Изменение кинетических параметров слабо-связанной с клеточной стенкой пероксидазы суспензионных клеток картофеля в начальный период инфицирования

Автор: Граскова И.А., Антипина И.В., Потапенко О.Ю., Войников В.к
Журнал: Журнал стресс-физиологии и биохимии @jspb
Рубрика: Original article
Статья в выпуске: 1 т.2, 2006 года.
Бесплатный доступ
Изучали изменение кинетических параметров слабо-связанных с клеточной стенкой пероксидаз суспензионных клеток картофеля устойчивого (Луговской) и восприимчивого (Лукьяновский) сортов в начальный период заражения вирулентным штаммом 5369 Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh. Было показано, что слабо-связанные с клеточной стенкой пероксидазы устойчивого и восприимчивого сортов картофеля в отсутствии патогена конкурентно ингибированы. В первые моменты инфицирования резкое повышение активности фермента устойчивого сорта происходит за счет увеличения сродства фермента к субстрату в результате снижения конкурентного ингибирования. У восприимчивого сорта картофеля увеличение активности фермента происходит по другому механизму.
Патогенез, слабо-связанная с клеточной стенкой пероксидаза
Короткий адрес: https://sciup.org/14323451
IDR: 14323451
The change of kinetik parameters of the weak-associated with wall cell peroxidase in the suspension culture of potato cells in the beginning of infection
The change in kinetic parameters of extracellular peroxidase of suspension cells of resistant potato variety (Lugovskoi) and sensitive variety (Lukynovskii) in the initial period of infection by 5369 Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh. pathogen was examined. Extracellular peroxidases of resistant and sensitive potato variety without pathogens were shown to be concurrently inhibited. At the beginning of infection enzyme activity was extremely increased due to enhanced affinity to substrate as a result of reducing of competitive inhibiting. In increasing enzyme activity is sensitive potato variant evidently caused by other mechanism.
Текст обзорной статьи Изменение кинетических параметров слабо-связанной с клеточной стенкой пероксидазы суспензионных клеток картофеля в начальный период инфицирования
Изучали изменение кинетических параметров слабо-связанных с клеточной стенкой пероксидаз суспензионных клеток картофеля устойчивого (Луговской) и восприимчивого (Лукьяновский) сортов в начальный период заражения вирулентным штаммом 5369 Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh. Было показано, что слабосвязанные с клеточной стенкой пероксидазы устойчивого и восприимчивого сортов картофеля в отсутствии патогена конкурентно ингибированы. В первые моменты инфицирования резкое повышение активности фермента устойчивого сорта происходит за счет увеличения сродства фермента к субстрату в результате снижения конкурентного ингибирования. У восприимчивого сорта картофеля увеличение активности фермента происходит по другому механизму.
Изучение особенностей метаболизма растений при патогенезе является важным условием для понимания механизмов как поражения растения патогеном, так и его устойчивости при инфицировании. Известно, что при патогенезе возникают различные нарушения в обмене веществ растительной клетки (Дмитриев, 2003). При этом значительные изменения претерпевают ее мембранные и белковые компоненты, в том числе ферменты (Ладыженская, Проценко, 2002). Особая роль в защитной реакции растительной клетки на патогенез принадлежит пероксидазам (Минибаева, Гордон, 2003; Huckelboven et al, 2003). Увеличение активации этих ферментов в ответ на заражение отмечено у многих видов растений и при различных по природе инфекциях (Хайрулин и др., 2000; Bolvel et al, 2002). В последнее время особое внимание уделяется изучению внеклеточных пероксидаз (пероксидаз клеточной стенки) (Bolvel et al, 2002). Это внимание объясняется тем, что именно внеклеточные пероксидазы первыми сталкиваются с патогенами и именно они первыми вступают в борьбу с ними (Минибаева, Гордон, 2003).
Ранее при изучении динамики активности слабо-связанных с клеточной стенкой пероксидаз картофеля при патогенезе Clavibacter michiganensis subsp. sepedonicus было установлено, что активность этих пероксидаз устойчивого сорта картофеля выше, чем восприимчивого как в контроле, так и после инфицирования (Граскова и др., 2004). Причем наибольший пик активности этих ферментов у восприимчивого сорта картофеля наблюдается через 1.5 – 2 часа после инфицирования и он зависит от экспрессии генома. У устойчивого сорта картофеля резкое повышение активности слабо-связанных с клеточной стенкой пероксидаз происходит в первые минуты инфицирования и это повышение активности не зависит от синтеза белка de novo. Второй пик активности ферментов у этого сорта наблюдается через 1.5 – 2 часа после инфицирования и он связан с изменением экспрессии генома.
Можно предположить, что резкое и не связанное с синтезом белков повышение активности слабо-связанных с клеточной стенкой пероксидаз устойчивого сорта картофеля в первые моменты инфицирования обусловлено, вероятно, изменением кинетических параметров ферментов. Поэтому целью настоящей работы было провести сравнительное изучение кинетических параметров слабо-связанных с клеточной стенкой пероксидаз контрастных по устойчивости к кольцевой гнили сортов картофеля и проверить возможность регуляции активности этого фермента в первые моменты инфицирования за счет изменения его кинетических параметров.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовали суспензионные культуры клеток сортов картофеля ( Solanum tuberosum L.) Луговской (устойчивый к патогену) и Лукьяновский (восприимчивый к патогену). Пробирочные растения выращивали из черенков на агаризированной МС-среде с добавлением гормонов и витаминов (Бутенко и др., 1984). Для получения суспензионных культур клеток картофеля 2-5 мг каллусной листовой ткани помещали в 100 мл жидкой питательной МС-среды с добавлением гормонов и витаминов (Бутенко и др., 1984) и выращивали при непрерывном встряхивании до получения суспензионных культур.
Для заражения использовали штамм 5369 (вирулентный, агрессивный) Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh, полученный из НИИ картофельного хозяйства (пос. Коренево, Московская обл.). К суспензионным культурам тканей клеток картофеля добавляли 1 мл свежеприготовленной бактериальной суспензии (2х108 кл/мл) и культивировали в течение 3 часов при температуре 250С. Отборы проб проводили через каждые 5 мин в течение первого часа и через 30 мин в последующие 2 часа.
Пероксидазную активность анализировали в суспензионных культурах клеток картофеля. Для экстракции внеклеточных пероксидаз навески по 1 г суспензионных тканей картофеля помещали в шприц, заливали 10 мл холодного цитратно-фосфатного буфера (0.1 М, рН 6.2- устойчивый сорт, рН 5.4 – восприимчивый сорт) и дважды выдерживали при вакуумной фильтрации по 1 мин. В фильтрате определяли пероксидазную активность (Граскова и др., 2004). Для этого измеряли увеличение оптической плотности при 580 нм в реакционной смеси из 0.5 мл 0.1 М цитратно-фосфатного буфера (рН 6.2 и 5.4), 0.5 мл 0.3% Н 2 О 2 и 0.5 мл (0.035%, 0.04%, 0.045%, 0.05%,0.055%) гваякола (“Sigma”); конечные концентрации. Активность пероксидазы определяли при 25оС через 5 мин в первый час и через 30 мин в последующие 2 часа. Активность фермента рассчитывали по методу Бояркина с модификациями (Bolvel et al., 1999). Количество белка определяли по методу Lowry (Lowry et al., 1951). Константы рассчитывали по методу Лайнуивера-Берка (Виноградова, 1978). На рисунках указаны средние значения и стандартные ошибки.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Заражение клеток суспензионных культур картофеля вирулентным штаммом 5369 Clavibacter michiganensis subsp. sepedonicus вызывало увеличение активности слабосвязанных с клеточной стенкой пероксидаз как у устойчивого, так восприимчивого сортов картофеля (рис.1). Обращает на себя внимание резкое увеличение активности этого фермента у устойчивого сорта в первые моменты инфицирования. Поэтому было проведено изучение кинетических параметров слабосвязанных с клеточной стенкой пероксидаз, выделенных из суспензии клеток картофеля именно в это время.
Зависимость активности фермента от концентрации субстрата выражается гиперболической функцией и подчиняется уравнению Михаэлиса-Ментен. Графическое изображение этой зависимости в двойных обратных координатах дало серию прямых линий, различающихся между собой по углу наклона в зависимости как от инфицирования, так и от устойчивости сорта картофеля к патогену (рис.2). В контрольном варианте опыта (без патогена) наибольшее значение кажущейся константы Михаэлиса (Км) имели слабо-связанных с клеточной стенкой пероксидазы восприимчивого к патогену сорта картофеля. Км устойчивого сорта имела меньшее значение. Причем величина Vmax у ферментов обоих сортов была равной (табл.1). Это свидетельствует о том, что в отсутствие патогена слабо-связанных с клеточной стенкой пероксидазы восприимчивого сорта конкурентно ингибированы, по сравнению с устойчивым сортом картофеля.
На этом основании можно предположить, что по этой причине в контрольном варианте (без патогена) активность ферментов выше у устойчивого сорта по сравнению с восприимчивым cортом. Возможно, что меньшая активность слабо-связанных с клеточной стенкой пероксидаз восприимчивого сорта из-за их конкурентного ингибирования, является одной из причин чувствительности этого сорта к патогену.
Добавление патогена к клеткам устойчивого сорта привело к резкому снижению К м (рис.2). При этом величина V max достоверно не изменилась (табл.1). Это свидетельствует о том, что до инфицирования слабо-связанных с клеточной стенкой пероксидазы у устойчивого сорта были конкурентно ингибированы, а в ответ на инфицирование произошло снижение этого ингибирования и как следствие - увеличение их активности. Эти данные позволяют заключить, что механизм резкой активации слабо-связанных с клеточной стенкой пероксидаз устойчивого сорта в первые моменты действия патогена, связан с изменением кинетических параметров этих ферментов, в частности, с резким увеличением их сродства к субстрату в результате снижения конкурентного ингибирования. Каков молекулярный механизм этого события предстоит еще выяснить.
Таблица 1 Изменение кинетических параметров слабо-связанных с клеточной стенкой пероксидазы суспензионных клеток, контрастных по устойчивости к патогену сортов картофеля.
|
Сорт |
Контроль |
+ Cms, 10 мин |
|
Луговской (устойчивый сорт) |
K m 8,9 + 3.3 |
K m 3,1 + 0,9 |
|
V max 0.9 + 0.3 |
V max 1,12 + 0,1 |
|
|
Лукьяновский (восприимчивый сорт) |
K m 21,3 + 4,6 |
K m 2,1 + 0,46 |
|
V max 0,9 + 0,12 |
V max 0,3 + 0,03 |
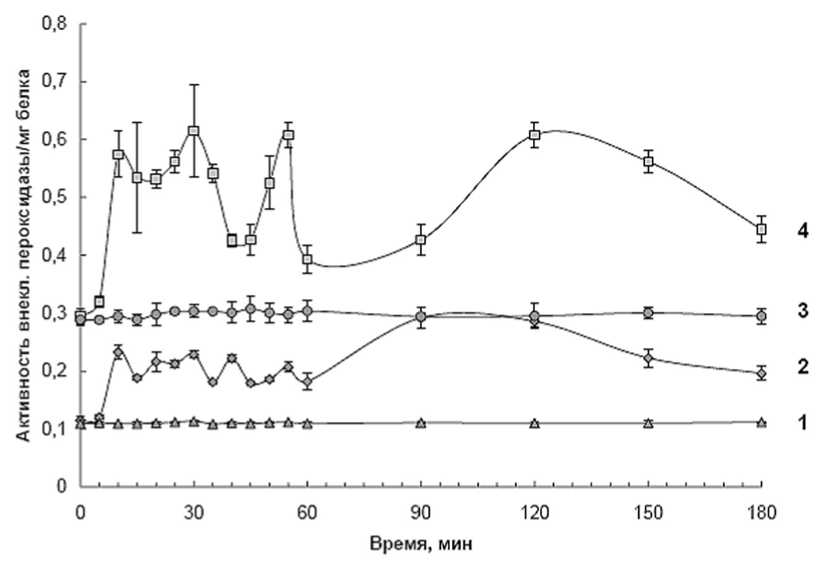
Рис. 1. Динамика активности слабо-связанной с клеточной стенкой пероксидазы суспензионных клеток картофеля двух контрастных по устойчивости к патогену сортов: Лукьяновский (восприимчивый) и Луговской (устойчивый).
1- Контроль (сорт Лукьяновский); 2 – культивирование с патогеном (сорт Лукьяновский); 3 – Контроль (сорт Луговской); 4 – культивирование с патогеном (сорт Луговской).
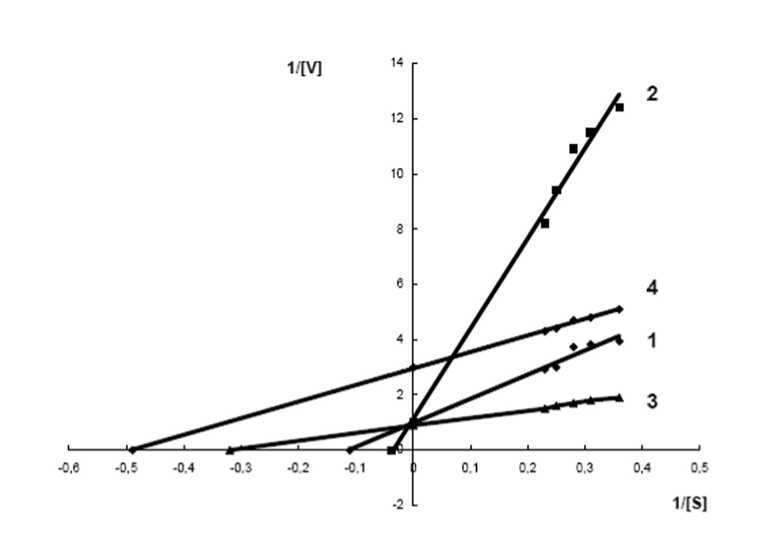
Рис. 2. Изменение кинетических параметров слабо-связанной с клеточной стенкой пероксидазы устойчивого и восприимчивого сортов картофеля в начальный период инфицирования.
1 - Сорт Луговской, контроль; 2 – Сорт Лукьяновский, контроль; 3 – Сорт Луговской, заражение 10 мин; 4 – Сорт Лукьяновский, заражение 10 мин.
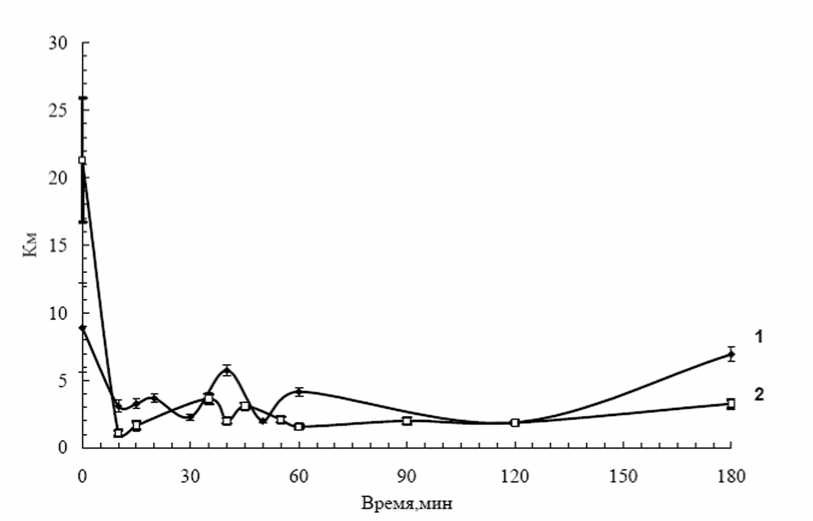
Рис. 3. Динамика К м слабо-связанной с клеточной стенкой пероксидазы устойчивого (Луговской) и восприимчивого (Лукьяновский) сортов картофеля при трех часовом заражении
патогеном.
1 – сорт Луговской; 2 – сорт Лукьяновский.
У восприимчивого сорта картофеля патоген так же вызывал снижение величины Км, но при этом происходило резкое снижение значения Vmax (рис.2). Следовательно, можно говорить о том, что в случае восприимчивого сорта в первые моменты патогенеза происходит, с одной стороны, увеличение сродства фермента и субстрата (снижение Км), а с другой - снижение скорости образования и распада фермент-субстратного комплекса (снижение Vmax).
Одновременное протекание этих двух процессов, разнонаправленных в отношении активации фермента, возможно, являются причиной меньшего (по сравнению с устойчивым сортом) увеличения активности слабо-связанных с клеточной стенкой пероксидаз при патогенезе.
В течение эксперимента (0-180 мин инкубации с патогеном) значения К м внеклеточных пероксидаз обоих сортов картофеля изменялись (рис.3). При этом максимальные значения К м соответствовали минимальным активностям ферментов и, наоборот, максимумы Км - минимумам активности (рис.1,3). Так, у устойчивого сорта максимальные значения активности ферментов, отмеченные через 10, 30, 50-55 и 120 минут инкубации клеток с патогеном, совпадали с минимальными К м . Максимумы значений К м через 20, 40 и 60 минут инкубации клеток картофеля с патогенными бактериями соответствовало "провалам" в активности ферментов (рис.1,3). Сходная закономерность отмечена и для восприимчивого сорта. Эти результаты позволяют предполагать, что при действии патогена изменения активности слабосвязанных с клеточной стенкой пероксидаз клеток картофеля, отмеченные при изучении динамики их активности, обусловлены изменениями кинетических параметров этих ферментов, в частности, они обусловлены изменениями в сродстве ферментов к субстратам.
Следовательно, на основании изложенных в этой статье и полученных ранее (Граскова и др., 2004) результатов можно предполагать, что при патогенезе регуляция активности слабосвязанных с клеточной стенкой пероксидаз суспензионных клеток картофеля осуществляется за счет изменения кинетических параметров этих ферментов. Причем в первые моменты инфицирования клеток изменяются кинетические параметры уже существующих молекул ферментов. С увеличение длительности действия патогена, вероятно, синтезируются новые молекулы ферментов (Граскова и др., 2003) (возможно, другие их изоформы) с другими кинетическими параметрами.
Таким образом, слабо-связанных с клеточной стенкой пероксидазы устойчивого и восприимчивого сортов картофеля в отсутствии патогена конкурентно ингибированы. В первые моменты инфицирования резкое повышение активности ферментов устойчивого сорта происходит за счет увеличения их сродства к субстрату в результате снижения конкурентного ингибирования. У восприимчивого сорта картофеля увеличение активности ферментов происходит по другому механизму.
Список литературы Изменение кинетических параметров слабо-связанной с клеточной стенкой пероксидазы суспензионных клеток картофеля в начальный период инфицирования
- Бояркин А.Н. (1951) Быстрый метод определения активности пероксидазы. Биохимия, 16, Вып.4, 352-355.
- Бутенко Р.Г., Хромова Л.М., Седнина Т.В. (1984) Методические указания по получению вариантных линий и растений у разных сортов картофеля. ВАСХНИЛ, М, 28.
- Виноградова Г.П. (1978) Молекулярные основы действия ферментов. Вища школа, Киев, 279.
- Граскова И.А., Боровский Г.Б., Колесниченко А.В., Войников В.К. (2004) Пероксидаза как компонент сигнальной системы клеток картофеля при патогенезе кольцевой гнили. Физиология растений. 51,№5, 692-697.
- Граскова И.А., Боровский Г.Б., КолесниченкоА.В., Войников В.К. (2003) Различия в механизмах изменения активности пероксидазыустойчивогои восприимчивого сортов картофеля при патогенезе кольцевой гнили. Вестник ХНАУ. Серия Биология. 3(2), 70-71.
- Дмитриев А.П. (2003) Сигнальные молекулы растений для активации защитных реакций в ответ на биотический стресс. Физиология растений. 50, №3, 465-474.
- Ладыженская Э.П., Проценко М.А. (2002) Биохимические механизмы передачи внешних сигналов через плазмалемму растительной клетки при регуляции покоя и устойчивости. Биохимия. 67, вып.2, 181-193.
- Минибаева Ф.М., Гордон Л.Х. (2003) Продукция супероксида и активность внеклеточной пероксидазы в растительных тканях при стрессе. Физиология растений. 50, №3, 459-464.
- Хайрулин P.M., Юсупова З.Р., Трошина Н.Б. (2000) Защитные реакции пшеницы при инфицировании грибными патогенами. 2. Активация анионных изоформ пероксидазы в проростках пшеницы при инфицировании спорами Tilletia caries. Физиология растений. 47, №1, 114-119.
- Bolwell G.P., Bindschedler L.V., Blee K.A., Butt V.S., Davies D.R., Gardner S.L., Gerrich C., Minibayeva F. (2002) The Apoplastic oxidative burst in response to biotic stress in plants: a tree-component system. J. Exp. Bot. 53, 1367-1376.
- Bolwell G.P., Blee K.A., Butt V.S., Davies D.R., Gardner S.L., Gerrich C, Minibayeva F., Rowntree E.G., Wojtaszek P. (1999) Recent Advances in Understanding the Origin of the Apoplastik Oxidative Burst in Plant Cells. Free Radical Res. 31, 137-145.
- Huckelboven R., Kogel K.-H. (2003) Reactive oxygen intermediates in plant-microbe interactions: who is who in powdery mildew resistance? Planta. 216, N6, 891-902.
- Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. (1951) Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 193, 265-275.
