 g/t of betaine, in group IV with 400 g/t of betaine. Betaine was added by thoroughly mixing it with the compound feed manually. Feeding was ad libitum using crumbled compound feed. The feeding cycle was three-stage, the first stage (up to 14 days of life) with the starter compound feed Start (SGC Zagorskoye EPH, Russia); the second stage (15-21 days) with growth-forming compound feed Rost (SGC Zagorskoe EPH, Russia), and the third stage (22-35 days) with the finishing compound feed Finish (SGC Zagorskoe EPH, Russia). At the end of the experiment, the birds were decapitated and the caeca tissues were collected for gene expression analysis. Gene expression analysis was performed by quantitative real-time polymerase chain reaction (qRT-PCR). Total RNA was isolated from tissue samples using the commercial Aurum™ Total RNA kit (Bio-Rad, USA) according to the manufacturer's protocol. qRT-PCR was performed (a DTlite 4S1 detection amplifier, NPK DNA-technology, Russia) using the SsoAdvanced™ Universal SYBR® Green Supermix kit (Bio-Rad, USA). Primers were used to the genes IL6 , IL8L2 , PTGS2 , encoding proinflammatory cytokines, AvBD1, AvBD2, AvBD9, AvBD10, AvBD11, encoding antimicrobial peptides, Casps6 , encoding caspase-6, MUC2 , encoding mucin synthesis, SGLT2 , encoding sodium-glucose cotransporter 2, SOD1 , encoding superoxide dismutase 1. Relative gene expression was calculated by the 2-ΔΔCT method. The chickens’ bodyweight, average daily live weight gain, slaughter yield and abdominal fat content in carcasses were assessed individually. The most significant increase in the live weight of cockerels (by 7.2 and 7.3 %, p ≤ 0.05) was observed at higher doses betaine (300 and 400 g/t), demonstrating the previously known properties of betaine in improving the productivity of Smena 9 broiler chickens. We also revealed a significant decrease in abdominal fat deposition (p ≤ 0.05). Betaine caused a differentiated response characterized by an increase in the expression of the SOD1 gene, a key gene for antioxidant defense. The effect of betaine on the expression of many immune genes depended on the sex of the bird. Thus, in hens, there was a more than 8-fold increase in the expression of the AvBD1 gene for 200 g/t betaine (p ≤ 0.01). At higher doses, the expression of other genes of the AvBD family decreased (p ≤ 0.05). A decrease in the mRNA level of proinflammatory genes IL6 , IL8 , and PTGS2 (p ≤ 0.05) occurred. The Casp6 mRNA level increased in hens fed 200 g/t betaine by 13.9 times compared to the control (p ≤ 0.01). In cockerels of the group fed 200 g/t betaine, there was a sharp jump in the expression of SGLT2 (sodium-glucose transporter type 2) by 362 times (p ≤ 0.001), with an increase by 39.4 and 13.5 times for 300 and 400 g/t, respectively (p ≤ 0.01). Thus, when metabolizable energy, lysine and methionine in feeds are reduced, the dietary betaine has a positive effect on the production indicators of Smena 9 cross broiler chickens. The results obtained demonstrate complex and sex- and dosage-dependent effects of betaine on the immune system of the birds, as well as its potential role in modulating inflammatory processesand improving glucose metabolism through the regulation of gene expression.
Текст научной статьи Изменение экспрессии генов у цыплят-бройлеров (Gallus gallus) кросса Смена 9 разного пола при введении бетаина в рацион
В современном птицеводстве оптимизация кормления — это основополагающий фактор, который определяет экономическую эффективность отрасли (1). Полноценные протеиновые корма занимают центральное место в обеспечении роста и развития птицы (2). Такое биологически активное вещество, как бетаин (триметилглицин) играет важную роль для поддержания здоровья и продуктивности сельскохозяйственной птицы (3). Бетаин выступает в качестве донора метильных групп в реакциях, катализируемых бетаин-гомоцистеинметилтрансферазой, играя роль в регенерации метионина из гомоцистеина (4). Благодаря этому бетаин снижает потребность птицы в дорогостоящем метионине (5). Также бетаин выступает в качестве осмопротектора, помогая организму справляться с осмотическим стрессом (6). Бетаин способен усиливать аппетит и способствовать лучшей усвояемости питательных веществ из корма, что позитивно сказывается на его конверсии (7). Бетаин также облегчает преобразование и всасывание жирных кислот, увеличивая их поступление в кровь и ткани, что улучшает обмен веществ и эффективность использования корма. Это особенно важно при скармливании низкопитательных или нестандартных рационов, где необходима дополнительная поддержка организма (8).
Известно, что бетаин поддерживает целостность эпителия кишечника, улучшая его барьерную функцию и снижая риск проникновения патогенных микроорганизмов (9). Повышенная плотность микроворсинок в кишечнике способствует более эффективному всасыванию питательных веществ (10).
Одна из важных особенностей бетаина заключается в его участии в липидном обмене. Вещество препятствует чрезмерному накоплению жира, активизируя мобилизацию жирных кислот из депо и ускоряя их использование в обменных процессах. Это приводит к формированию высококачественного мяса с оптимальным содержанием жира (11).
Традиционное кормление птицы строится на соблюдении оптимальных норм обмена энергии, аминокислот и других питательных веществ, важных для максимального раскрытия генетического потенциала. Тем не менее желание снизить затраты вынуждает производителей мяса и яиц искать альтернативные решения, которые позволят использовать менее полноценные по составу корма без ущерба для продуктивности (12). Предполагается, что включение в рацион бетаина компенсирует недостаток питательных веществ и энергии в корме без снижения показателей продуктивности птицы.
Показано, что дополнение рационов цыплят-бройлеров бетаином способствует улучшению переваримости и использования питательных веществ, повышению выхода живой массы и качества мяса (13). Одним из аспектов влияния бетаина на организм птицы, вероятно, служит его способность воздействовать на экспрессию ключевых генов, регулирующих 1098
иммунитет, пищеварение и метаболизм. Так, добавление бетаина в питьевую воду цыплят-бройлеров изменяло экспрессию мРНК аквапоринов ( AQP1-4 и AQP 9 ), связанных с транспортом воды, рецепторов глюкокортикоидов ( GR ), а также проопиомеланокортина ( POMC ), участвующего в регуляции стресса (14). Включение бетаина в рацион цыплят-бройлеров изменило в тканях печени экспрессию генов, связанных с липидным обменом ( PLIN1 , ACACA , DGAT1 ), углеводным обменом ( PKLR , PGAM1 ) и иммунной системой ( IFITM3 , MX1 , ISG15 ) (15). На сегодняшний день влияние бетаина на экспрессию ключевых генов у цыплят-бройлеров кросса Смена 9 не изучалось.
Ранее было установлено, что экспрессия генов иммунитета ( IL8 , IRF7 , PTGS2 , AvBD1 , AvBD2 , AvBD9 , AvBD10 , Casp6 ) и адаптационного потенциала ( CAT1 , HSF1 , HSF2 , SOD , Gpx1 , HO-1 ) различается в зависимости от генотипа (линии СМ5 и СМ9) и пола у кур и петухов родительского поголовья мясного кросса Смена 9. В частности, у петухов линии СМ5 экспрессия генов HSF1 и HSF2 была в среднем в 1,6 и 3,0 раза выше, чем у кур той же линии (р ≤ 0,05), а у петухов линии СМ9 экспрессия генов AvBD2 , AvBD9 , AvBD10 , IL8 и PTGS2 усиливалась по сравнению с петухами линии СМ5 соответственно в 7,6; 5,3; 2,1; 6,3 и 1,5 раза (р ≤ 0,05) (16).
В настоящей работе впервые получены результаты, демонстрирующие влияние введения в рацион бетаина на экспрессию ключевых генов, связанных с иммунитетом, воспалительными процессами, барьерной функцией кишечника, антиоксидантной активностью и транспортом питательных веществ у цыплят-бройлеров кросса Смена 9 на фоне рационов с пониженным содержанием обменной энергии (на 5 %), лизина (на 10 %) и метионина (на 10 %).
Целью работы было изучение влияния различных дозировок бетаина (200-400 г/т корма) на экспрессию ряда генов в тканях слепых отростков кишечника у цыплят-бройлеров кросса Смена 9, а также оценка влияния бетаина на мясную продуктивность птицы, получавшей рацион с пониженным содержанием обменной энергии, лизина и метионина.
Ìåòîäèêà. Исследование проводили на цыплятах-бройлерах ( Gallus gallus ) кросса Смена 9 на базе ФГБУ СГЦ «Загорское ЭПХ» ВНИТИП (г. Сергиев Посад) в 2025 году. Были сформированы четыре группы птицы ( n = 36 в каждой группе с равным разделением по полу): I группа (контрольная) получала основной рацион (ОР) с пониженным содержанием обменной энергии (на 5 %), лизина (на 10 %) и метионина (на 10 %) в соответствии с рекомендациями ВНИТИП (17); II группа — ОР с добавлением 200 г/т комбикорма бетаина в форме кристаллического порошка (содержание 95 % триметилглицина, «Taian Havay Chemicals Co., Ltd», Китай), III группа — ОР с добавлением 300 г/т бетаина, IV группа — ОР с добавлением 400 г/т бетаина. Пониженное содержание обменной энергии, лизина и метионина было смоделировано для имитации условий удешевления рациона, что нередко характерно для практики птицеводства. Бетаин вносили посредством тщательного смешивания с комбикормом вручную.
Для всех групп птицы, содержащихся в клеточных батареях («Big Dutchman AG», Германия), поддерживались стандартные условия (включая плотность посадки, освещение, температуру и влажность) в соответствии с рекомендациями, разработанными для кросса (18). Птицу кормили вволю с использованием рассыпных комбикормов. Цикл кормления был трехэтапным: первый этап (до 14-х сут жизни) — использование стартерного комбикорма Старт (СГЦ «Загорское ЭПХ», Россия); второй этап (15-21-е сут) — применение ростоформирующего комбикорма Рост (СГЦ «Загорское ЭПХ»,
Россия; третий этап (22-35-е сут) — завершающий с применением финишного комбикорма Финиш (СГЦ «Загорское ЭПХ», Россия). В состав комбикормов добавляли премиксы, обеспечивающие необходимое количество витаминов и микроэлементов.
В конце эксперимента птицу убивали посредством декапитации и отбирали ткани слепых отростков кишечника для анализа экспрессии генов. Ткани фиксировали (консервировали) с помощью раствора «RNAlater» («In-vitrogen Corporation», подразделение корпорации «Thermo Fisher Scientific,
Inc.», США) согласно протоколу производителя.
Экспрессию генов анализировали методом количественной полимеразной цепной реакции в реальном времени (qRT-PCR). Общую РНК выделяли из образцов тканей с использованием коммерческого набора Aurum™ Total RNA («Bio-Rad», США) в соответствии с протоколом производителя. РНК подвергали обратной транскрипции в комплементарную ДНК (кДНК) с помощью набора iScript™ Reverse Transcription Supermix («Bio-Rad», США). qRT-PCR выполняли с использованием набора SsoAdvanced™ Universal SYBR® Green Supermix («Bio-Rad», США) на детектирующем амплифика-торе DTlite 4S1 (НПК «ДНК-технология», Россия). Режим и условия амплификации: 5 мин при 95 ° C; 30 с при 95 ° C, 30 с при 60 ° C, 30 с при 70 ° C (40 циклов). Смесь для амплификации содержала 10 мкл SsoAdvanced™ Universal SYBR® Green Supermix, 0,2 мкл прямого и 0,2 мкл обратного праймеров, 1 мкл кДНК и была доведена водой до общего объема.
Использовали праймеры для генов IL6, IL8L2, PTGS2, кодирующих провоспалительные цитокины (играют важную роль в развитии воспалительного ответа), AvBD1, AvBD2, AvBD9, AvBD10, AvBD11, кодирующих антимикробные пептиды (участвуют в защите слизистых оболочек от патогенов), Casps6, кодирующего каспазу-6 (вовлечен в процесс апоптоза клеток), MUC2, кодирующего синтез муцина (основной компонент слизи, защищающий слизистую оболочку кишечника), SGLT2, кодирующего натрий-глю-козный котранспортер 2 (участвует в транспорте глюкозы в кишечнике), SOD1, кодирующего супероксиддисмутазу 1 (антиоксидантный фермент, за- щищающий клетки от окислительного стресса):
Ген Нуклеотидные последовательности праймеров (5´ → 3´)
SOD1 , супероксиддисмутаза 1
F: CGGGCCAGTAAAGGTTACTGGAA
R: TGTTGTCTCCAAATTCATGCACATG
AvBDl , р -дефензин 1
AvBD2 , р -дефензин 2
AvBD9 , птичий бета-дефензин 9
AvBD10 , птичий бета-дефензин 9
AvBD11 , птичий бета-дефензин 11
IL6 , интерлейкин 6
IL8L2 ( IL8 ), интерлейкин 8
PTGS2 , простагландин-эндоперок-сидсинтаза
Casp6 , каспаза 6
SGLT2, натрий-глюкозного котранспортер 2-го типа
MUC2 , муцин 2
-
F: CCGTTTCTGTCACCGTCA
R: CCTTTGCTAAAAATCCCTTC
-
F: GCACTCCAGGTTTCTCCA
R: GGCGTCCGACTTTGATTA
-
F: AACACCGTCAGGCATCTTCACA
R: CGTCTTCTTGGCTGTAAGCTGGA
-
F: GCTCTTCGCTGTTCTCCTCT
R: CCAGAGATGGTGAAGGTG
-
F: AGTCTGCAATTCGTTAGAGGCG
R: GGATGTGGTTTCCAAGGGTTTA
F: AGGACGAGATGTGCAAGAAGTTC
R: TTGGGCAGGTTGAGGTTGTT
-
F: GGAAGAGAGGTGTGCTTGGA
R: TAACATGAGGCACCGATGTG
-
F: TCGAGATCACACTTGATTGACA
R: TTTGTGCCTTGTGGGTCAG
-
F: CAGAGGAGACAAGTGCCAGA,
R: CCAGGAGCCGTTTACAGTTT
-
F: ACCAAGTACTGCAAGGCGAA
R: TGAGGGTTCCTCTTCTGGCT
-
F: CTGGCTCCTTGTGGCTCCTC
R: AGCTGCATGACTGGAGACAACTG
Относительную экспрессию генов рассчитывали с использованием метода 2 - AA CT (19). Уровень экспрессии генов в контрольных группах был принят за 1.
Одновременно с исследованием экспрессии генов фиксировали живую массу цыплят индивидуально, среднесуточный прирост живой массы, убойный выход и содержание абдоминального жира в тушах (20).
При проведении эксперимента руководствовались принципами гуманного обращения с животными согласно международным стандартам защиты лабораторных животных. Все процедуры выполняли в строгом соответствии с нормами Европейского сообщества относительно использования позвоночных животных в научных исследованиях, принятыми в рамках Конвенции Совета Европы ETS ¹ 123 (21).
Математическую и статистическую обработку результатов осуществляли методом многофакторного дисперсионного анализа (ANOVA) в программах Microsoft Excel XP/2003, R-Studio v. 1.1.453 , США). Вычисляли средние значения (M) и стандартные ошибки среднего (±SEM). Для сравнения двух контрольной и опытной групп применяли t-критерий Стьюдента. Средние значения сравнивали с использованием теста достоверно значимой разницы Тьюки (HSD) и функции TukeyHSD в пакете R Stats Package (США).
Результаты. Состав и питательная ценность каждого из этих комбикормов, использованных на разных этапах выращивания птицы, представлены в таблице 1. Содержание премиксов в составе кормов приведено в таблице 2.
-
1. Состав (%) комбикормов, используемых при выращивании цыплят-бройлеров ( Gallus gallus ) кросса Смена 9 (ФГБУ СГЦ «Загорское ЭПХ» ВНИТИП, г. Сергиев Посад, 2025 год)
-
2. Содержание витаминов и микроэлементов в 1 кг комбикормов, используемых при выращивании цыплят-бройлеров ( Gallus gallus ) кросса Смена 9 (ФГБУ СГЦ «Загорское ЭПХ» ВНИТИП, г. Сергиев Посад, 2025 год)
Компонент
Старт (до 14-х сут)
Рост (15-21-е сут)
Финиш (22-35-е сут)
Витамин А, тыс. МЕ/кг
12,00
10,00
10,00
Витамин Д 3 , тыс. МЕ/кг
3,50
3,00
3,00
Витамин Е, мг/кг
30,00
20,00
20,00
Витамин К 3 , мг/кг
2,00
1,00
1,00
Витамин В 1 , мг/кг
2,00
1,00
1,00
Витамин В 2 , мг/кг
8,00
6,00
6,00
Витамин В 6 , мг/кг
3,00
3,00
3,00
Витамин В 12 , мг/кг
0,025
0,025
0,025
Биотин, мг/кг
0,10
0,05
0,05
Холин, мг/кг
500,00
500,00
500,00
Фолиевая кислота, мг/кг
0,50
0,50
0,50
Никотиновая кислота, мг/кг
30,00
20,00
20,00
|
Компонент |
Старт (до 14-х сут) |
Рост (15-21-е сут) |
Финиш (22-35-е сут) |
|
Пшеница |
41,03 |
41,86 |
44,95 |
|
Соевый шрот |
29,50 |
21,28 |
16,75 |
|
Кукуруза |
15,00 |
15,00 |
15,00 |
|
Соя полножирная |
5,00 |
10,00 |
10,00 |
|
Масло подсолнечное |
3,02 |
3,82 |
4,53 |
|
Мука рыбная |
2,50 |
1,50 |
– |
|
Жмых подсолнечный |
– |
3,02 |
4,73 |
|
Монокальцийфосфат |
1,44 |
1,12 |
1,28 |
|
Известняк (Са 36 %) |
1,01 |
1,03 |
1,15 |
|
Премикс (0,5 %) |
0,50 |
0,50 |
0,50 |
|
Соль |
0,27 |
0,29 |
0,34 |
|
Лизин сульфат |
0,25 |
0,22 |
0,35 |
|
DL-гидроксиметионин, содержащий 88 % OH-метионина |
0,30 |
0,24 |
0,24 |
|
Треонин |
0,09 |
0,04 |
0,09 |
|
Холин хлорид (60 %) |
0,08 |
0,08 |
0,08 |
|
Фекорд-Концентрат (ООО «Фермент», Россия) |
0,01 |
– |
0,01 |
|
Всего |
100,0 |
100,0 |
100,0 |
Примечание. Прочерки означают, что указанный компонент отсутствует в составе комбикорма.
Продолжение таблицы 2
|
Пантотеновая кислота, мг/кг |
10,00 |
10,00 |
10,00 |
|
Марганец, мг/кг |
100 |
100 |
100 |
|
Цинк, мг/кг |
70,00 |
70,00 |
70,00 |
|
Железо, мг/кг |
25,00 |
25,00 |
25,00 |
|
Медь, мг/кг |
3,50 |
3,50 |
3,50 |
|
Йод, мг/кг |
0,70 |
0,70 |
0,70 |
|
Селен, мг/кг |
0,300 |
0,300 |
0,300 |
При введении бетаина в различных дозировках в рацион цыплят-бройлеров кросса Смена 9 были отмечены некоторые изменения в зоотехнических показателях (рис. 1). Сохранность поголовья птицы во всех группах составила 100 %.
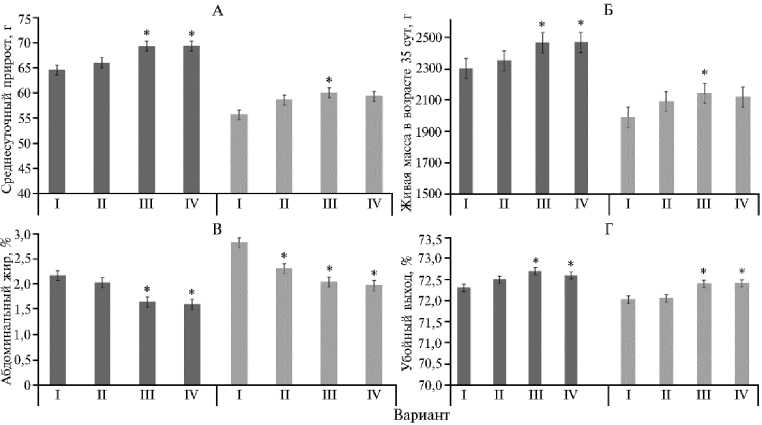
Рис. 1. Среднесуточные прирост живой массы (А) ; живая масса в возрасте 35 сут (Б) ; абдоминальный жир (В) ; убойный выход (Г) у петушков (слева) и курочек (справа) ( Gallus gallus ) кросса Смена 9 в зависимости от включения в рацион добавки бетаина в разных дозах ( n = 3, M ±SEM; ФГБУ СГЦ «Загорское ЭПХ» ВНИТИП, г. Сергиев Посад, 2025 год) . Описание групп см. в разделе «Методика».
* Различия с контролем статистически значимы при р ≤ 0,05.
Так, добавление бетаина в дозировках 300 г/т (III опытная группа) и 400 г/т (IV опытная группа) привело к наибольшему (на 7,2-7,3 %) увеличению живой массы петушков в возрасте 35 сут относительно контроля (2300±28,9 г) — соответственно до 2465±16,33 и 2467±10,07 г (p ≤ 0,05). У курочек наблюдалась аналогичная тенденция: максимальная живая масса в возрасте 35 сут была отмечена в III опытной группе (2141±11,97 г) по сравнению с I контрольной группой (1987±24,48 г) (увеличение на 7,8 %; p ≤ 0,05). Среднесуточный прирост живой массы демонстрировал сходную динамику при введении бетаина. Помимо этого, введение бетаина способствовало снижению отложения абдоминального жира как у петушков, так и у курочек (p ≤ 0,05). Например, у курочек отложения абдоминального жира снижались в 1,4 раза в III и IV опытных группах по сравнению с контролем (p ≤ 0,05).
Таким образом, введение бетаина в рацион цыплят-бройлеров в условиях дефицита обменной энергии и некоторых аминокислот может способствовать улучшению показателей мясной продуктивности. Тенденция к снижению количества абдоминального жира при повышении дозировки бетаина свидетельствует о его вероятном влиянии на метаболизм липидов. Это согласуется с данными о роли бетаина в метилировании и 1102
осмопротекции (22). Ранее было показано, что добавление бетаина в рацион крыс с ожирением улучшило метаболизм метионина в печени, снизило содержание гомоцистеина и предотвращало накопление жира (23). Добавление бетаина в рацион серебряного карпа улучшало показатели роста и снижало отложение липидов (24). Аналогичные данные были получены на свиньях (25). Добавка бетаина уменьшала содержание жира в брюшной полости ягнят (26). Добавление бетаина в рацион гусей снижало потребление корма в период откорма, однако повышало усвояемость аминокислот и липолитическую активность (27).
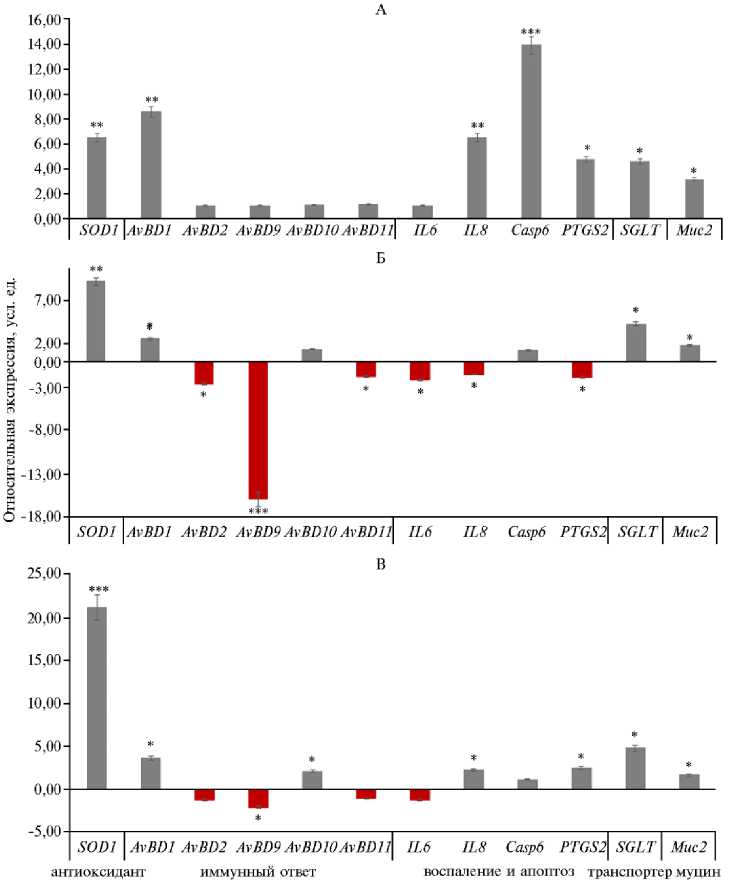
Рис. 2. Экспрессия ключевых генов у курочек ( Gallus gallus ) кросса Смена 9 в зависимости от включения в рацион добавки бетаина в разных дозах: А — II опытная группа, Б — III опытная группа, В — IV опытная группа. По оси абсцисс представлены изменения в экспрессии генов в опытных группах (II-IV) относительно контрольной группы I, где экспрессия генов условно принята за единицу (1). Снижение экспрессии генов в опытных группах по сравнению с контрольной характеризовалось отрицательными значениями, тогда как увеличение экспрессии — положительными значениями. Численные значения указывают на кратность отклонения экспрессии в опытных группах от экспрессии в контрольной группе. Описание групп см. в разделе «Методика».
*, ** и *** Различия при сравнении II-IV групп с контролем статистически значимы соответ- ственно при p ≤ 0,05, p ≤ 0,01 и p ≤ 0,001 согласно t-критерию Стьюдента.
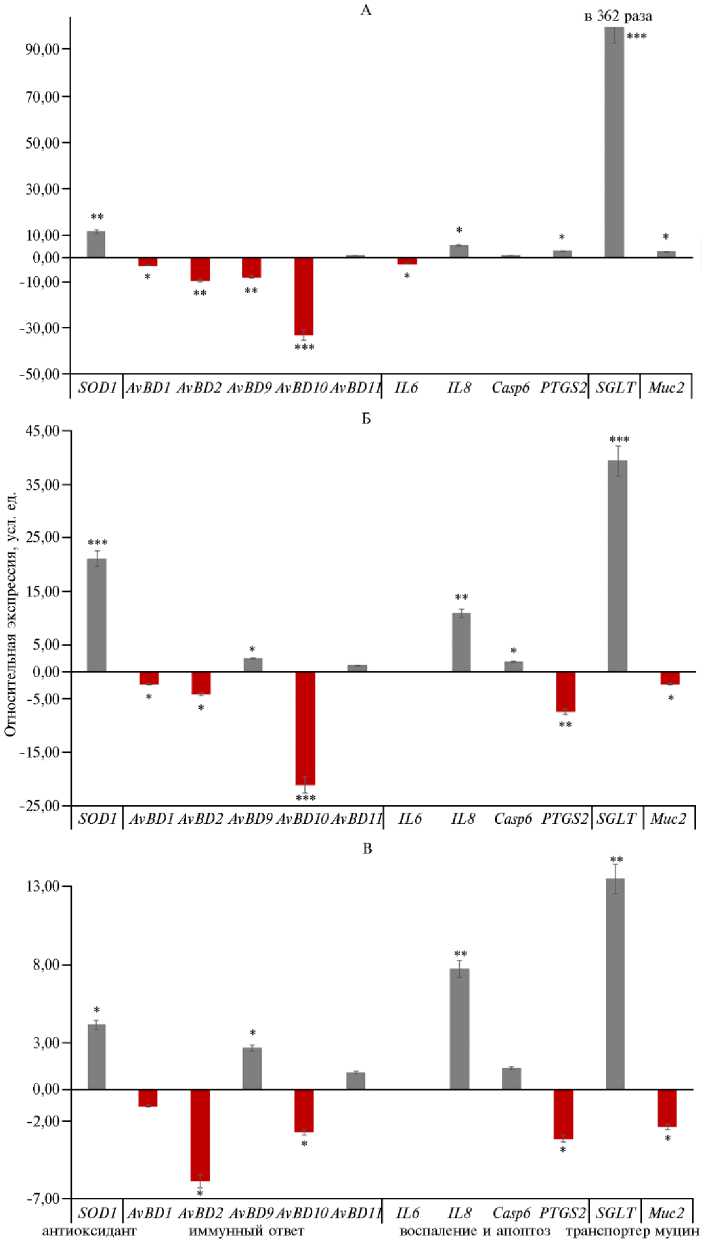
Рис. 3. Экспрессия ключевых генов у петушков (Gallus gallus) кросса Смена 9 в зависимости от включения в рацион добавки бетаина в разных дозах: А — II опытная группа, Б — III опытная группа, В — IV опытная группа. По оси абсцисс представлены изменения в экспрессии генов в опытных группах (II-IV) относительно контрольной группы I, где экспрессия генов условно принята за единицу (1). Снижение экспрессии генов в опытных группах по сравнению с контрольной характеризовалось отрицательными значениями, тогда как увеличение экспрессии — положительными значениями. Численные значения указывают на кратность отклонения экспрессии в опытных группах от экспрессии в контрольной группе. Описание групп см. в разделе «Методика».
-
*, ** и *** Различия при сравнении II-IV групп с контролем статистически значимы соответственно при p ≤ 0,05, p ≤ 0,01 и p ≤ 0,001 согласно t -критерию Стьюдента..
Одновременно с изучением зоотехнических показателей была проведена оценка экспрессии генов в образцах тканей кишечника. Данные количественной ПЦР с обратной транскрипцией (рис. 2 и 3) показали значительную вариацию экспрессии генов в зависимости от дозировки бетаина и пола цыплят-бройлеров.
У курочек экспрессия гена SOD1 , кодирующего супероксиддисму-тазу 1 — ключевой антиоксидантный фермент (28) увеличилась при введении 200 г/т бетаина (II группа) в 6,5 раза, 300 г/т бетаина (III группа) — в 9,2 раза (p ≤ 0,01), 400 г/т бетаина (IV группа) — в 21,1 раза (p ≤ 0,001) по сравнению с контролем (I группа). У петушков наблюдалось аналогичное увеличение в 11,3 и 21,1 раза для II и III групп (p ≤ 0,01), то есть происходило заметное улучшение антиоксидантной защиты под влиянием добавки, что было показано в других исследованиях (29). Ранее M. Shakeri с соавт. (30) обнаружили, что добавление бетаина может снизить окислительный стресс у цыплят-бройлеров при повышенных температурах окружающей среды за счет увеличения активности антиоксидантных ферментов, таких как глутатионпероксидаза (GSH Px). R. Chen с соавт. (31) установили, что добавление бетаина не только улучшило показатели роста бройлеров, но и предотвратило ухудшение качества мяса при стрессе от транспортировки за счет изменения анаэробного гликолиза в мышцах и антиоксидантной активности.
Интересно, что в экспрессии генов, связанных с синтезом антимикробных пептидов (группа AvBD ), наблюдались выраженные половые различия (p ≤ 0,05). Так, у курочек экспрессия AvBD1 в ответ на введение в комбикорм 200 г/т бетаина достигала 8,6-кратного увеличения (p ≤ 0,01) по сравнению с контролем, что может свидетельствовать о более эффективной антимикробной защите слизистой желудочно-кишечного тракта (32). Однако при добавлении 300 г/т бетаина наблюдалось снижение количества мРНК AvBD2 и AvBD9 (соответственно в 2,6 и 16 раз, p ≤ 0,05). Это указывает на иммунную регуляцию в ответ на избыточное количество бетаина (33). У петушков также были отмечены аналогичные изменения, особенно выраженные для AvBD2 . Относительное количество мРНК этого гена снизился на 9,9 раза во II опытной группе (p ≤ 0,01) по сравнению с I группой. Вероятно, большие дозы бетаина вызывают регуляторные реакции в антимикробной системе, что, в свою очередь, оказывает влияние на защитные функции организма птицы (6).
У курочек экспрессия провоспалительных генов IL6, IL8 и PTGS2 (циклооксигеназа-2) снижалась в III группе в 1,57-2,22 раза (p ≤ 0,05) по сравнению с контролем. Ослабление экспрессии провоспалительных цитокинов и PTGS2 может свидетельствовать о потенциальном противовоспалительном действии бетаина. Известно, что PTGS2 играет ключевую роль в синтезе простагландинов — медиаторов воспаления (34). Воспаление — это один из существенных факторов, способствующих развитию стресса у птицы (35). Поэтому снижение экспрессии PTGS2 приводит к повышению устойчивости к стрессу, улучшению общего физиологического состояния и росту продуктивности. Экспрессия гена Casp6, напротив, значительно увеличивалась у курочек при скармливании 200 г/т бетаина (в 13,9 раза по сравнению с контролем; p ≤ 0,01).
Ген Casp6 кодирует каспазу-6 — ключевой фермент в каскаде апоптоза (36). Индукция его экспрессии приводит к удалению поврежденных или дисфункциональных клеток, что, в конечном итоге, способствует поддержанию клеточного гомеостаза и предотвращению развития патологических состояний (37). У петушков из III группы экспрессия гена IL8 увеличилось в 10,9 раза и отмечалось снижение экспрессии гена PTGS2 в 7,5 раза (p ≤ 0,01). Эти изменения подчеркивают различную модулирующую роль бетаина в регуляции экспрессии генов, связанных с воспалением и апоптозом, у птиц разных полов. Они могут быть связаны с половыми различиями в метаболизме, гормональном статусе и иммунной регуляции (38).
Очень интересные результаты были получены при оценке экспрессии генов, связанных с транспортом глюкозы. У петушков экспрессия SGLT2 возросла в 362 раза во II группе (p ≤ 0,001) и достигала увеличения в 39,4 и 13,5 раза соответственно в III и IV группах по сравнению с I группой (p ≤ 0,01). SGLT2 — белок-транспортер, присутствующий в эпителиальных клетках слизистой оболочки кишечника птицы (39). Основная функция SGLT2 — осуществлять вторично активный транспорт глюкозы из просвета кишечника в клетки, используя электрохимический градиент ионов натрия (40). Поэтому активация экспрессии мРНК гена SGLT2 указывает на улучшение всасывания глюкозы. Уменьшение количества мРНК гена SGLT2 при повышении концентрации бетаина могло означать, что бетаин имеет оптимальные концентрации для проявления эффекта и дальнейшее увеличение дозы подавляет метаболические процессы, необходимые для экспрессии SGLT2 . Аналогичные, но значительно менее выраженные изменения наблюдались и у курочек: в опытных группах экспрессия SGLT2 возросла в 4,3-4,8 раза (p ≤ 0,05).
Экспрессия гена MUC2 у курочек умеренно увеличивалась (в 3,2 раза) во II группе по сравнению с контролем (p ≤ 0,05), что может благоприятно сказаться на состоянии слизистой оболочки кишечника. У петушков также наблюдалось повышение лишь при 200 г/т бетаина в корме (в 2,6 раза по сравнению с контролем; p ≤ 0,05).
MUC2 (муцин 2) кодирует основной гель-образующий белок муцин, который служит ключевым компонентом слизистого слоя, выстилающего поверхность кишечника (41). Слизистый слой выполняет множество важных функций, включая физическую защиту эпителиальных клеток от воздействия патогенных микроорганизмов, токсинов, механического повреждения и агрессивных факторов пищеварения (42). Он служит средой для адгезии комменсальных бактерий, которые влияют на иммунную функцию, пищеварение и общее здоровье особи (43). Индукция экспрессии MUC2 может способствовать снижению проницаемости кишечника для патогенов и токсинов, уменьшению воспаления, усилению всасывания питательных веществ и устойчивости к заболеваниям, улучшению общего здоровья пищеварительной системы и организма в целом (44).
Значительные различия в экспрессии генов между курочками и петушками могут быть обусловлены рядом факторов, например гормональным статусом (45). Тестостерон (доминирующий гормон у самцов) снижает чувствительность к воспалительным процессам (46). Самцы и самки различаются по строению внутренних органов, особенностям метаболизма и гормональному профилю (47). У самцов более развит мышечный аппарат и костяк, требующие большего количества энергии и питательных веществ (48). При этом самцы более подвижны и тратят больше энергии на движения и территориальное поведение, что также увеличивает их потребность в питательных веществах (49). Как следствие. реакция организма петушков на добавление бетаина в корм может отличаться от таковой у курочек. Гены, связанные прежде всего с иммунитетом и распределением питательных веществ, тоже могут проявлять различную экспрессию в зависимости от пола птицы.
Таким образом, введение бетаина в рацион цыплят-бройлеров кросса Смена 9, получающих корм с пониженным содержанием обменной энергии (на 5 %), лизина (на 10 %) и метионина (на 10 %), вызывает каскад взаимосвязанных эффектов, приводящих к улучшениям в зоотехнических показателях и модуляции экспрессии ряда ключевых генов в тканях слепых отростков кишечника. Наиболее значительное увеличение живой массы на 7,2 и 7,3 % (p ≤ 0,05) наблюдалось при повышенных дозировках бетаина (300 и 400 г/т). При этом было выявлено существенное снижение отложения абдоминального жира (p ≤ 0,05), что указывает на положительное влияние бетаина на липидный метаболизм. На молекулярно-клеточном уровне бетаин вызывал дифференцированную реакцию, характеризующуюся повышением экспрессии SOD1 — ключевого гена антиоксидантной защиты. Особо стоит отметить впервые выявленные половые различия в модуляции экспрессии генов иммунной системы под воздействием бетаина. У курочек наблюдалось более чем 8-кратное увеличение экспрессии гена AvBD1 при дозе бетаина 200 г/т (p ≤ 0,01), что указывает на укрепление защитных свойств слизистой оболочки кишечника. Однако при более высоких дозах отмечалось снижение экспрессии других генов семейства AvBD (p ≤ 0,05). Одновременное уменьшение количества мРНК провоспалительных генов IL6, IL8 и PTGS2 предполагает, что бетаин обладает противовоспалительными свойствами и может способствовать снижению воспалительных реакций в тканях, в особенности у курочек. Кроме того, усиление барьерной функции кишечника, обусловленное повышенной экспрессией MUC2, вероятно, позволяет улучшать усвояемость аминокислот и других ценных веществ, что способствует поддержанию продуктивности в условиях недостатка питательных веществ и энергии. Особое внимание заслуживает изменение экспрессии гена SGLT2 (натрий-глюкозный транспортер второго типа), который ответственен за поглощение глюкозы в кишечнике. У петушков в группе с дозировкой бетаина 200 г/т произошел резкий скачок экспрессии SGLT2 — в 362 раза (p ≤ 0,001), а при увеличении дозировки до 300 и 400 г/т — соответственно в 39,4 и 13,5 раза (p ≤ 0,01), что отражает возможное повышение эффективности транспорта глюкозы и энергетического обеспечения организма. В целом, полученные данные демонстрируют преимущества добавления бетаина в рацион цыплят-бройлеров. Представляют интерес выявленные половые и дозозависимые различия в ответе на бетаин. Настоящая работа впервые демонстрирует сложный, дифференцированный эффект бетаина на экспрессию генов, связанных с иммунитетом, воспалением и метаболизмом глюкозы, в зависимости от пола и дозировки. Половые и дозозависимые различия в ответе на бетаин подчеркивают необходимость индивидуального подхода к применению и дозировкам этой кормовой добавки в рационах сельскохозяйственной птицы. На следующем этапе исследования мы планируем провести секвенированием полного тран-скриптома (РНК-секвенирование) с целью выявления глобального профиля экспрессии генов в различных органах и тканях птицы в ответ на введение бетаина. Это позволит определить дополнительные сигнальные пути и механизмы, вовлеченные в адаптивную реакцию организма цыплят-бройлеров на дефицит питательных веществ и энергии, выявить новые РНК-маркеры действия бетаина, а также подтвердить существующие гипотезы относительно половой специфичности уровней экспрессии ключевых генов.
1 000 «БИОТРОФ+»,
